
Antimicrobial Use and Resistance in Plant Agriculture: A One Health Perspective




Abstract
1. The Problem of Antimicrobial Resistance (AMR)
- Resistance in plant pathogens to pesticides upon repeated exposure, which may be avoided by utilizing different management approaches.
- Resistance in human and animal microorganisms evolved through pesticide use.
2. Mechanisms of Resistance
3. Use of Pesticides
3.1. Antimicrobials in the Environment
3.2. Antibiotic-Resistant Organisms in the Plant Production Environment
4. Resistance to Pesticides Associated with Antimicrobials
4.1. Antibiotics
4.2. Aminoglycosides
4.2.1. Streptomycin
4.2.2. Gentamicin
4.2.3. Kasugamycin
4.3. Tetracyclines
4.4. Quinolones
4.5. The Misuse of Antibiotics
4.6. Antimicrobial Use Selecting for Resistance

{kind=link}
{kind=link}
{kind=link}
| First Author | Key Findings | Reference |
|---|---|---|
| Tolba |
| [110] |
| Heuer |
| [36] |
| van Overbeek |
| [111] |
| Manulis |
| [105] |
| Popowska |
| [112] |
| Yashiro |
| [113] |
| Duffy |
| [114] |
| Shade |
| [115] |
| Walsh |
| [116] |
| Shade |
| [117] |
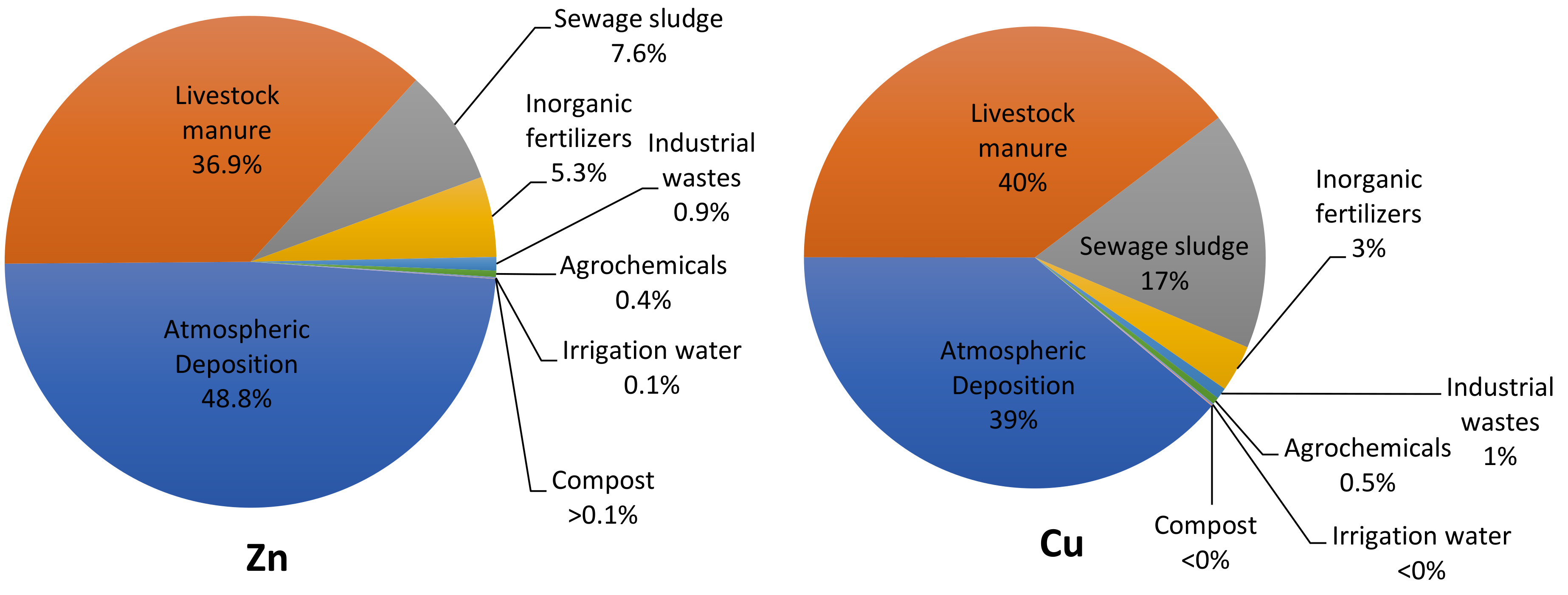
4.7. Metals
4.8. Copper, Zinc, and Arsenic
4.9. Other Fungicides
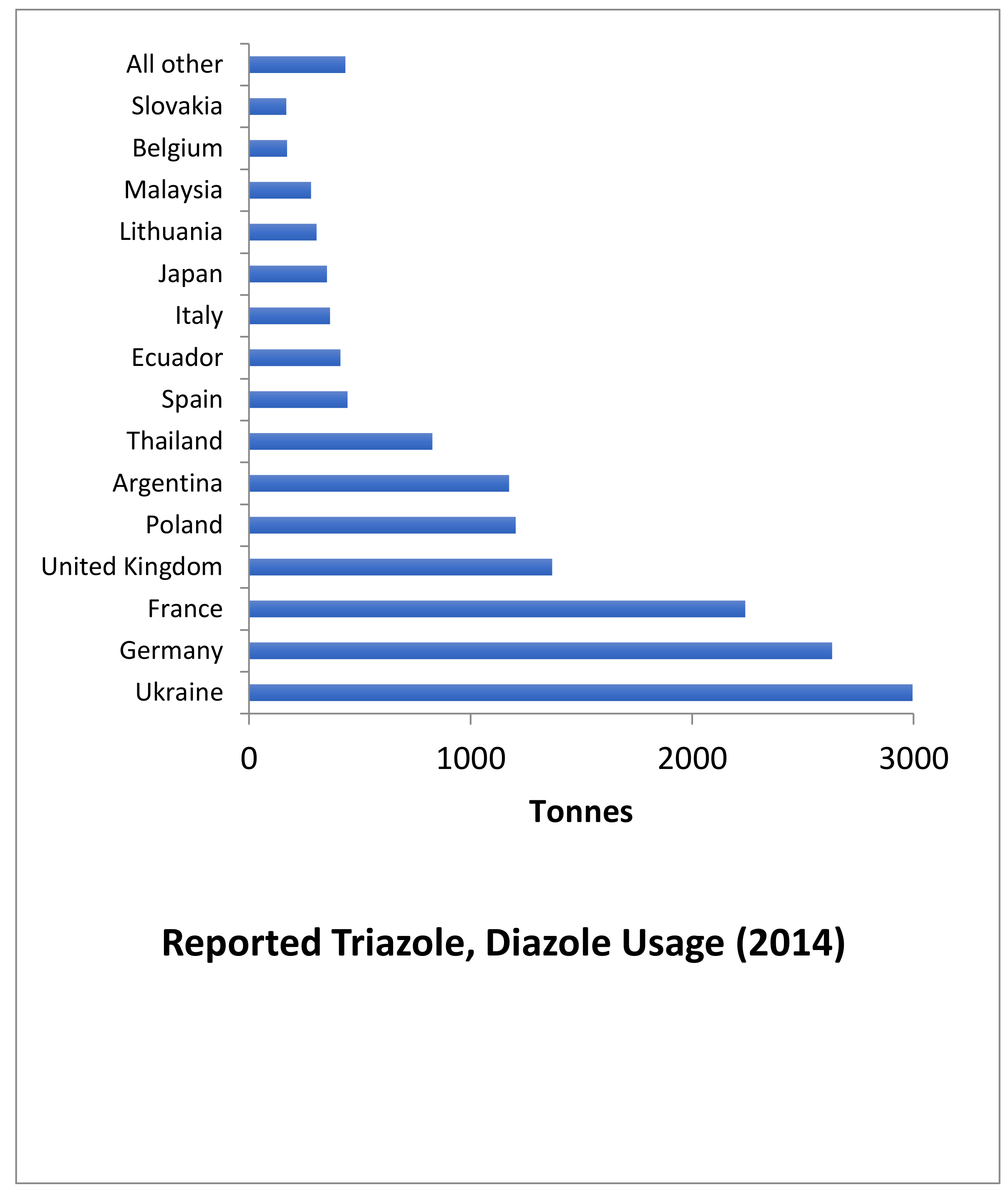
4.9.1. Azoles
4.9.2. Dithiocarbamates
4.9.3. Others
4.10. Fungicide Resistance Action Committee (FRAC)
4.11. Herbicides
5. Data Needs, Recommendations and Conclusions
5.1. Data Needs—Accessible, Reliable Estimates of Pesticides Associated with Antimicrobial Use (AMU) Worldwide
5.2. Data Needs—Surveillance of AMR Organisms in Horticulture and the Environment
5.3. Approaches to Support Judicious Pesticide Use
5.4. Sustainable Production, and Its Contribution in Preventing AMR Development
5.5. Improved Regulation on Pesticides Associated with Antimicrobials
5.6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Glossary
| Antibiotic | An antimicrobial with a spectrum of activity limited to bacteria. |
| Antibiotic resistance | The inherited or acquired characteristic of microorganisms to survive or proliferate in concentrations of an antibiotic that would otherwise kill or inhibit them. |
| Antimicrobial | Any substance of natural, semi-synthetic, or synthetic origin that at in vivo concentrations kills or inhibits the growth of viruses and species of the kingdoms Archaebacteria, Chromista, Eubacteria, Protista, and Fungi. For the purpose of this publication, we consider antimicrobials to include antibiotics (bactericides) and fungicides, and not nematicides, herbicides, plant growth regulators, insecticides, botanicals, or biological control agents. |
| Antimicrobial resistance | The inherited or acquired characteristic of microorganisms to survive or proliferate in concentrations of an antimicrobial that would otherwise kill or inhibit them. |
| Cross-resistance | The ability of a microorganism to multiply or persist in the presence of other members of a particular class of antimicrobial agent or across different classes due to shared mechanisms of resistance. |
| Co-resistance | The ability of a microorganism to multiply or persist in the presence of different classes of antimicrobial agents due to possession of various resistance mechanisms. |
| Pesticide | A substance that “prevents, destroys, or controls a harmful organism (‘pest’) or disease, or protects plants or plant products during production, storage and transport”. The term includes, amongst others: herbicides, fungicides, insecticides, acaricides, nematicides, molluscicides, rodenticides, growth regulators, repellents, and biocides. |
References
- Pimentel, D.; Burgess, M. Environmental and economic costs of the application of pesticides primarily in the United States. In Integrated Pest Management; Pimentel, R., Peshin, R., Eds.; Springer: New York, NY, USA, 2014. [Google Scholar]
- Levy, S.B. Factors impacting on the problem of antibiotic resistance. J. Antimicrob. Chemother. 2002, 49, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Abe, M.; Kimoto, M.; Furukawa, S.; Nakazawa, T. The DsbA-DsbB disulfide bond formation system of Burkholderia cepacia is involved in the production of protease and alkaline phosphatase, motility, metal resistance, and multi-drug resistance. Microbiol. Immunol. 2000, 44, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Gunn, L.; Wall, P.; Fanning, S. Antimicrobial resistance and its association with tolerance to heavy metals in agriculture production. Food Microbiol. 2017, 64, 23–32. [Google Scholar] [CrossRef]
- Delp, C.J. Coping with resistance to plant disease control agents. Plant Dis. 1980, 64, 652–657. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Available online: https://apo.org.au/sites/default/files/resource-files/2016-05/apo-nid63983.pdf (accessed on 19 January 2022).
- FAO. Drivers, Dynamics and Epidemiology of Antimicrobial Resistance in Animal Production; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- WHO. Global Action Plan on Antimicrobial Resistance. 2016. Available online: https://www.who.int/publications/i/item/9789241509763/ (accessed on 19 January 2022).
- Greene, S.; Reid, A. Moving targets: Fighting resistance in infections, cancers, pest. Microbe 2013, 8, 279–285. [Google Scholar] [CrossRef][Green Version]
- Nelson, M.L.; Dinardo, A.; Hochberg, J.; Armelagos, G.J. Brief communication: Mass spectroscopic characterization of tetracycline in the skeletal remains of an ancient population from Sudanese Nubia 350-550 CE. Am. J. Phys. Anthropol. 2010, 143, 151–154. [Google Scholar] [CrossRef]
- Fleming, A. On the antibacterial action of cultures of a Penicillium, with special reference to their use in the isolation of B. influenzae. Br. J. Exp. Pathol. 1929, 10, 226–236. [Google Scholar] [CrossRef]
- Bud, R. Antibiotics: The epitome of a wonder drug. BMJ 2007, 334, s6. [Google Scholar] [CrossRef]
- Fleming, A. Penicillin. Sir Alexander Fleming—Nobel Lecture. NobelPrize.org. 1945. Available online: https://www.nobelprize.org/prizes/medicine/1945/fleming/lecture/ (accessed on 19 January 2022).
- Cloeckaert, A.; Zygmunt, M.; Doublet, B. Editorial: Genetics of acquired antimicrobial resistance in animal and zoonotic pathogens. Front. Microbiol. 2017, 8, 2428. [Google Scholar] [CrossRef]
- MacLean, R.C.; Millan, A.S. The evolution of antibiotic resistance. Science 2019, 365, 1082–1083. [Google Scholar] [CrossRef]
- Walsh, C. Antibiotics: Actions, Origins, Resistance; American Society for Microbiology (ASM): Washington, DC, USA, 2003. [Google Scholar]
- European Commission. Pesticides. 2017. Available online: https://ec.europa.eu/food/plant/pesticides_en (accessed on 17 July 2017).
- Euopean Commission. EU Pesticide Database. 2017. Available online: https://ec.europa.eu/food/plants/pesticides/eu-pesticides-database_en (accessed on 19 January 2022).
- FAO. New Standards to Curb the Global Spread of Plant Pests and Diseases. 2019. Available online: http://www.fao.org/news/story/en/item/1187738/icode/ (accessed on 19 January 2022).
- Researchandmarkets.com. Global Crop Protection Market—By Type, Crop Type, Regions and Vendors—Market Size, Demand Forecasts, Industry Trends and Updates (2016–2022). 2017. Available online: http://www.researchandmarkets.com/reports/4176102/global-crop-protection-market-by-type-crop#pos-9 (accessed on 19 January 2022).
- Strange, R.N.; Scott, P.R. Plant Disease: A Threat to Global Food Security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- Latgé, J.-P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef] [PubMed]
- Shenge, K.; LeJeune, J. One Health: A focus on interdisciplinary collaboration. Adv. Plants Agric. Res. 2014, 1, 00018. [Google Scholar] [CrossRef][Green Version]
- Tyrrell, C.; Burgess, C.M.; Brennan, F.P.; Walsh, F. Antibiotic resistance in grass and soil. Biochem. Soc. Trans. 2019, 47, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Baltz, R.H. Renaissance in antibacterial discovery from actinomycetes. Curr. Opin. Pharmacol. 2008, 8, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Steen, A.D.; Crits-Christoph, A.; Carini, P.; DeAngelis, K.M.; Fierer, N.; Lloyd, K.G.; Thrash, J.C. High proportions of bacteria and archaea across most biomes remain uncultured. ISME J. 2019, 13, 3126–3130. [Google Scholar] [CrossRef]
- Katz, M.; Hover, B.M.; Brady, S.F. Culture-independent discovery of natural products from soil metagenomes. J. Ind. Microbiol. Biotechnol. 2016, 43, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Larsson, D.G. Antibiotics in the environment. Ups. J. Med. Sci. 2014, 119, 108–112. [Google Scholar] [CrossRef]
- Kim, K.-R.; Owens, G.; Kwon, S.-I.; So, K.-H.; Lee, D.-B.; Ok, Y.S. Occurrence and Environmental Fate of Veterinary Antibiotics in the Terrestrial Environment. Water Air Soil Pollut. 2011, 214, 163–174. [Google Scholar] [CrossRef]
- Dolliver, H.; Gupta, S. Antibiotic Losses in Leaching and Surface Runoff from Manure-Amended Agricultural Land. J. Environ. Qual. 2008, 37, 1227–1237. [Google Scholar] [CrossRef]
- Holvoet, K.; Sampers, I.; Callens, B.; Dewulf, J.; Uyttendaele, M. Moderate Prevalence of Antimicrobial Resistance in Escherichia coli Isolates from Lettuce, Irrigation Water, and Soil. Appl. Environ. Microbiol. 2013, 79, 6677–6683. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antimicrobial activity of essential oils and other plant extracts. J. Appl. Microbiol. 1999, 86, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Elbehri, A. Biopharming and the food system: Examining the potential benefits and risks. AgBioForum 2005, 8, 18–25. [Google Scholar]
- MoRST. Biotechnologies to 2025. Available online: https://www.mcguinnessinstitute.org/wp-content/uploads/2021/04/MoRST-Biotechnology-Roadmap.pdf (accessed on 19 January 2022).
- WHO. Critically Important Antimicrobials for Human Medicine—5th rev. 2017. Available online: https://apps.who.int/iris/bitstream/handle/10665/255027/9789241512220-eng.pdf (accessed on 19 January 2022).
- Heuer, H.; Krögerrecklenfort, E.; Wellington, E.M.H.; Egan, S.; Van Elsas, J.D.; Van Overbeek, L.; Collard, J.-M.; Guillaume, G.; Karagouni, A.D.; Nikolakopoulou, T.L.; et al. Gentamicin resistance genes in environmental bacteria: Prevalence and transfer. FEMS Microbiol. Ecol. 2002, 42, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Mtui, H. Effect of mulch and different pre-harvest fungicide spray regimes on shelf life of tomato (Solanum lycopersicum L.) in Tanzania. Asian J. Plant Sci. Res. 2014, 4, 9–13. [Google Scholar]
- Bhat, K.A.; Bhat, N.A.; Mohiddin, F.A.; Mir, S.A.; Mir, M.R. Management of post-harvest Pectobacterium soft rot of cabbage (Brassica oleracea var capitata L.) by biocides and packing material. Afr. J. Agric. Res. 2012, 7, 4066–4074. [Google Scholar] [CrossRef]
- Mathers, H.; Lowe, S.; Scagel, C.; Struve, D.; Case, L. Abiotic Factors Influencing Root Growth of Woody Nursery Plants in Containers. HortTechnology 2007, 17, 151–162. [Google Scholar] [CrossRef]
- Dayan, A. Allergy to antimicrobial residues in food: Assessment of the risk to man. Vet. Microbiol. 1993, 35, 213–226. [Google Scholar] [CrossRef]
- Tinkelman, D.G.; Bock, S.A. Anaphylaxis presumed to be caused by beef containing streptomycin. Ann. Allergy 1984, 53, 243–244. [Google Scholar]
- Brasse, D. Stellunghname der BBA zum Streptomycin-Problem. ADIZ 2001, 6, 24–25. [Google Scholar]
- Graham, F.; Paradis, L.; Bégin, P.; Paradis, J.; Babin, Y.; Roches, A.D. Risk of allergic reaction and sensitization to antibiotics in foods. Ann. Allergy Asthma Immunol. 2014, 113, 329–330. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Gupta, S.C.; Baidoo, S.K.; Chander, Y.; Rosen, C.J. Antibiotic Uptake by Plants from Soil Fertilized with Animal Manure. J. Environ. Qual. 2005, 34, 2082–2085. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chan, K.K.J.; Chan, W. Plant Uptake and Metabolism of Nitrofuran Antibiotics in Spring Onion Grown in Nitrofuran-Contaminated Soil. J. Agric. Food Chem. 2017, 65, 4255–4261. [Google Scholar] [CrossRef] [PubMed]
- Boxall, A.B.A.; Johnson, P.; Smith, E.J.; Sinclair, C.J.; Stutt, A.E.; Levy, L.S. Uptake of Veterinary Medicines from Soils into Plants. J. Agric. Food Chem. 2006, 54, 2288–2297. [Google Scholar] [CrossRef]
- Tasho, R.P.; Cho, J.Y. Veterinary antibiotics in animal waste, its distribution in soil and uptake by plants: A review. Sci. Total Environ. 2016, 563–564, 366–376. [Google Scholar] [CrossRef]
- Huang, R.; Ding, P.; Huang, D.; Yang, F. Antibiotic pollution threatens public health in China. Lancet 2015, 385, 773–774. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Chaudhury, R.R. Antibiotic Resistance in India: Drivers and Opportunities for Action. PLoS Med. 2016, 13, e1001974. [Google Scholar] [CrossRef]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilbäck, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of Resistant Bacteria at Very Low Antibiotic Concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The Shared Antibiotic Resistome of Soil Bacteria and Human Pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef]
- Goldstein, D.; Tinland, B.; Gilbertson, L.; Staub, J.; Bannon, G.; Goodman, R.; McCoy, R.; Silvanovich, A. Human safety and genetically modified plants: A review of antibiotic resistance markers and future transformation selection technologies. J. Appl. Microbiol. 2005, 99, 7–23. [Google Scholar] [CrossRef]
- EFSA. Opinion of the Scientific Panel on Genetically Modified Organisms on the use of antibiotic resistance genes as marker genes in genetically modified plants1 (Question N° EFSA-Q-2003-109). EFSA J. 2004, 48, 1–18. [Google Scholar]
- EFSA. EFSA-Q-2009-00589 and EFSA-Q-2009-00593. Consolidated presentation of the joint Scientific Opinion of the GMO and BIOHAZ Panels on the “Use of Antibiotic Resistance Genes as Marker Genes in Genetically Modified Plants” and the Scientific Opinion of the GMO Panel on “Consequences of the Opinion on the Use of Antibiotic Resistance Genes as Marker Genes in Genetically Modified Plants on Previous EFSA Assessments of Individual GM Plants”. EFSA 2009. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/1108 (accessed on 19 January 2022).
- Beuchat, L.R. Vectors and conditions for preharvest contamination of fruits and vegetables with pathogens capable of causing enteric diseases. Br. Food J. 2006, 108, 38–53. [Google Scholar] [CrossRef]
- Kang, Y.; Shen, M.; Xia, D.; Ye, K.; Zhao, Q.; Hu, J. Caution of intensified spread of antibiotic resistance genes by inadvertent introduction of beneficial bacteria into soil. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2017, 108, 1–7. [Google Scholar] [CrossRef]
- Hölzel, C.S.; Tetens, J.L.; Schwaiger, K. Unraveling the Role of Vegetables in Spreading Antimicrobial-Resistant Bacteria: A Need for Quantitative Risk Assessment. Foodborne Pathog. Dis. 2018, 15, 671–688. [Google Scholar] [CrossRef]
- Guinée, P.; Ugueto, N.; van Leeuwen, N. Escherichia coli with resistance factors in vegetarians, babies, and nonvegetarians. Appl. Microbiol. 1970, 20, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Sannes, M.R.; Belongia, E.A.; Kieke, B.; Smith, K.; Kieke, A.; Vandermause, M.; Bender, J.; Clabots, C.; Winokur, P.; Johnson, J.R. Predictors of antimicrobial-resistant Escherichia coli in the feces of vegetarians and newly hospitalized adults in Minnesota and Wisconsin. J. Infect. Dis. 2008, 197, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Braak, N.V.D.; Van Belkum, A.; Kreft, D.; Verbrugh, H.; Endtz, H. Dietary habits and gastrointestinal colonization by antibiotic resistant microorganisms. J. Antimicrob. Chemother. 2001, 47, 498–500. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Losasso, C.; Di Cesare, A.; Mastrorilli, E.; Patuzzi, I.; Cibin, V.; Eckert, E.M.; Fontaneto, D.; Vanzo, A.; Ricci, A.; Corno, G. Assessing antimicrobial resistance gene load in vegan, vegetarian and omnivore human gut microbiota. Int. J. Antimicrob. Agents 2018, 52, 702–705. [Google Scholar] [CrossRef] [PubMed]
- Furuya-Kanamori, L.; Stone, J.; Yakob, L.; Kirk, M.; Collignon, P.; Mills, D.J.; Lau, C.L. Risk factors for acquisition of multidrug-resistant Enterobacterales among international travellers: A synthesis of cumulative evidence. J. Travel Med. 2019, 271-10. [Google Scholar] [CrossRef] [PubMed]
- Scherer, A.; Vogt, H.R.; Vilei, E.M.; Frey, J.; Perreten, V. Enhanced antibiotic multi-resistance in nasal and faecal bacteria after agricultural use of streptomycin. Environ. Microbiol. 2013, 15, 297. [Google Scholar] [CrossRef]
- CDC. Multistate Outbreak of Salmonella Poona Infections Linked to Imported Cucumbers (Final Update). Available online: https://www.cdc.gov/salmonella/poona-09-15/index.html (accessed on 19 January 2022).
- CDC. Multistate Outbreak of Salmonella Urbana Infections Linked to Imported Maradol Papayas (Final Update). Available online: https://www.cdc.gov/salmonella/urbana-09-17/index.html (accessed on 19 January 2022).
- Hassan, R.; Whitney, B.; Williams, D.L.; Holloman, K.; Grady, D.; Thomas, D.; Omoregie, E.; Lamba, K.; Leeper, M.; Gieraltowski, L.; et al. Multistate outbreaks of Salmonella infections linked to imported Maradol papayas–United States, December 2016–September 2017. Epidemiol. Infect. 2019, 147, e265. [Google Scholar] [CrossRef] [PubMed]
- USEPA. What Are Antimicrobial Pesticides? 2017. Available online: https://www.epa.gov/pesticide-registration/what-are-antimicrobial-pesticides (accessed on 19 January 2022).
- OIE. OIE List of Antimicrobial Agents of Veterinary Importance. 2019. Available online: https://www.oie.int/fileadmin/Home/eng/Our_scientific_expertise/docs/pdf/AMR/A_OIE_List_antimicrobials_July2019.pdf (accessed on 19 January 2022).
- Davis, B.D. Mechanism of bactericidal action of aminoglycosides. Microbiol. Rev. 1987, 51, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Registration Certificate of the Last Streptomycin for Agriculture Use Expires in China. 2016. Available online: http://www.cnchemicals.com/Detail/Readonline.aspx?id=5887&type=n&cid=20879621633&site=online (accessed on 18 July 2017).
- Erickson, B.E. Groups challenge EPA approval of streptomycin use on citrus. Chem. Eng. News 2021, 14. [Google Scholar] [CrossRef]
- Davies, J.; Wright, G. Bacterial resistance to aminoglycoside antibiotics. Trends Microbiol. 1997, 5, 234–240. [Google Scholar] [CrossRef]
- Waksman, S.; Reilly, H.C.; Schatz, A. Strain Specificity and Production of Antibiotic Substances. Proc. Natl. Acad. Sci. USA 1945, 31, 157–164. [Google Scholar] [CrossRef]
- van Duijkeren, E.; Schink, A.-K.; Roberts, M.C.; Wang, Y.; Schwarz, S. Mechanisms of bacterial resistance to antimicrobial agents. Microbiol. Spectr. 2018, 6, 2–6. [Google Scholar]
- Springer, B.; Kidan, Y.G.; Prammananan, T.; Ellrott, K.; Bottger, E.C.; Sander, P. Mechanisms of streptomycin resistance: Selection of mutations in the 16S rRNA gene conferring resistance. Antimicrob. Agents Chemother. 2001, 45, 2877–2884. [Google Scholar] [CrossRef]
- Förster, H.; McGhee, G.C.; Sundin, G.W.; Adaskaveg, J.E. Characterization of Streptomycin Resistance in Isolates of Erwinia amylovora in California. Phytopathology 2015, 105, 1302–1310. [Google Scholar] [CrossRef]
- Sundin, G.W.; Bender, C.L. Expression of the strA-strB streptomycin resistance genes in Pseudomonas syringae and Xanthomonas campestris and characterization of IS6100 in X. campestris. Appl. Env. Microbiol. 1995, 61, 2891. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, X.-F.; Zhou, M.-G.; Kuang, J.; Zhang, Y.; Shang, Y.; Wang, J.-X. Status of streptomycin resistance development in Xanthomonas oryzae pv. oryzae and Xanthomonas oryzae pv. oryzicola in China and their resistance characters. J. Phytopathol. 2010, 158, 601. [Google Scholar]
- Schnabel, E.L.; Jones, A.L. Distribution of Tetracycline Resistance Genes and Transposons among Phylloplane Bacteria in Michigan Apple Orchards. Appl. Environ. Microbiol. 1999, 65, 4898–4907. [Google Scholar] [CrossRef] [PubMed]
- McManus, P.S. Does a drop in the bucket make a splash? Assessing the impact of antibiotic use on plants. Curr. Opin. Microbiol. 2014, 19, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.J.; Rather, P.N.; Hare, R.S.; Miller, G.H. Molecular genetics of aminoglycoside resistance genes and familial relationships of the aminoglycoside-modifying enzymes. Microbiol. Rev. 1993, 57, 138–163. [Google Scholar] [CrossRef] [PubMed]
- McManus, P.S.; Jones, A.L. Epidemiology and genetic analysis of streptomycin-resistant Erwinia amylovora from Michigan and evaluation of oxytetracycline for control. Phytopathology 1994, 84, 627. [Google Scholar] [CrossRef]
- Chiou, C.S.; Jones, A.L. The analysis of plasmid-mediated streptomycin resistance in Erwinia amylovora. Phytopathology 1991, 81, 710. [Google Scholar] [CrossRef]
- Serio, A.W.; Magalhães, M.L.; Blanchard, J.S.; Connolly, L.E. Aminoglycosides: Mechanisms of Action and Resistance. In Antimicrobial Drug Resistance: Mechanisms of Drug Resistance; Mayers, D.L., Ed.; Springer International Publishing: Cham, Switzerland, 2017; Volume 1, pp. 213–229. [Google Scholar]
- Sobiczewski, P.; Jones, A.L.; Chiou, C.S. Streptomycin-resistant epiphytic bacteria with homologous DNA for streptomycin resistance in Michigan apple orchards. Plant Dis. 1991, 75, 1110. [Google Scholar] [CrossRef]
- Huang, T.-C.; Burr, T.J. Characterization of plasmids that encode streptomycin-resistance in bacterial epiphytes of apple. J. Appl. Microbiol. 1999, 86, 741–751. [Google Scholar] [CrossRef]
- Sundin, G.W.; Bender, C.L. Dissemination of the strA-strB streptomycin-resistance genes among commensal and pathogenic bacteria from humans, animals, and plants. Mol. Ecol. 1996, 5, 133–143. [Google Scholar] [CrossRef]
- Sundin, G.W.; Bender, C.L. Ecological and genetic analysis of copper and streptomycin resistance in Pseudomonas syringae pv. syringae. Appl. Environ. Microbiol. 1993, 59, 1018–1024. [Google Scholar] [CrossRef]
- Weinstein, M.J.; Luedemann, G.M.; Oden, E.M.; Wagman, G.H.; Rosselet, J.P.; Marquez, J.A.; Coniglio, C.T.; Charney, W.; Herzog, H.L.; Black, J. Gentamicin, a new antibiotic complex from micromonospora. J. Med. Chem. 1963, 6, 463–464. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updates 2010, 13, 151–171. [Google Scholar] [CrossRef] [PubMed]
- WHO. Integrated Surveillance of Antimicrobial Resistance in Foodborne Bacteria: Application of a One Health Approach: Guidance from the WHO Advisory Group on Integrated Surveillanec of Antimicrobial Resistance (AGISAR); World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Umezawa, H.; Hamada, M.; Suhara, Y.; Hashimoto, T.; Ikekawa, T. Kasugamycin, a new antibiotic. Antimicrob. Agents Chemother. 1965, 5, 753–757. [Google Scholar]
- Levitan, A.A. In vitro antibacterial activity of kasugamycin. Appl. Microbiol. 1967, 15, 750–753. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, A.; Moriyama, H.; Fukuhara, T. The novel kasugamycin 2′-N-acetyltransferase gene aac (2′)-IIa, carried by the IncP island, confers kasugamycin resistance to rice-pathogenic bacteria. Appl. Environ. Microbiol. 2012, 78, 5555–5564. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, A.; Kobayashi, Y. Kasugamycin-resistant mutants of Bacillus subtilis. J. Bacteriol. 1978, 135, 1149–1150. [Google Scholar] [CrossRef] [PubMed]
- Fouts, E.K.; Barbour, S.D. Transductional mapping of ksgB and a new Tn5-induced kasugamycin resistance gene, ksgD, in Escherichia coli K-12. J. Bacteriol. 1981, 145, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Okuyama, A.; Tanaka, N. A Third Kasugamycin Resistance Locus, ksgC, Affecting Ribosomal Protein S2 in Escherichia coli K-12. J. Bacteriol. 1975, 122, 796–797. [Google Scholar] [CrossRef] [PubMed]
- Shiver, A.L.; Osadnik, H.; Kritikos, G.; Li, B.; Krogan, N.; Typas, A.; Gross, C.A. A Chemical-Genomic Screen of Neglected Antibiotics Reveals Illicit Transport of Kasugamycin and Blasticidin S. PLoS Genet. 2016, 12, e1006124. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef]
- Stockwell, V.; Duffy, B. Use of antibiotics in plant agriculture. Rev. Sci. Tech. l’OIE 2012, 31, 199–210. [Google Scholar] [CrossRef]
- Hughes, V.M.; Datta, N. Conjugative plasmids in bacteria of the ’pre-antibiotic’ era. Nature 1983, 302, 725–726. [Google Scholar] [CrossRef] [PubMed]
- Sundin, G.W.; Wang, N. Antibiotic Resistance in Plant-Pathogenic Bacteria. Annu. Rev. Phytopathol. 2018, 56, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhou, Q.; Luo, Y. Occurrence and source analysis of typical veterinary antibiotics in manure, soil, vegetables and groundwater from organic vegetable bases, northern China. Environ. Pollut. 2010, 158, 2992–2998. [Google Scholar] [CrossRef]
- Hikichi, Y.; Egami, H.; Oguri, Y.; Okuno, T. Fitness for Survival of Burkholderia glumae Resistant to Oxolinic Acid in Rice Plants. Jpn. J. Phytopathol. 1998, 64, 147–152. [Google Scholar] [CrossRef]
- Manulis, S.; Kleitman, F.; Shtienberg, D.; Shwartz, H.; Oppenheim, D.; Zilberstaine, M.; Shabi, E. Changes in the Sensitivity of Erwinia amylovora Populations to Streptomycin and Oxolinic Acid in Israel. Plant Dis. 2003, 87, 650–654. [Google Scholar] [CrossRef]
- Hikichi, Y.; Tsujiguchi, K.; Maeda, Y.; Okuno, T. Development of Increased Oxolinic Acid-resistance in Burkholderia glumae. J. Gen. Plant Pathol. 2001, 67, 58–62. [Google Scholar] [CrossRef]
- Shin, K.; Ascunce, M.S.; Narouei-Khandan, H.A.; Sun, X.; Jones, D.; Kolawole, O.O.; Goss, E.M.; van Bruggen, A.H.C. Effects and side effects of penicillin injection in huanglongbing affected grapefruit trees. Crop Prot. 2016, 90, 106–116. [Google Scholar] [CrossRef]
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The threat of antimicrobial resistance in developing countries: Causes and control strategies. Antimicrob. Resist. Infect. Control 2017, 6, 1–8. [Google Scholar] [CrossRef]
- Volkova, V.V.; KuKanich, B.; Riviere, J.E. Exploring Post-Treatment Reversion of Antimicrobial Resistance in Enteric Bacteria of Food Animals as a Resistance Mitigation Strategy. Foodborne Pathog. Dis. 2016, 13, 610–617. [Google Scholar] [CrossRef]
- Tolba, S.; Egan, S.; Kallifidas, D.; Wellington, E.M.H. Distribution of streptomycin resistance and biosynthesis genes in streptomycetes recovered from different soil sites. FEMS Microbiol. Ecol. 2002, 42, 269–276. [Google Scholar] [CrossRef][Green Version]
- Overbeek, L.S.; Wellington, E.M.; Egan, S.; Smalla, K.; Heuer, H.; Collard, J.-M.; Guillaume, G.; Karagouni, A.D.; Nikolakopoulou, T.L.; Elsas, J.D. Prevalence of streptomycin-resistance genes in bacterial populations in European habitats. FEMS Microbiol. Ecol. 2002, 42, 277–288. [Google Scholar] [CrossRef]
- Popowska, M.; Rzeczycka, M.; Miernik, A.; Krawczyk-Balska, A.; Walsh, F.; Duffy, B. Influence of Soil Use on Prevalence of Tetracycline, Streptomycin, and Erythromycin Resistance and Associated Resistance Genes. Antimicrob. Agents Chemother. 2011, 56, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Yashiro, E.; McManus, P.S. Effect of Streptomycin Treatment on Bacterial Community Structure in the Apple Phyllosphere. PLoS ONE 2012, 7, e37131. [Google Scholar] [CrossRef] [PubMed]
- Duffy, B.; Holliger, E.; Walsh, F. Streptomycin use in apple orchards did not increase abundance of mobile resistance genes. FEMS Microbiol. Lett. 2014, 350, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Shade, A.; McManus, P.S.; Handelsman, J. Unexpected Diversity during Community Succession in the Apple Flower Microbiome. mBio 2013, 4, e00602-12. [Google Scholar] [CrossRef]
- Walsh, F.; Smith, D.P.; Owens, S.M.; Duffy, B.; Frey, J.E. Restricted streptomycin use in apple orchards did not adversely alter the soil bacteria communities. Front. Microbiol. 2014, 4, 383. [Google Scholar] [CrossRef]
- Shade, A.; Klimowicz, A.K.; Spear, R.N.; Linske, M.; Donato, J.J.; Hogan, C.S.; McManus, P.S.; Handelsman, J. Streptomycin Application Has No Detectable Effect on Bacterial Community Structure in Apple Orchard Soil. Appl. Environ. Microbiol. 2013, 79, 6617–6625. [Google Scholar] [CrossRef]
- Lobanok, T.E.; Pesniakevich, A.G.; Fomichev, I.K. Characteristics of the conjugation transfer of the R plasmids in bacteria of the intestinal group to Erwinia cells. Γehemuka 1978, 14, 2119. [Google Scholar]
- Llama-Palacios, A.; Loópez-Solanilla, E.; Rodriíguez-Palenzuela, P. The ybiT Gene of Erwinia chrysanthemi Codes for a Putative ABC Transporter and Is Involved in Competitiveness against Endophytic Bacteria during Infection. Appl. Environ. Microbiol. 2002, 68, 1624–1630. [Google Scholar] [CrossRef]
- Kleitman, F.; Shtienberg, D.; Blachinsky, D.; Oppenheim, D.; Zilberstaine, M.; Dror, O.; Manulis, S. Erwinia amylovora populations resistant to oxolinic acid in Israel: Prevalence, persistence and fitness. Plant Pathol. 2005, 54, 108–115. [Google Scholar] [CrossRef]
- Nicholson, F.A.; Smith, S.R.; Alloway, B.J.; Carlton-Smith, C.; Chambers, B.J. An inventory of heavy metals inputs to agricultural soils in England and Wales. Sci. Total Environ. 2003, 311, 205–219. [Google Scholar] [CrossRef]
- AROMIS. Assessment and Reduction of Heavy Metal Input into Agro-Ecosystems. Available online: https://www.aramis.admin.ch/Texte/?ProjectID=12813&Sprache=fr-CH (accessed on 18 July 2017).
- Punshon, T.; Jackson, B.P.; Meharg, A.A.; Warczack, T.; Scheckel, K.; Guerinot, M.L. Understanding arsenic dynamics in agronomic systems to predict and prevent uptake by crop plants. Sci. Total Environ. 2017, 581–582, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Bencko, V.; Foong, F.Y.L. The history of arsenical pesticides and health risks related to the use of Agent Blue. Ann. Agric. Environ. Med. 2017, 24, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Brammer, H.; Ravenscroft, P. Arsenic in groundwater: A threat to sustainable agriculture in South and South-east Asia. Environ. Int. 2009, 35, 647–654. [Google Scholar] [CrossRef]
- Rutherford, D.W.; Bednar, A.J.; Garbarino, J.R.; Needham, R.; Staver, K.W.; Wershaw, R.L. Environmental Fate of Roxarsone in Poultry Litter. Part II. Mobility of Arsenic in Soils Amended with Poultry Litter. Environ. Sci. Technol. 2003, 37, 1515–1520. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G.J. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acids Res. 2014, 42, D737–D743. [Google Scholar] [CrossRef]
- Voloudakis, A.E.; Bender, C.L.; Cooksey, D.A. Similarity between Copper Resistance Genes from Xanthomonas campestris and Pseudomonas syringae. Appl. Environ. Microbiol. 1993, 59, 1627–1634. [Google Scholar] [CrossRef]
- Araújo, E.; Pereira, R.; Ferreira, M.; Quezado-Duval, A.; Café-Filho, A. Sensitivity of Xanthomonads Causing Tomato Bacterial Spot to Copper and Streptomycin and in vivo Infra-Specific Competitive Ability in Xanthomonas perforans Resistant and Sensitive to Copper. J. Plant Pathol. 2012, 94, 79–87. [Google Scholar]
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Co-occurrence of resistance genes to antibiotics, biocides and metals reveals novel insights into their co-selection potential. BMC Genom. 2015, 16, 1–14. [Google Scholar] [CrossRef]
- Hasman, H.; Kempf, I.; Chidaine, B.; Cariolet, R.; Ersbøll, A.K.; Houe, H.; Bruun Hansen, H.C.; Aarestrup, F.M. Copper Resistance in Enterococcus faecium, Mediated by the tcrB Gene, Is Selected by Supplementation of Pig Feed with Copper Sulfate. Appl. Environ. Microbiol. 2006, 72, 5784–5789. [Google Scholar] [CrossRef] [PubMed]
- Mourão, J.; Novais, C.; Machado, J.; Peixe, L.; Antunes, P. Metal tolerance in emerging clinically relevant multidrug-resistant Salmonella enterica serotype 4,5,12:i:− clones circulating in Europe. Int. J. Antimicrob. Agents 2015, 45, 610–616. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Food and Agriculture Organization of the United Nations. 2020. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 11 June 2021).
- Jiang, A.-L.; Hu, W.-Z.; Tian, M.; Fan, S.-D. Study on utilization of natamycin in storage of strawberry fruit. Food Sci. 2007, 28, 515–520. [Google Scholar]
- Boothe, D. Antifungal Agents. 2017. Available online: http://www.merckvetmanual.com/pharmacology/antifungal-agents (accessed on 19 January 2022).
- Ahmed, M.Z.; Rao, T.; Saeed, A.; Mutahir, Z.; Hameed, S.; Inayat, S.; Shahzad, H.; Ullah, N.; Abaid-Ullah, M.; Ibrahim, M.; et al. Antifungal Drugs: Mechanism of Action and Resistance. In Biochemistry of Drug Resistance; Ahmed, S., Chandra Ojha, S., Najam-ul-Haq, M., Younus, M., Hashmi, M.Z., Eds.; Springer: Cham, Switzerland, 2021; pp. 143–165. [Google Scholar]
- Palanti, S.; Susco, D. A new wood preservative based on heated oil treatment combined with triazole fungicides developed for above-ground conditions. Int. Biodeterior. Biodegrad. 2004, 54, 337–342. [Google Scholar] [CrossRef]
- Hof, H. Critical Annotations to the Use of Azole Antifungals for Plant Protection. Antimicrob. Agents Chemother. 2001, 45, 2987–2990. [Google Scholar] [CrossRef]
- Sharma, C.; Nelson-Sathi, S.; Singh, A.; Pillai, M.R.; Chowdhary, A. Genomic perspective of triazole resistance in clinical and environmental Aspergillus fumigatus isolates without cyp51A mutations. Fungal Genet. Biol. 2019, 132, 103265. [Google Scholar] [CrossRef]
- Hamey, P.Y.; Harris, C.A. The variation of pesticide residues in fruits and vegetables and the associated assessment of risk. Regul. Toxicol. Pharmacol. 1999, 30, S34–S41. [Google Scholar] [CrossRef]
- Ribes e Ribes, A.D.; Spolti, P.; Del Ponte, E.M.; Donato, K.Z.; Schrekker, H.; Fuentefria, A.M. Is the emergence of fungal resistance to medical triazoles related to their use in the agroecosystems? A mini review. Braz. J. Microbiol. 2016, 47, 793–799. [Google Scholar] [CrossRef]
- Snelders, E.; Van Der Lee, H.A.L.; Kuijpers, J.; Rijs, A.J.M.M.; Varga, J.; Samson, R.A.; Mellado, E.; Donders, A.R.T.; Melchers, W.; Verweij, P.E. Emergence of Azole Resistance in Aspergillus fumigatus and Spread of a Single Resistance Mechanism. PLoS Med. 2008, 5, e219. [Google Scholar] [CrossRef]
- Snelders, E.; Camps, S.M.T.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.; Van Der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.G.; Verweij, P.E. Triazole Fungicides Can Induce Cross-Resistance to Medical Triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef]
- Vermeulen, E.; Largou, K.; Verweij, P.E. Azole resistance in Aspergillus fumigatus: A growing public health concern. Curr. Opin. Infect. Dis. 2013, 26, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Thorrold, C.; Letsoalo, M.; Dusé, A.; Marais, E. Efflux pump activity in fluoroquinolone and tetracycline resistant Salmonella and E. coli implicated in reduced susceptibility to household antimicrobial cleaning agents. Int. J. Food Microbiol. 2007, 113, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Wang, C.; Chung, Y.-R.; Kim, J.-Y.; Choi, E.-S.; Kim, S.-W. Enhancement of geraniol resistance of Escherichia coli by MarA overexpression. J. Biosci. Bioeng. 2013, 115, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.-M.; Amaral, L.; Bolla, J.-M. Geraniol Restores Antibiotic Activities against Multidrug-Resistant Isolates from Gram-Negative Species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef]
- Morton, V.; Staub, T. A Short History of Fungicides. Available online: http://davidmoore.org.uk/21st_Century_Guidebook_to_Fungi_PLATINUM/Assets/Printed_documents/MortonStaub_History_of_Fungicides.pdf (accessed on 19 January 2022).
- Miller, D.B. Neurotoxicity of the pesticidal carbamates. Neurobehav. Toxicol. Teratol. 1982, 4, 779–787. [Google Scholar]
- Andrade, A.C.; Del Sorbo, G.; Van Nistelrooy, J.G.M.; De Waard, M.A. The ABC transporter AtrB from Aspergillus nidulans mediates resistance to all major classes of fungicides and some natural toxic compounds. Microbiology 2000, 146, 1987–1997. [Google Scholar] [CrossRef]
- Yang, C.; Hamel, C.; Vujanovic, V.; Gan, Y. Fungicide: Modes of Action and Possible Impact on Nontarget Microorganisms. ISRN Ecol. 2011, 2011, 1–8. [Google Scholar] [CrossRef]
- Zeiger, E.; Anderson, B.; Haworth, S.; Lawlor, T.; Mortelmans, K. Salmonella mutagenicity tests: V. Results from the testing of 311 chemicals. Environ. Mol. Mutagen. 1992, 19, 2–141. [Google Scholar] [CrossRef]
- Bordas, A.C.; Brady, M.S.; Siewierski, M.; Katz, S.E. In Vitro Enhancement of Antibiotic Resistance Development—Interaction of Residue Levels of Pesticides and Antibiotics. J. Food Prot. 1997, 60, 531–536. [Google Scholar] [CrossRef]
- Kleiner, D.K.; Katz, S.E.; Ward, P.-M.L. Development of in vitro Antimicrobial Resistance in Bacteria Exposed to Residue Level Exposures of Antimicrobial Drugs, Pesticides and Veterinary Drugs. Chemotherapy 2007, 53, 132–136. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef] [PubMed]
- Woodburn, A.T. Glyphosate: Production, pricing and use worldwide. Pest Manag. Sci. 2000, 56, 309–312. [Google Scholar] [CrossRef]
- Hertel, R.; Gibhardt, J.; Martienssen, M.; Kuhn, R.; Commichau, F.M. Molecular mechanisms underlying glyphosate resistance in bacteria. Environ. Microbiol. 2021, 23, 2891–2905. [Google Scholar] [CrossRef] [PubMed]
- Kurenbach, B.; Marjoshi, D.; Amábile-Cuevas, C.F.; Ferguson, G.C.; Godsoe, W.; Gibson, P.; Heinemann, J.A. Sublethal Exposure to Commercial Formulations of the Herbicides Dicamba, 2,4-Dichlorophenoxyacetic Acid, and Glyphosate Cause Changes in Antibiotic Susceptibility in Escherichia coli and Salmonella enterica serovar Typhimurium. mBio 2015, 6, e00009-15. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Reeder, R. Antibiotic use on crops in low and middle-income countries based on recommendations made by agricultural advisors. CABI Agric. Biosci. 2020, 1, 1–14. [Google Scholar] [CrossRef]



| Group | I | II | III |
|---|---|---|---|
| Restriction | None | Field trials, not for market products | Not to be used |
| Examples | Kanamycin, hygromycin | Chloramphenicol, ampicillin, streptomycin, spectinomycin, | Amikacin, tetracyclines |
| Examples | OIE | WHO | |
|---|---|---|---|
| Aminoglycosides | Streptomycin, kasugamycin (in plant protection only) | Very Critically Important Antimicrobial (VCIA) | Critically important |
| Tetracyclines | Oxytetracycline | VCIA | Highly important antimicrobial |
| Quinolones | Oxolinic acid | VCIA | Highly important antimicrobial |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, S.A.; Ferreira, J.P.; LeJeune, J.T. Antimicrobial Use and Resistance in Plant Agriculture: A One Health Perspective. Agriculture 2022, 12, 289. https://doi.org/10.3390/agriculture12020289
Miller SA, Ferreira JP, LeJeune JT. Antimicrobial Use and Resistance in Plant Agriculture: A One Health Perspective. Agriculture. 2022; 12(2):289. https://doi.org/10.3390/agriculture12020289
Chicago/Turabian StyleMiller, Sally A., Jorge Pinto Ferreira, and Jeffrey T. LeJeune. 2022. "Antimicrobial Use and Resistance in Plant Agriculture: A One Health Perspective" Agriculture 12, no. 2: 289. https://doi.org/10.3390/agriculture12020289
APA StyleMiller, S. A., Ferreira, J. P., & LeJeune, J. T. (2022). Antimicrobial Use and Resistance in Plant Agriculture: A One Health Perspective. Agriculture, 12(2), 289. https://doi.org/10.3390/agriculture12020289