

Advancements in Understanding the Hide-and-Seek Strategy of Hibernating Breast Cancer Cells and Their Implications in Oncology from a Broader Perspective: A Comprehensive Overview


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Dormancy in Breast Cancer
3. Mechanisms of Hibernating Breast Cancer Cells
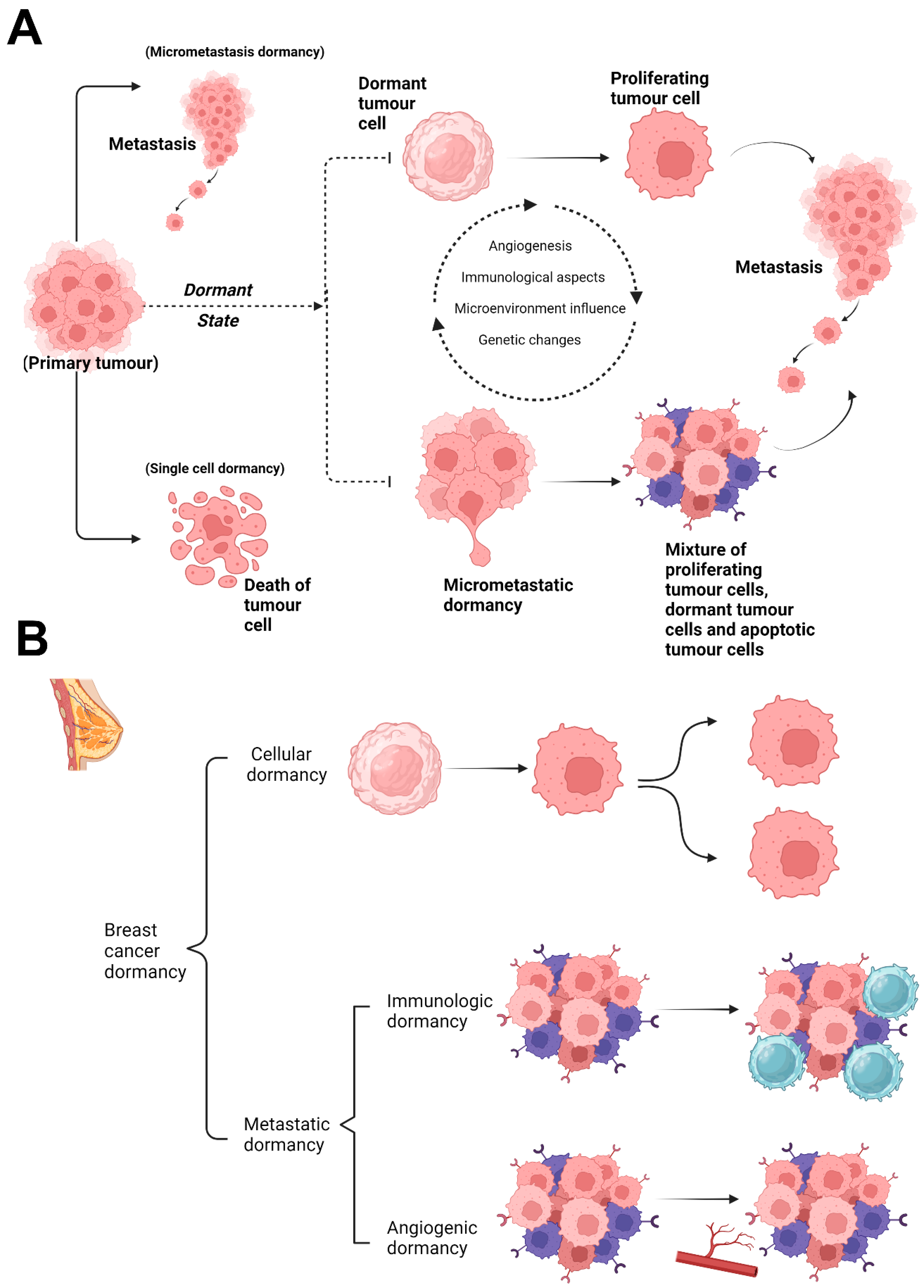
3.1. Single-Cell Dormancy
3.2. Micrometastatic Dormancy
3.2.1. Angiogenic Dormancy
3.2.2. Immunologic Dormancy
4. Identification and Characterisation of Hibernating Breast Cancer Cells
5. Potential Therapeutic Strategies Targeting Hibernating Breast Cancer Cells
5.1. Targeting the Microenvironment
5.2. Targeting Angiogenesis
5.3. Targeting Signal Transduction
5.4. Activating the Immune System
6. Evolution and Cutting-Edge Developments in Hormone-Targeted Therapy and Immunotherapeutics for Breast Cancer
6.1. Historical Perspective and Breakthroughs in Hormone-Targeted Therapy
6.2. Recent Advances in Immunotherapeutics
6.3. Most Recent Innovations
7. Epigenetic Regulation of Tumour Dormancy
7.1. DNA Methylation
7.2. Histone Modifications
7.3. Chromatin Remodelling
8. The Epigenetic Influence of G9a Enzyme in Breast Cancer Relapse and Most Recent Therapeutic Insights
9. Emerging Biomarkers in Breast Cancer
9.1. Circulating Tumour Cells (CTCs)
9.2. Liquid Biopsies
9.3. Gene Expression Signatures
9.4. Tumour Microenvironment
10. Innovative Bispecific Antibodies: A Promising Approach in Breast Cancer Therapy
11. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, J.; Chan, P.S.; Lok, V.; Chen, X.; Ding, H.; Jin, Y.; Yuan, J.; Lao, X.; Zheng, Z.-J.; Wong, M.C. Global Incidence and Mortality of Breast Cancer: A Trend Analysis. Aging 2021, 13, 5748–5803. [Google Scholar] [CrossRef]
- Berrino, F.; Villarini, A.; Gargano, G.; Krogh, V.; Grioni, S.; Bellegotti, M.; Venturelli, E.; Raimondi, M.; Traina, A.; Zarcone, M.; et al. The Effect of Diet on Breast Cancer Recurrence: The DIANA-5 Randomized Trial. Clin. Cancer Res. 2024, 30, 965–974. [Google Scholar] [CrossRef]
- Roy, M.; Biswas, J.; Datta, A. Breast Cancer: Epidemiology, Types, Diagnosis, and Treatment. In Genetics and Epigenetics of Breast Cancer; Roy, M., Biswas, J., Datta, A., Eds.; Springer Nature: Singapore, 2023; pp. 1–24. ISBN 978-981-19992-5-3. [Google Scholar]
- Roheel, A.; Khan, A.; Anwar, F.; Akbar, Z.; Akhtar, M.F.; Imran Khan, M.; Sohail, M.F.; Ahmad, R. Global Epidemiology of Breast Cancer Based on Risk Factors: A Systematic Review. Front. Oncol. 2023, 13, 1240098. [Google Scholar] [CrossRef]
- Duggan, C.; Trapani, D.; Ilbawi, A.M.; Fidarova, E.; Laversanne, M.; Curigliano, G.; Bray, F.; Anderson, B.O. National Health System Characteristics, Breast Cancer Stage at Diagnosis, and Breast Cancer Mortality: A Population-Based Analysis. Lancet Oncol. 2021, 22, 1632–1642. [Google Scholar] [CrossRef]
- Sinha, G.; Ferrer, A.I.; Moore, C.A.; Naaldijk, Y.; Rameshwar, P. Gap Junctions and Breast Cancer Dormancy. Trends Cancer 2020, 6, 348–357. [Google Scholar] [CrossRef]
- Blasco, M.T.; Espuny, I.; Gomis, R.R. Ecology and Evolution of Dormant Metastasis. Trends Cancer 2022, 8, 570–582. [Google Scholar] [CrossRef]
- Rossari, F.; Zucchinetti, C.; Buda, G.; Orciuolo, E. Tumor Dormancy as an Alternative Step in the Development of Chemoresistance and Metastasis-Clinical Implications. Cell. Oncol. 2020, 43, 155–176. [Google Scholar] [CrossRef]
- Dalla, E.; Sreekumar, A.; Aguirre-Ghiso, J.A.; Chodosh, L.A. Dormancy in Breast Cancer. Cold Spring Harb. Perspect. Med. 2023, 13, a041331. [Google Scholar] [CrossRef]
- Demicheli, R.; Abbattista, A.; Miceli, R.; Valagussa, P.; Bonadonna, G. Time Distribution of the Recurrence Risk for Breast Cancer Patients Undergoing Mastectomy: Further Support about the Concept of Tumor Dormancy. Breast Cancer Res. Treat. 1996, 41, 177–185. [Google Scholar] [CrossRef]
- Parker, A.L.; Cox, T.R. The Role of the ECM in Lung Cancer Dormancy and Outgrowth. Front. Oncol. 2020, 10, 567139. [Google Scholar] [CrossRef]
- Balayan, V.; Guddati, A.K. Tumor Dormancy: Biologic and Therapeutic Implications. World J. Oncol. 2022, 13, 8–19. [Google Scholar] [CrossRef]
- Rabinovsky, R.; Uhr, J.W.; Vitetta, E.S.; Yefenof, E. Cancer Dormancy: Lessons from a B Cell Lymphoma and Adenocarcinoma of the Prostate. In Advances in Cancer Research; Academic Press: Cambridge, MA, USA, 2007; Volume 97, pp. 189–202. [Google Scholar]
- Werner, S.; Heidrich, I.; Pantel, K. Clinical Management and Biology of Tumor Dormancy in Breast Cancer. Semin. Cancer Biol. 2022, 78, 49–62. [Google Scholar] [CrossRef]
- Ring, A.; Spataro, M.; Wicki, A.; Aceto, N. Clinical and Biological Aspects of Disseminated Tumor Cells and Dormancy in Breast Cancer. Front. Cell Dev. Biol. 2022, 10, 929893. [Google Scholar] [CrossRef]
- Aguirre-Ghiso, J.A. Models, Mechanisms and Clinical Evidence for Cancer Dormancy. Nat. Rev. Cancer 2007, 7, 834–846. [Google Scholar] [CrossRef]
- Ramamoorthi, G.; Kodumudi, K.; Gallen, C.; Zachariah, N.N.; Basu, A.; Albert, G.; Beyer, A.; Snyder, C.; Wiener, D.; Costa, R.L.B.; et al. Disseminated Cancer Cells in Breast Cancer: Mechanism of Dissemination and Dormancy and Emerging Insights on Therapeutic Opportunities. Semin. Cancer Biol. 2022, 78, 78–89. [Google Scholar] [CrossRef]
- Sosa, M.S.; Bragado, P.; Aguirre-Ghiso, J.A. Mechanisms of Disseminated Cancer Cell Dormancy: An Awakening Field. Nat. Rev. Cancer 2014, 14, 611–622. [Google Scholar] [CrossRef]
- Townson, J.L.; Chambers, A.F. Dormancy of Solitary Metastatic Cells. Cell Cycle 2006, 5, 1744–1750. [Google Scholar] [CrossRef]
- Weston, W.A.; Barr, A.R. A Cell Cycle Centric View of Tumour Dormancy. Br. J. Cancer 2023, 129, 1535–1545. [Google Scholar] [CrossRef]
- Richbourg, N.R.; Irakoze, N.; Kim, H.; Peyton, S.R. Outlook and Opportunities for Engineered Environments of Breast Cancer Dormancy. Sci. Adv. 2024, 10, eadl0165. [Google Scholar] [CrossRef]
- Ranganathan, A.C.; Adam, A.P.; Aguirre-Ghiso, J.A. Opposing Roles of Mitogenic and Stress Signaling Pathways in the Induction of Cancer Dormancy. Cell Cycle 2006, 5, 1799–1807. [Google Scholar] [CrossRef]
- Naumov, G.N.; MacDonald, I.C.; Weinmeister, P.M.; Kerkvliet, N.; Nadkarni, K.V.; Wilson, S.M.; Morris, V.L.; Groom, A.C.; Chambers, A.F. Persistence of Solitary Mammary Carcinoma Cells in a Secondary Site: A Possible Contributor to Dormancy1. Cancer Res. 2002, 62, 2162–2168. [Google Scholar]
- Bhojani, M.S.; Ross, B.D.; Rehemtulla, A. TRAIL and Anti-Tumor Responses. Cancer Biol. Ther. 2003, 2, 70–77. [Google Scholar] [CrossRef]
- Phipps, L.E.; Hino, S.; Muschel, R.J. Targeting Cell Spreading: A Method of Sensitizing Metastatic Tumor Cells to TRAIL-Induced Apoptosis. Mol. Cancer Res. 2011, 9, 249–258. [Google Scholar] [CrossRef]
- Fisher, J.L.; Thomas-Mudge, R.J.; Elliott, J.; Hards, D.K.; Sims, N.A.; Slavin, J.; Martin, T.J.; Gillespie, M.T. Osteoprotegerin Overexpression by Breast Cancer Cells Enhances Orthotopic and Osseous Tumor Growth and Contrasts with That Delivered Therapeutically. Cancer Res. 2006, 66, 3620–3628. [Google Scholar] [CrossRef]
- Neville-Webbe, H.L.; Cross, N.A.; Eaton, C.L.; Nyambo, R.; Evans, C.A.; Coleman, R.E.; Holen, I. Osteoprotegerin (OPG) Produced by Bone Marrow Stromal Cells Protects Breast Cancer Cells from TRAIL-Induced Apoptosis. Breast Cancer Res. Treat. 2004, 86, 271–282. [Google Scholar] [CrossRef]
- Maroun, C.R.; Rowlands, T. The Met Receptor Tyrosine Kinase: A Key Player in Oncogenesis and Drug Resistance. Pharmacol. Ther. 2014, 142, 316–338. [Google Scholar] [CrossRef]
- Zhang, X.H.-F.; Wang, Q.; Gerald, W.; Hudis, C.A.; Norton, L.; Smid, M.; Foekens, J.A.; Massagué, J. Latent Bone Metastasis in Breast Cancer Tied to Src-Dependent Survival Signals. Cancer Cell 2009, 16, 67–78. [Google Scholar] [CrossRef]
- Heim, S.; Teixeira, M.R.; Dietrich, C.U.; Pandis, N. Cytogenetic Polyclonality in Tumors of the Breast. Cancer Genet. Cytogenet. 1997, 95, 16–19. [Google Scholar] [CrossRef]
- Klein, C.A.; Blankenstein, T.J.; Schmidt-Kittler, O.; Petronio, M.; Polzer, B.; Stoecklein, N.H.; Riethmüller, G. Genetic Heterogeneity of Single Disseminated Tumour Cells in Minimal Residual Cancer. Lancet 2002, 360, 683–689. [Google Scholar] [CrossRef]
- Zhang, Y.; Che, G. Development of the Relationship between Angiogenesis and Tumor Dormancy. Chin. J. Clin. Oncol. 2007, 4, 277–281. [Google Scholar] [CrossRef]
- Indraccolo, S.; Stievano, L.; Minuzzo, S.; Tosello, V.; Esposito, G.; Piovan, E.; Zamarchi, R.; Chieco-Bianchi, L.; Amadori, A. Interruption of Tumor Dormancy by a Transient Angiogenic Burst within the Tumor Microenvironment. Proc. Natl. Acad. Sci. USA 2006, 103, 4216–4221. [Google Scholar] [CrossRef]
- Demicheli, R.; Retsky, M.W.; Hrushesky, W.J.; Baum, M. Tumor Dormancy and Surgery-Driven Interruption of Dormancy in Breast Cancer: Learning from Failures. Nat. Rev. Clin. Oncol. 2007, 4, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Hussein, O.; Komarova, S.V. Breast Cancer at Bone Metastatic Sites: Recent Discoveries and Treatment Targets. J. Cell Commun. Signal. 2011, 5, 85–99. [Google Scholar] [CrossRef]
- Young, S.A.E.; Heller, A.-D.; Garske, D.S.; Rummler, M.; Qian, V.; Ellinghaus, A.; Duda, G.N.; Willie, B.M.; Grüneboom, A.; Cipitria, A. From Breast Cancer Cell Homing to the Onset of Early Bone Metastasis: The Role of Bone (Re)Modeling in Early Lesion Formation. Sci. Adv. 2024, 10, eadj0975. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Z.; Wu, K.-L.; Yu, L.; Wang, C.; Ding, H.; Gao, Y.; Sun, H.; Wu, Y.-H.; Xia, M.; et al. Siglec-15/Sialic Acid Axis as a Central Glyco-Immune Checkpoint in Breast Cancer Bone Metastasis. Proc. Natl. Acad. Sci. USA 2024, 121, e2312929121. [Google Scholar] [CrossRef] [PubMed]
- Barkan, D.; Kleinman, H.; Simmons, J.L.; Asmussen, H.; Kamaraju, A.K.; Hoenorhoff, M.J.; Liu, Z.; Costes, S.V.; Cho, E.H.; Lockett, S.; et al. Inhibition of Metastatic Outgrowth from Single Dormant Tumor Cells by Targeting the Cytoskeleton. Cancer Res. 2008, 68, 6241–6250. [Google Scholar] [CrossRef]
- Adair, T.H.; Montani, J.-P. Overview of Angiogenesis. In Angiogenesis; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010. [Google Scholar]
- Folkman, J. Role of Angiogenesis in Tumor Growth and Metastasis. Semin. Oncol. 2002, 29, 15–18. [Google Scholar] [CrossRef]
- Saaristo, A.; Karpanen, T.; Alitalo, K. Mechanisms of Angiogenesis and Their Use in the Inhibition of Tumor Growth and Metastasis. Oncogene 2000, 19, 6122–6129. [Google Scholar] [CrossRef]
- Rak, J.W.; St Croix, B.D.; Kerbel, R.S. Consequences of Angiogenesis for Tumor Progression, Metastasis and Cancer Therapy. Anti-Cancer Drugs 1995, 6, 3. [Google Scholar] [CrossRef]
- Folkman, J. Tumor Angiogenesis: A Possible Control Point in Tumor Growth. Ann. Intern. Med. 1975, 82, 96–100. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis and Apoptosis. Semin. Cancer Biol. 2003, 13, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Naumov, G.N.; Bender, E.; Zurakowski, D.; Kang, S.-Y.; Sampson, D.; Flynn, E.; Watnick, R.S.; Straume, O.; Akslen, L.A.; Folkman, J.; et al. A Model of Human Tumor Dormancy: An Angiogenic Switch from the Nonangiogenic Phenotype. JNCI J. Natl. Cancer Inst. 2006, 98, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Li, X.; Xu, K.; Wang, R.; Guan, X. C-MYC Mediates the Crosstalk between Breast Cancer Cells and Tumor Microenvironment. Cell Commun. Signal. 2023, 21, 28. [Google Scholar] [CrossRef] [PubMed]
- Zangouei, A.S.; Zangoue, M.; Taghehchian, N.; Zangooie, A.; Reza Rahimi, H.; Saburi, E.; Sadat Alavi, M.; Moghbeli, M. Cell Cycle Related Long Non-Coding RNAs as the Critical Regulators of Breast Cancer Progression and Metastasis. Biol. Res. 2023, 56, 1. [Google Scholar] [CrossRef] [PubMed]
- Marvalim, C.; Datta, A.; Lee, S.C. Role of P53 in Breast Cancer Progression: An Insight into P53 Targeted Therapy. Theranostics 2023, 13, 1421–1442. [Google Scholar] [CrossRef] [PubMed]
- Bertram, J.S. The Molecular Biology of Cancer. Mol. Asp. Med. 2000, 21, 167–223. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, R.A.; Weinberg, R.A. The Biology of Cancer; W.W. Norton & Company: New York, NY, USA, 2013; ISBN 978-0-203-85256-9. [Google Scholar]
- Crespi, B.; Summers, K. Evolutionary Biology of Cancer. Trends Ecol. Evol. 2005, 20, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Eroles, P.; Bosch, A.; Alejandro Pérez-Fidalgo, J.; Lluch, A. Molecular Biology in Breast Cancer: Intrinsic Subtypes and Signaling Pathways. Cancer Treat. Rev. 2012, 38, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Nolan, E.; Lindeman, G.J.; Visvader, J.E. Deciphering Breast Cancer: From Biology to the Clinic. Cell 2023, 186, 1708–1728. [Google Scholar] [CrossRef]
- MacKie, R.M.; Reid, R.; Junor, B. Fatal Melanoma Transferred in a Donated Kidney 16 Years after Melanoma Surgery. N. Engl. J. Med. 2003, 348, 567–568. [Google Scholar] [CrossRef]
- Suranyi, M.G.; Hogan, P.G.; Falk, M.C.; Axelsen, R.A.; Rigby, R.; Hawley, C.; Petrie, J. ADVANCED DONOR-ORIGIN MELANOMA IN A RENAL TRANSPLANT RECIPIENT: Immunotherapy, Cure, and Retransplantation. Transplantation 1998, 66, 655. [Google Scholar] [CrossRef] [PubMed]
- Fattouh, K.; Ducroux, E.; Decullier, E.; Kanitakis, J.; Morelon, E.; Boissonnat, P.; Sebbag, L.; Jullien, D.; Euvrard, S. Increasing Incidence of Melanoma after Solid Organ Transplantation: A Retrospective Epidemiological Study. Transpl. Int. 2017, 30, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Granata, S.; Tessari, G.; Stallone, G.; Zaza, G. Skin Cancer in Solid Organ Transplant Recipients: Still an Open Problem. Front. Med. 2023, 10, 1189680. [Google Scholar] [CrossRef]
- Rosales, B.M.; Hedley, J.; De La Mata, N.; Cavazzoni, E.; Vajdic, C.M.; Thompson, J.F.; Kelly, P.J.; Wyburn, K.; Webster, A.C. Transmission and Non-Transmission of Melanoma from Deceased Solid Organ Donors to Transplant Recipients: Risks and Missed Opportunities. Transplantation 2024, 108, 1623–1631. [Google Scholar] [CrossRef] [PubMed]
- Wenande, E.; Togsverd-Bo, K.; Hastrup, A.; Lei, U.; Philipsen, P.A.; Haedersdal, M. Skin Cancer Development Is Strongly Associated with Actinic Keratosis in Solid Organ Transplant Recipients: A Danish Cohort Study. Dermatology 2023, 239, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Kulbat, A.; Richter, K.; Krzysztofik, M.; Batko, K.; Karwańska, A.; Kołodziej-Rzepa, M.; Wojewoda, T.; Wysocki, W.M. Melanoma Incidence in 17,252 Organ Transplant Recipients in Poland in 2010–2022. Nowotw. J. Oncol. 2024, 74, 173–179. [Google Scholar] [CrossRef]
- Das, U.; Banik, S.; Nadumane, S.S.; Chakrabarti, S.; Gopal, D.; Kabekkodu, S.P.; Srisungsitthisunti, P.; Mazumder, N.; Biswas, R. Isolation, Detection and Analysis of Circulating Tumour Cells: A Nanotechnological Bioscope. Pharmaceutics 2023, 15, 280. [Google Scholar] [CrossRef] [PubMed]
- Vishnoi, M.; Peddibhotla, S.; Yin, W.; Scamardo, A.T.; George, G.C.; Hong, D.S.; Marchetti, D. The Isolation and Characterization of CTC Subsets Related to Breast Cancer Dormancy. Sci. Rep. 2015, 5, 17533. [Google Scholar] [CrossRef] [PubMed]
- Truskowski, K.; Amend, S.R.; Pienta, K.J. Dormant Cancer Cells: Programmed Quiescence, Senescence, or Both? Cancer Metastasis Rev. 2023, 42, 37–47. [Google Scholar] [CrossRef]
- Nasr, M.M.; Lynch, C.C. How Circulating Tumor Cluster Biology Contributes to the Metastatic Cascade: From Invasion to Dissemination and Dormancy. Cancer Metastasis Rev. 2023, 42, 1133–1146. [Google Scholar] [CrossRef]
- Min, H.-Y.; Lee, H.-Y. Cellular Dormancy in Cancer: Mechanisms and Potential Targeting Strategies. Cancer Res. Treat. 2023, 55, 720–736. [Google Scholar] [CrossRef]
- Wu, S.; Zhao, S.; Cui, D.; Xie, J. Advances in the Biology, Detection Techniques, and Clinical Applications of Circulating Tumor Cells. J. Oncol. 2022, 2022, 7149686. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.M.; Chen, C.S. Deconstructing the Third Dimension–How 3D Culture Microenvironments Alter Cellular Cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef]
- Rijal, G.; Li, W. A Versatile 3D Tissue Matrix Scaffold System for Tumor Modeling and Drug Screening. Sci. Adv. 2017, 3, e1700764. [Google Scholar] [CrossRef]
- Ghajar, C.M.; Peinado, H.; Mori, H.; Matei, I.R.; Evason, K.J.; Brazier, H.; Almeida, D.; Koller, A.; Hajjar, K.A.; Stainier, D.Y.R.; et al. The Perivascular Niche Regulates Breast Tumour Dormancy. Nat. Cell Biol. 2013, 15, 807–817. [Google Scholar] [CrossRef]
- Ren, Q.; Khoo, W.H.; Corr, A.P.; Phan, T.G.; Croucher, P.I.; Stewart, S.A. Gene Expression Predicts Dormant Metastatic Breast Cancer Cell Phenotype. Breast Cancer Res. 2022, 24, 10. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-Cell RNA-Seq Highlights Intratumoral Heterogeneity in Primary Glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef]
- Navin, N.; Kendall, J.; Troge, J.; Andrews, P.; Rodgers, L.; McIndoo, J.; Cook, K.; Stepansky, A.; Levy, D.; Esposito, D.; et al. Tumour Evolution Inferred by Single-Cell Sequencing. Nature 2011, 472, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, D.; Peng, M.; Tang, L.; Ouyang, J.; Xiong, F.; Guo, C.; Tang, Y.; Zhou, Y.; Liao, Q.; et al. Single-cell RNA Sequencing in Cancer Research. J. Exp. Clin. Cancer Res. 2021, 40, 81. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Waters, J.; Leung, M.L.; Unruh, A.; Roh, W.; Shi, X.; Chen, K.; Scheet, P.; Vattathil, S.; Liang, H.; et al. Clonal Evolution in Breast Cancer Revealed by Single Nucleus Genome Sequencing. Nature 2014, 512, 155–160. [Google Scholar] [CrossRef]
- Kedrin, D.; Gligorijevic, B.; Wyckoff, J.; Verkhusha, V.V.; Condeelis, J.; Segall, J.E.; van Rheenen, J. Intravital Imaging of Metastatic Behavior through a Mammary Imaging Window. Nat. Methods 2008, 5, 1019–1021. [Google Scholar] [CrossRef] [PubMed]
- Ellenbroek, S.I.J.; van Rheenen, J. Imaging Hallmarks of Cancer in Living Mice. Nat. Rev. Cancer 2014, 14, 406–418. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, S.; Bhoopathi, P.; Emdad, L.; Das, S.; Sarkar, D.; Fisher, P.B. Chapter Two-Dormancy and Cancer Stem Cells: An Enigma for Cancer Therapeutic Targeting. In Advances in Cancer Research; Civin, C.I., Kingsbury, T.J., Kim, M., Fisher, P.B., Eds.; Cancer Stem Cells; Academic Press: Cambridge, MA, USA, 2019; Volume 141, pp. 43–84. [Google Scholar]
- Aguirre-Ghiso, J.A. Translating the Science of Cancer Dormancy to the Clinic. Cancer Res. 2021, 81, 4673–4675. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Croucher, P.I. The Dormant Cancer Cell Life Cycle. Nat. Rev. Cancer 2020, 20, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Morales-Valencia, J.; David, G. The Origins of Cancer Cell Dormancy. Curr. Opin. Genet. Dev. 2022, 74, 101914. [Google Scholar] [CrossRef]
- Risson, E.; Nobre, A.R.; Maguer-Satta, V.; Aguirre-Ghiso, J.A. The Current Paradigm and Challenges Ahead for the Dormancy of Disseminated Tumor Cells. Nat. Cancer 2020, 1, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, A.I.; Trinidad, J.R.; Sandiford, O.; Etchegaray, J.-P.; Rameshwar, P. Epigenetic Dynamics in Cancer Stem Cell Dormancy. Cancer Metastasis Rev. 2020, 39, 721–738. [Google Scholar] [CrossRef] [PubMed]
- Summers, M.A.; McDonald, M.M.; Croucher, P.I. Cancer Cell Dormancy in Metastasis. Cold Spring Harb. Perspect. Med. 2020, 10, a037556. [Google Scholar] [CrossRef] [PubMed]
- Rack, B.K.; Schindlbeck, C.; Andergassen, U.; Schneeweiss, A.; Zwingers, T.; Lichtenegger, W.; Beckmann, M.; Sommer, H.L.; Pantel, K.; Janni, W.; et al. Use of Circulating Tumor Cells (CTC) in Peripheral Blood of Breast Cancer Patients before and after Adjuvant Chemotherapy to Predict Risk for Relapse: The SUCCESS Trial. J. Clin. Oncol. 2010, 28, 1003. [Google Scholar] [CrossRef]
- Becker, S.; Solomayer, E.; Becker-Pergola, G.; Wallwiener, D.; Fehm, T. Primary Systemic Therapy Does Not Eradicate Disseminated Tumor Cells in Breast Cancer Patients. Breast Cancer Res. Treat. 2007, 106, 239–243. [Google Scholar] [CrossRef]
- Damen, M.P.F.; van Rheenen, J.; Scheele, C.L.G.J. Targeting Dormant Tumor Cells to Prevent Cancer Recurrence. FEBS J. 2021, 288, 6286–6303. [Google Scholar] [CrossRef] [PubMed]
- Sauer, S.; Reed, D.R.; Ihnat, M.; Hurst, R.E.; Warshawsky, D.; Barkan, D. Innovative Approaches in the Battle Against Cancer Recurrence: Novel Strategies to Combat Dormant Disseminated Tumor Cells. Front. Oncol. 2021, 11, 659963. [Google Scholar] [CrossRef] [PubMed]
- Francescangeli, F.; De Angelis, M.L.; Rossi, R.; Cuccu, A.; Giuliani, A.; De Maria, R.; Zeuner, A. Dormancy, Stemness, and Therapy Resistance: Interconnected Players in Cancer Evolution. Cancer Metastasis Rev. 2023, 42, 197–215. [Google Scholar] [CrossRef] [PubMed]
- Triana-Martínez, F.; Loza, M.I.; Domínguez, E. Beyond Tumor Suppression: Senescence in Cancer Stemness and Tumor Dormancy. Cells 2020, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Sistigu, A.; Musella, M.; Galassi, C.; Vitale, I.; De Maria, R. Tuning Cancer Fate: Tumor Microenvironment’s Role in Cancer Stem Cell Quiescence and Reawakening. Front. Immunol. 2020, 11, 2166. [Google Scholar] [CrossRef] [PubMed]
- Bakhshandeh, S.; Werner, C.; Fratzl, P.; Cipitria, A. Microenvironment-Mediated Cancer Dormancy: Insights from Metastability Theory. Proc. Natl. Acad. Sci. USA 2022, 119, e2111046118. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Bui, T.; Muller, W.J. Exploiting Mouse Models to Recapitulate Clinical Tumor Dormancy and Recurrence in Breast Cancer. Endocrinology 2022, 163, bqac055. [Google Scholar] [CrossRef] [PubMed]
- Burstein, H.J.; Lacchetti, C.; Anderson, H.; Buchholz, T.A.; Davidson, N.E.; Gelmon, K.A.; Giordano, S.H.; Hudis, C.A.; Solky, A.J.; Stearns, V.; et al. Adjuvant Endocrine Therapy for Women with Hormone Receptor–Positive Breast Cancer: ASCO Clinical Practice Guideline Focused Update. J. Clin. Oncol. 2019, 37, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.; Kyriakides, S.; Ohno, S.; Penault-Llorca, F.; Poortmans, P.; Rubio, I.T.; Zackrisson, S.; Senkus, E. Early Breast Cancer: ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up. Ann. Oncol. 2019, 30, 1194–1220. [Google Scholar] [CrossRef]
- Davies, C.; Pan, H.; Godwin, J.; Gray, R.; Arriagada, R.; Raina, V.; Abraham, M.; Alencar, V.H.M.; Badran, A.; Bonfill, X.; et al. Long-Term Effects of Continuing Adjuvant Tamoxifen to 10 Years versus Stopping at 5 Years after Diagnosis of Oestrogen Receptor-Positive Breast Cancer: ATLAS, a Randomised Trial. Lancet 2013, 381, 805–816. [Google Scholar] [CrossRef]
- Early Breast Cancer Trialists’ Collaborative Group (EBCTCG). Aromatase Inhibitors versus Tamoxifen in Early Breast Cancer: Patient-Level Meta-Analysis of the Randomised Trials. Lancet 2015, 386, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Kuźnik, A.; Październiok-Holewa, A.; Jewula, P.; Kuźnik, N. Bisphosphonates—Much More than Only Drugs for Bone Diseases. Eur. J. Pharmacol. 2020, 866, 172773. [Google Scholar] [CrossRef] [PubMed]
- Drake, M.T.; Clarke, B.L.; Khosla, S. Bisphosphonates: Mechanism of Action and Role in Clinical Practice. Mayo Clin. Proc. 2008, 83, 1032–1045. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.D.; Watts, N.B. Bisphosphonates. In Osteoporosis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 123–143. ISBN 978-1-118-31629-0. [Google Scholar]
- Center, J.R.; Lyles, K.W.; Bliuc, D. Bisphosphonates and Lifespan. Bone 2020, 141, 115566. [Google Scholar] [CrossRef] [PubMed]
- Gralow, J.R.; Barlow, W.E.; Paterson, A.H.G.; M’iao, J.L.; Lew, D.L.; Stopeck, A.T.; Hayes, D.F.; Hershman, D.L.; Schubert, M.M.; Clemons, M.; et al. Phase III Randomized Trial of Bisphosphonates as Adjuvant Therapy in Breast Cancer: S0307. JNCI J. Natl. Cancer Inst. 2020, 112, 698–707. [Google Scholar] [CrossRef]
- Gnant, M.; Pfeiler, G.; Steger, G.G.; Egle, D.; Greil, R.; Fitzal, F.; Wette, V.; Balic, M.; Haslbauer, F.; Melbinger-Zeinitzer, E.; et al. Adjuvant Denosumab in Postmenopausal Patients with Hormone Receptor-Positive Breast Cancer (ABCSG-18): Disease-Free Survival Results from a Randomised, Double-Blind, Placebo-Controlled, Phase 3 Trial. Lancet Oncol. 2019, 20, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, T.; Teufel, I.; Geiger, K.; Vater, Y.; Aicher, W.K.; Klein, G.; Fehm, T. Bisphosphonates Modulate Vital Functions of Human Osteoblasts and Affect Their Interactions with Breast Cancer Cells. Breast Cancer Res. Treat. 2013, 140, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.; Ezratty, C.; Mhango, G.; Lin, J.J. Clinical and Sociodemographic Risk Factors Associated with the Development of Second Primary Cancers among Postmenopausal Breast Cancer Survivors. Breast Cancer 2023, 30, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Holen, I.; Coleman, R.E. Anti-Tumour Activity of Bisphosphonates in Preclinical Models of Breast Cancer. Breast Cancer Res. 2010, 12, 214. [Google Scholar] [CrossRef]
- Clézardin, P.; Ebetino, F.H.; Fournier, P.G.J. Bisphosphonates and Cancer-Induced Bone Disease: Beyond Their Antiresorptive Activity. Cancer Res. 2005, 65, 4971–4974. [Google Scholar] [CrossRef]
- Bedard, P.L.; Body, J.-J.; Piccart-Gebhart, M.J. Sowing the Soil for Cure? Results of the ABCSG-12 Trial Open a New Chapter in the Evolving Adjuvant Bisphosphonate Story in Early Breast Cancer. J. Clin. Oncol. 2009, 27, 4043–4046. [Google Scholar] [CrossRef] [PubMed]
- Wiemer, A.J.; Hohl, R.J.; Wiemer, D.F. The Intermediate Enzymes of Isoprenoid Metabolism as Anticancer Targets. Anti-Cancer Agents Med. Chem. 2009, 9, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Stresing, V.; Fournier, P.G.; Bellahcène, A.; Benzaïd, I.; Mönkkönen, H.; Colombel, M.; Ebetino, F.H.; Castronovo, V.; Clézardin, P. Nitrogen-Containing Bisphosphonates Can Inhibit Angiogenesis in Vivo without the Involvement of Farnesyl Pyrophosphate Synthase. Bone 2011, 48, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Benzaïd, I.; Mönkkönen, H.; Stresing, V.; Bonnelye, E.; Green, J.; Mönkkönen, J.; Touraine, J.-L.; Clézardin, P. High Phosphoantigen Levels in Bisphosphonate-Treated Human Breast Tumors Promote Vgamma9Vdelta2 T-Cell Chemotaxis and Cytotoxicity in Vivo. Cancer Res. 2011, 71, 4562–4572. [Google Scholar] [CrossRef] [PubMed]
- Winter, M.C.; Holen, I.; Coleman, R.E. Exploring the Anti-Tumour Activity of Bisphosphonates in Early Breast Cancer. Cancer Treat. Rev. 2008, 34, 453–475. [Google Scholar] [CrossRef] [PubMed]
- Gnant, M.; Clézardin, P. Direct and Indirect Anticancer Activity of Bisphosphonates: A Brief Review of Published Literature. Cancer Treat. Rev. 2012, 38, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.X. Bevacizumab and Breast Cancer: Current Therapeutic Progress and Future Perspectives. Expert Rev. Anticancer Ther. 2009, 9, 1715–1725. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Wu, L.; Chen, J.; Na, N.; Lou, G. Neoadjuvant Nivolumab plus Bevacizumab Therapy Improves the Prognosis of Triple-Negative Breast Cancer in Humanized Mouse Models. Breast Cancer 2024, 31, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, P.; Xiang, F. Efficacy Evaluation of Bevacizumab Combined with Capecitabine in the Treatment of HER2-Negative Metastatic Breast Cancer: A Meta-Analysis. J. Oncol. 2023, 2023, e8740221. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Yamashiro, H.; Schneeweiss, A.; Müller, V.; Gluz, O.; Klare, P.; Aktas, B.; Magdolna, D.; Büdi, L.; Pikó, B.; et al. Factors Affecting Prognosis in Patients Treated with Bevacizumab plus Paclitaxel as First-Line Chemotherapy for HER2-Negative Metastatic Breast Cancer: An International Pooled Analysis of Individual Patient Data from Four Prospective Observational Studies. Breast Cancer 2023, 30, 88–100. [Google Scholar] [CrossRef]
- Matsui, K.; Earashi, M.; Yoshikawa, A.; Fukushima, W.; Nozaki, Z.; Oyama, K.; Maeda, K.; Nakakura, A.; Morita, S.; Fujii, T. Real-world Effect of Bevacizumab and Eribulin on Metastatic Breast Cancer Using a Propensity Score Matching Analysis. Mol. Clin. Oncol. 2023, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.L.; Tayob, N.; Ren, S.; Savoie, J.J.; Spigel, D.R.; Burris, H.A.; Ryan, P.D.; Harris, L.N.; Winer, E.P.; Burstein, H.J. A Randomized Phase II Study of Metronomic Cyclophosphamide and Methotrexate (CM) with or without Bevacizumab in Patients with Advanced Breast Cancer. Breast Cancer Res. Treat. 2024, 204, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Miles, D.W.; Diéras, V.; Cortés, J.; Duenne, A.-A.; Yi, J.; O’Shaughnessy, J. First-Line Bevacizumab in Combination with Chemotherapy for HER2-Negative Metastatic Breast Cancer: Pooled and Subgroup Analyses of Data from 2447 Patients. Ann. Oncol. 2013, 24, 2773–2780. [Google Scholar] [CrossRef] [PubMed]
- Romond, E.H.; Perez, E.A.; Bryant, J.; Suman, V.J.; Geyer, C.E., Jr.; Davidson, N.E.; Tan-Chiu, E.; Martino, S.; Paik, S.; Kaufman, P.A.; et al. Trastuzumab plus Adjuvant Chemotherapy for Operable HER2-Positive Breast Cancer. N. Engl. J. Med. 2005, 353, 1673–1684. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.; Braybrooke, J.; Gray, R.; Hills, R.; Liu, Z.; Peto, R.; Davies, L.; Dodwell, D.; McGale, P.; Pan, H.; et al. Trastuzumab for Early-Stage, HER2-Positive Breast Cancer: A Meta-Analysis of 13 864 Women in Seven Randomised Trials. Lancet Oncol. 2021, 22, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- von Minckwitz, G.; Huang, C.-S.; Mano, M.S.; Loibl, S.; Mamounas, E.P.; Untch, M.; Wolmark, N.; Rastogi, P.; Schneeweiss, A.; Redondo, A.; et al. Trastuzumab Emtansine for Residual Invasive HER2-Positive Breast Cancer. N. Engl. J. Med. 2019, 380, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Viani, G.A.; Afonso, S.L.; Stefano, E.J.; De Fendi, L.I.; Soares, F.V. Adjuvant Trastuzumab in the Treatment of Her-2-Positive Early Breast Cancer: A Meta-Analysis of Published Randomized Trials. BMC Cancer 2007, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Shi, Y.; Yan, R.; Yin, S.; Bu, H.; Huang, J. Efficacy and Safety of Trastuzumab Deruxtecan in Treating Human Epidermal Growth Factor Receptor 2-Low/Positive Advanced Breast Cancer:A Meta-Analysis of Randomized Controlled Trials. Crit. Rev. Oncol./Hematol. 2024, 196, 104305. [Google Scholar] [CrossRef] [PubMed]
- Falcón González, A.; Cruz Jurado, J.; Llabrés Valenti, E.; Urbano Cubero, R.; Álamo de la Gala, M.C.; Martínez Guisado, M.A.; Álvarez Ambite, R.; Rodríguez González, C.J.; Amérigo Góngora, M.; Rodríguez Pérez, L.; et al. Real-World Experience with Pertuzumab and Trastuzumab Combined with Chemotherapy in Neoadjuvant Treatment for Patients with Early-Stage HER2-Positive Breast Cancer: The NEOPERSUR Study. Clin. Transl. Oncol. 2024. [Google Scholar] [CrossRef]
- Huang, L.; Pang, D.; Yang, H.; Li, W.; Wang, S.; Cui, S.; Liao, N.; Wang, Y.; Wang, C.; Chang, Y.-C.; et al. Neoadjuvant–Adjuvant Pertuzumab in HER2-Positive Early Breast Cancer: Final Analysis of the Randomized Phase III PEONY Trial. Nat. Commun. 2024, 15, 2153. [Google Scholar] [CrossRef]
- Sharman Moser, S.; Apter, L.; Livnat, I.; Ginsburg, R.; Yarden, A.; Drori, M.; Drizon, A.; Chodick, G.; Siegelmann-Danieli, N. Clinical Outcomes of Patients with HER2 Positive Metastatic Breast Cancer to the Brain, with First-Line Trastuzumab, Pertuzumab and Chemotherapy, in a Real-World Setting. Breast Cancer Targets Ther. 2024, 16, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Godolphin, W.; Jones, L.A.; Holt, J.A.; Wong, S.G.; Keith, D.E.; Levin, W.J.; Stuart, S.G.; Udove, J.; Ullrich, A.; et al. Studies of the HER-2/Neu Proto-Oncogene in Human Breast and Ovarian Cancer. Science 1989, 244, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, N.; Banys, M.; Neubauer, H.; Solomayer, E.-F.; Gall, C.; Hahn, M.; Becker, S.; Bachmann, R.; Wallwiener, D.; Fehm, T. HER2 Status on Persistent Disseminated Tumor Cells after Adjuvant Therapy May Differ from Initial HER2 Status on Primary Tumor. Anticancer Res. 2009, 29, 4019–4024. [Google Scholar] [PubMed]
- Meng, S.; Tripathy, D.; Shete, S.; Ashfaq, R.; Haley, B.; Perkins, S.; Beitsch, P.; Khan, A.; Euhus, D.; Osborne, C.; et al. HER-2 Gene Amplification Can Be Acquired as Breast Cancer Progresses. Proc. Natl. Acad. Sci. USA 2004, 101, 9393–9398. [Google Scholar] [CrossRef] [PubMed]
- Jueckstock, J.; Rack, B.; Schindlbeck, C.; Hofmann, S.; Zill, B.; Mylonas, I.; Blankenstein, T.; Janni, W.; Friese, K. Treatment with Trastuzumab in Recurrence Free Patients with Early Breast Cancer and Persistent Disseminated Tumor Cells (DTC) in Bone Marrow. Cancer Res. 2009, 69, 3135. [Google Scholar] [CrossRef]
- Saxena, M.; van der Burg, S.H.; Melief, C.J.M.; Bhardwaj, N. Therapeutic Cancer Vaccines. Nat. Rev. Cancer 2021, 21, 360–378. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.J.; Svensson-Arvelund, J.; Lubitz, G.S.; Marabelle, A.; Melero, I.; Brown, B.D.; Brody, J.D. Cancer Vaccines: The next Immunotherapy Frontier. Nat. Cancer 2022, 3, 911–926. [Google Scholar] [CrossRef] [PubMed]
- Vajari, M.K.; Sanaei, M.-J.; Salari, S.; Rezvani, A.; Ravari, M.S.; Bashash, D. Breast Cancer Vaccination: Latest Advances with an Analytical Focus on Clinical Trials. Int. Immunopharmacol. 2023, 123, 110696. [Google Scholar] [CrossRef] [PubMed]
- Debien, V.; De Caluwé, A.; Wang, X.; Piccart-Gebhart, M.; Tuohy, V.K.; Romano, E.; Buisseret, L. Immunotherapy in Breast Cancer: An Overview of Current Strategies and Perspectives. npj Breast Cancer 2023, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Schlom, J. Therapeutic Cancer Vaccines: Current Status and Moving Forward. JNCI J. Natl. Cancer Inst. 2012, 104, 599–613. [Google Scholar] [CrossRef]
- Ubowski, M.M.; VanSice, R.; Marriott, M.; Yacobucci, M.J.; Chablani, L. Amplifying Immune Responses: Microparticulate Vaccine Approach Against Breast Cancer. Breast Cancer Targets Ther. 2024, 16, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Morse, M.A.; Crosby, E.J.; Force, J.; Osada, T.; Hobeika, A.C.; Hartman, Z.C.; Berglund, P.; Smith, J.; Lyerly, H.K. Clinical Trials of Self-Replicating RNA-Based Cancer Vaccines. Cancer Gene Ther. 2023, 30, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Disis, M.L.; Guthrie, K.A.; Liu, Y.; Coveler, A.L.; Higgins, D.M.; Childs, J.S.; Dang, Y.; Salazar, L.G. Safety and Outcomes of a Plasmid DNA Vaccine Encoding the ERBB2 Intracellular Domain in Patients with Advanced-Stage ERBB2-Positive Breast Cancer: A Phase 1 Nonrandomized Clinical Trial. JAMA Oncol. 2023, 9, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ko, B.K.; Kawano, K.; Murray, J.L.; Disis, M.L.; Efferson, C.L.; Kuerer, H.M.; Peoples, G.E.; Ioannides, C.G. Clinical Studies of Vaccines Targeting Breast Cancer1. Clin. Cancer Res. 2003, 9, 3222–3234. [Google Scholar] [PubMed]
- Curigliano, G.; Spitaleri, G.; Pietri, E.; Rescigno, M.; de Braud, F.; Cardillo, A.; Munzone, E.; Rocca, A.; Bonizzi, G.; Brichard, V.; et al. Breast Cancer Vaccines: A Clinical Reality or Fairy Tale? Ann. Oncol. 2006, 17, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.E.; Dowsett, M. Aromatase Inhibitors in Breast Cancer. N. Engl. J. Med. 2003, 348, 2431–2442. [Google Scholar] [CrossRef] [PubMed]
- Cuzick, J.; Powles, T.; Veronesi, U.; Forbes, J.; Edwards, R.; Ashley, S.; Boyle, P. Overview of the Main Outcomes in Breast-Cancer Prevention Trials. Lancet 2003, 361, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Howell, A.; Cuzick, J.; Baum, M.; Buzdar, A.; Dowsett, M.; Forbes, J.F.; Hoctin-Boes, G.; Houghton, J.; Locker, G.Y.; Tobias, J.S.; et al. Results of the ATAC (Arimidex, Tamoxifen, Alone or in Combination) Trial after Completion of 5 Years’ Adjuvant Treatment for Breast Cancer. Lancet 2005, 365, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Di Leo, A.; Jerusalem, G.; Petruzelka, L.; Torres, R.; Bondarenko, I.N.; Khasanov, R.; Verhoeven, D.; Pedrini, J.L.; Smirnova, I.; Lichinitser, M.R.; et al. Final Overall Survival: Fulvestrant 500 Mg vs 250 Mg in the Randomized CONFIRM Trial. J. Natl. Cancer Inst. 2014, 106, djt337. [Google Scholar] [CrossRef]
- Adams, S.; Loi, S.; Toppmeyer, D.; Cescon, D.W.; Laurentiis, M.D.; Nanda, R.; Winer, E.P.; Mukai, H.; Tamura, K.; Armstrong, A.; et al. Pembrolizumab Monotherapy for Previously Untreated, PD-L1-Positive, Metastatic Triple-Negative Breast Cancer: Cohort B of the Phase II KEYNOTE-086 Study. Ann. Oncol. 2019, 30, 405–411. [Google Scholar] [CrossRef]
- Schmid, P.; Cortes, J.; Pusztai, L.; McArthur, H.; Kümmel, S.; Bergh, J.; Denkert, C.; Park, Y.H.; Hui, R.; Harbeck, N.; et al. Pembrolizumab for Early Triple-Negative Breast Cancer. N. Engl. J. Med. 2020, 382, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Robson, M.; Im, S.-A.; Senkus, E.; Xu, B.; Domchek, S.M.; Masuda, N.; Delaloge, S.; Li, W.; Tung, N.; Armstrong, A.; et al. Olaparib for Metastatic Breast Cancer in Patients with a Germline BRCA Mutation. N. Engl. J. Med. 2017, 377, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Litton, J.K.; Rugo, H.S.; Ettl, J.; Hurvitz, S.A.; Gonçalves, A.; Lee, K.-H.; Fehrenbacher, L.; Yerushalmi, R.; Mina, L.A.; Martin, M.; et al. Talazoparib in Patients with Advanced Breast Cancer and a Germline BRCA Mutation. N. Engl. J. Med. 2018, 379, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Hortobagyi, G.N.; Stemmer, S.M.; Burris, H.A.; Yap, Y.-S.; Sonke, G.S.; Paluch-Shimon, S.; Campone, M.; Blackwell, K.L.; André, F.; Winer, E.P.; et al. Ribociclib as First-Line Therapy for HR-Positive, Advanced Breast Cancer. N. Engl. J. Med. 2016, 375, 1738–1748. [Google Scholar] [CrossRef] [PubMed]
- Sledge, G.W.; Toi, M.; Neven, P.; Sohn, J.; Inoue, K.; Pivot, X.; Burdaeva, O.; Okera, M.; Masuda, N.; Kaufman, P.A.; et al. MONARCH 2: Abemaciclib in Combination with Fulvestrant in Women With HR+/HER2- Advanced Breast Cancer Who Had Progressed While Receiving Endocrine Therapy. J. Clin. Oncol. 2017, 35, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.; Pearson, A.; Sharpe, R.; Lambros, M.; Geyer, F.; Lopez-Garcia, M.A.; Natrajan, R.; Marchio, C.; Iorns, E.; Mackay, A.; et al. FGFR1 Amplification Drives Endocrine Therapy Resistance and Is a Therapeutic Target in Breast Cancer. Cancer Res. 2010, 70, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Baylin, S.B. The Epigenomics of Cancer. Cell 2007, 128, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Stefansson, O.A.; Esteller, M. Epigenetic Modifications in Breast Cancer and Their Role in Personalized Medicine. Am. J. Pathol. 2013, 183, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Baylin, S.B.; Jones, P.A. A Decade of Exploring the Cancer Epigenome—Biological and Translational Implications. Nat. Rev. Cancer 2011, 11, 726–734. [Google Scholar] [CrossRef]
- Toyota, M.; Issa, J.-P.J. CpG Island Methylator Phenotypes in Aging and Cancer. Semin. Cancer Biol. 1999, 9, 349–357. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Schwaller, J. Learning from Mouse Models of MLL Fusion Gene-Driven Acute Leukemia. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2020, 1863, 194550. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.A.; Kouzarides, T. Cancer Epigenetics: From Mechanism to Therapy. Cell 2012, 150, 12–27. [Google Scholar] [CrossRef]
- Chi, P.; Allis, C.D.; Wang, G.G. Covalent Histone Modifications—Miswritten, Misinterpreted and Mis-Erased in Human Cancers. Nat. Rev. Cancer 2010, 10, 457–469. [Google Scholar] [CrossRef]
- Clapier, C.R.; Cairns, B.R. The Biology of Chromatin Remodeling Complexes. Annu. Rev. Biochem. 2009, 78, 273–304. [Google Scholar] [CrossRef] [PubMed]
- Lusser, A.; Kadonaga, J.T. Chromatin Remodeling by ATP-Dependent Molecular Machines. BioEssays 2003, 25, 1192–1200. [Google Scholar] [CrossRef]
- Hargreaves, D.C.; Crabtree, G.R. ATP-Dependent Chromatin Remodeling: Genetics, Genomics and Mechanisms. Cell Res. 2011, 21, 396–420. [Google Scholar] [CrossRef]
- Wood, L.D.; Parsons, D.W.; Jones, S.; Lin, J.; Sjöblom, T.; Leary, R.J.; Shen, D.; Boca, S.M.; Barber, T.; Ptak, J.; et al. The Genomic Landscapes of Human Breast and Colorectal Cancers. Science 2007, 318, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.M.; Eun Kwon, J.; Ali, T.; Jang, J.; Ullah, I.; Lee, Y.-G.; Park, D.W.; Park, J.; Jeang, J.W.; Kang, S.C. Triple-Negative Breast Cancer: Epidemiology, Molecular Mechanisms, and Modern Vaccine-Based Treatment Strategies. Biochem. Pharmacol. 2023, 212, 115545. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, G.; Heard, E. Advances in Epigenetics Link Genetics to the Environment and Disease. Nature 2019, 571, 489–499. [Google Scholar] [CrossRef]
- Bouyahya, A.; El Hachlafi, N.; Aanniz, T.; Bourais, I.; Mechchate, H.; Benali, T.; Shariati, M.A.; Burkov, P.; Lorenzo, J.M.; Wilairatana, P.; et al. Natural Bioactive Compounds Targeting Histone Deacetylases in Human Cancers: Recent Updates. Molecules 2022, 27, 2568. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Yang, Y.; Wang, Y. Predictive Biomarkers and Potential Drug Combinations of Epi-Drugs in Cancer Therapy. Clin. Epigenet. 2021, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Rosano, D.; Sofyali, E.; Dhiman, H.; Ghirardi, C.; Ivanoiu, D.; Heide, T.; Vingiani, A.; Bertolotti, A.; Pruneri, G.; Canale, E.; et al. Long-Term Multimodal Recording Reveals Epigenetic Adaptation Routes in Dormant Breast Cancer Cells. Cancer Discov. 2024, 14, 866–889. [Google Scholar] [CrossRef] [PubMed]
- Mabe, N.W.; Garcia, N.M.G.; Wolery, S.E.; Newcomb, R.; Meingasner, R.C.; Vilona, B.A.; Lupo, R.; Lin, C.-C.; Chi, J.-T.; Alvarez, J.V. G9a Promotes Breast Cancer Recurrence through Repression of a Pro-Inflammatory Program. Cell Rep. 2020, 33, 108341. [Google Scholar] [CrossRef] [PubMed]
- Poulard, C.; Noureddine, L.M.; Pruvost, L.; Le Romancer, M. Structure, Activity, and Function of the Protein Lysine Methyltransferase G9a. Life 2021, 11, 1082. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, C.A.; Eisenberg, L.M. G9a and G9a-Like Histone Methyltransferases and Their Effect on Cell Phenotype, Embryonic Development, and Human Disease. In The DNA, RNA, and Histone Methylomes; Jurga, S., Barciszewski, J., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 399–433. ISBN 978-3-030-14792-1. [Google Scholar]
- Lavin, D.P.; Tiwari, V.K. Unresolved Complexity in the Gene Regulatory Network Underlying EMT. Front. Oncol. 2020, 10, 554. [Google Scholar] [CrossRef] [PubMed]
- Nagrath, S.; Sequist, L.V.; Maheswaran, S.; Bell, D.W.; Irimia, D.; Ulkus, L.; Smith, M.R.; Kwak, E.L.; Digumarthy, S.; Muzikansky, A.; et al. Isolation of Rare Circulating Tumour Cells in Cancer Patients by Microchip Technology. Nature 2007, 450, 1235–1239. [Google Scholar] [CrossRef]
- Yu, M.; Bardia, A.; Wittner, B.S.; Stott, S.L.; Smas, M.E.; Ting, D.T.; Isakoff, S.J.; Ciciliano, J.C.; Wells, M.N.; Shah, A.M.; et al. Circulating Breast Tumor Cells Exhibit Dynamic Changes in Epithelial and Mesenchymal Composition. Science 2013, 339, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Stott, S.; Toner, M.; Maheswaran, S.; Haber, D.A. Circulating Tumor Cells: Approaches to Isolation and Characterization. J. Cell Biol. 2011, 192, 373–382. [Google Scholar] [CrossRef]
- Cristofanilli, M.; Budd, G.T.; Ellis, M.J.; Stopeck, A.; Matera, J.; Miller, M.C.; Reuben, J.M.; Doyle, G.V.; Allard, W.J.; Terstappen, L.W.M.M.; et al. Circulating Tumor Cells, Disease Progression, and Survival in Metastatic Breast Cancer. N. Engl. J. Med. 2004, 351, 781–791. [Google Scholar] [CrossRef]
- Diaz, L.A.; Bardelli, A. Liquid Biopsies: Genotyping Circulating Tumor DNA. J. Clin. Oncol. 2014, 32, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Hoon, D.S.B.; Pantel, K. Cell-Free Nucleic Acids as Biomarkers in Cancer Patients. Nat. Rev. Cancer 2011, 11, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Siravegna, G.; Marsoni, S.; Siena, S.; Bardelli, A. Integrating Liquid Biopsies into the Management of Cancer. Nat. Rev. Clin. Oncol. 2017, 14, 531–548. [Google Scholar] [CrossRef] [PubMed]
- Heitzer, E.; Ulz, P.; Geigl, J.B. Circulating Tumor DNA as a Liquid Biopsy for Cancer. Clin. Chem. 2015, 61, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Sparano, J.A.; Gray, R.J.; Makower, D.F.; Pritchard, K.I.; Albain, K.S.; Hayes, D.F.; Geyer, C.E.; Dees, E.C.; Goetz, M.P.; Olson, J.A.; et al. Adjuvant Chemotherapy Guided by a 21-Gene Expression Assay in Breast Cancer. N. Engl. J. Med. 2018, 379, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Paik, S.; Shak, S.; Tang, G.; Kim, C.; Baker, J.; Cronin, M.; Baehner, F.L.; Walker, M.G.; Watson, D.; Park, T.; et al. A Multigene Assay to Predict Recurrence of Tamoxifen-Treated, Node-Negative Breast Cancer. N. Engl. J. Med. 2004, 351, 2817–2826. [Google Scholar] [CrossRef] [PubMed]
- Blok, E.J.; van de Velde, C.J.; Smit, V.T. 70-Gene Signature in Early-Stage Breast Cancer. N. Engl. J. Med. 2016, 375, 2199. [Google Scholar] [CrossRef] [PubMed]
- Wirapati, P.; Sotiriou, C.; Kunkel, S.; Farmer, P.; Pradervand, S.; Haibe-Kains, B.; Desmedt, C.; Ignatiadis, M.; Sengstag, T.; Schütz, F.; et al. Meta-Analysis of Gene Expression Profiles in Breast Cancer: Toward a Unified Understanding of Breast Cancer Subtyping and Prognosis Signatures. Breast Cancer Res. 2008, 10, R65. [Google Scholar] [CrossRef]
- Denkert, C.; Loibl, S.; Noske, A.; Roller, M.; Müller, B.M.; Komor, M.; Budczies, J.; Darb-Esfahani, S.; Kronenwett, R.; Hanusch, C.; et al. Tumor-Associated Lymphocytes as an Independent Predictor of Response to Neoadjuvant Chemotherapy in Breast Cancer. J. Clin. Oncol. 2010, 28, 105–113. [Google Scholar] [CrossRef]
- Schalper, K.A.; Velcheti, V.; Carvajal, D.; Wimberly, H.; Brown, J.; Pusztai, L.; Rimm, D.L. In Situ Tumor PD-L1 mRNA Expression Is Associated with Increased TILs and Better Outcome in Breast Carcinomas. Clin Cancer Res. 2014, 20, 2773–2782. [Google Scholar] [CrossRef]
- Loi, S.; Sirtaine, N.; Piette, F.; Salgado, R.; Viale, G.; Van Eenoo, F.; Rouas, G.; Francis, P.; Crown, J.P.A.; Hitre, E.; et al. Prognostic and Predictive Value of Tumor-Infiltrating Lymphocytes in a Phase III Randomized Adjuvant Breast Cancer Trial in Node-Positive Breast Cancer Comparing the Addition of Docetaxel to Doxorubicin with Doxorubicin-Based Chemotherapy: BIG 02-98. J. Clin. Oncol. 2013, 31, 860–867. [Google Scholar] [CrossRef]
- Schmid, P.; Adams, S.; Rugo, H.S.; Schneeweiss, A.; Barrios, C.H.; Iwata, H.; Diéras, V.; Hegg, R.; Im, S.-A.; Shaw Wright, G.; et al. Atezolizumab and Nab-Paclitaxel in Advanced Triple-Negative Breast Cancer. N. Engl. J. Med. 2018, 379, 2108–2121. [Google Scholar] [CrossRef]
- Acheampong, D.O. Bispecific Antibody (bsAb) Construct Formats and Their Application in Cancer Therapy. Protein Pept. Lett. 2019, 26, 479–493. [Google Scholar] [CrossRef]
- Farhangnia, P.; Ghomi, S.M.; Akbarpour, M.; Delbandi, A.-A. Bispecific Antibodies Targeting CTLA-4: Game-Changer Troopers in Cancer Immunotherapy. Front. Immunol. 2023, 14, 1155778. [Google Scholar] [CrossRef]
- Lan, H.-R.; Chen, M.; Yao, S.-Y.; Chen, J.-X.; Jin, K.-T. Bispecific Antibodies Revolutionizing Breast Cancer Treatment: A Comprehensive Overview. Front. Immunol. 2023, 14, 1266450. [Google Scholar] [CrossRef]
- Klein, C.; Brinkmann, U.; Reichert, J.M.; Kontermann, R.E. The Present and Future of Bispecific Antibodies for Cancer Therapy. Nat. Rev. Drug Discov. 2024, 23, 301–319. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Ruwishan, A.; Amer, B.; Salem, A.; Abdi, A.; Chimpandu, N.; Esa, A.; Melemenis, A.; Saleem, M.Z.; Mathew, R.; Gamallat, Y. Advancements in Understanding the Hide-and-Seek Strategy of Hibernating Breast Cancer Cells and Their Implications in Oncology from a Broader Perspective: A Comprehensive Overview. Curr. Issues Mol. Biol. 2024, 46, 8340-8367. https://doi.org/10.3390/cimb46080492
Al-Ruwishan A, Amer B, Salem A, Abdi A, Chimpandu N, Esa A, Melemenis A, Saleem MZ, Mathew R, Gamallat Y. Advancements in Understanding the Hide-and-Seek Strategy of Hibernating Breast Cancer Cells and Their Implications in Oncology from a Broader Perspective: A Comprehensive Overview. Current Issues in Molecular Biology. 2024; 46(8):8340-8367. https://doi.org/10.3390/cimb46080492
Chicago/Turabian StyleAl-Ruwishan, Aiman, Bushra Amer, Ahmed Salem, Ahmed Abdi, Namoonga Chimpandu, Abdelmonem Esa, Alexandros Melemenis, Muhammad Zubair Saleem, Roselit Mathew, and Yaser Gamallat. 2024. "Advancements in Understanding the Hide-and-Seek Strategy of Hibernating Breast Cancer Cells and Their Implications in Oncology from a Broader Perspective: A Comprehensive Overview" Current Issues in Molecular Biology 46, no. 8: 8340-8367. https://doi.org/10.3390/cimb46080492