
Development and Validation of SNP and InDel Markers for Pod-Shattering Tolerance in Soybean

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
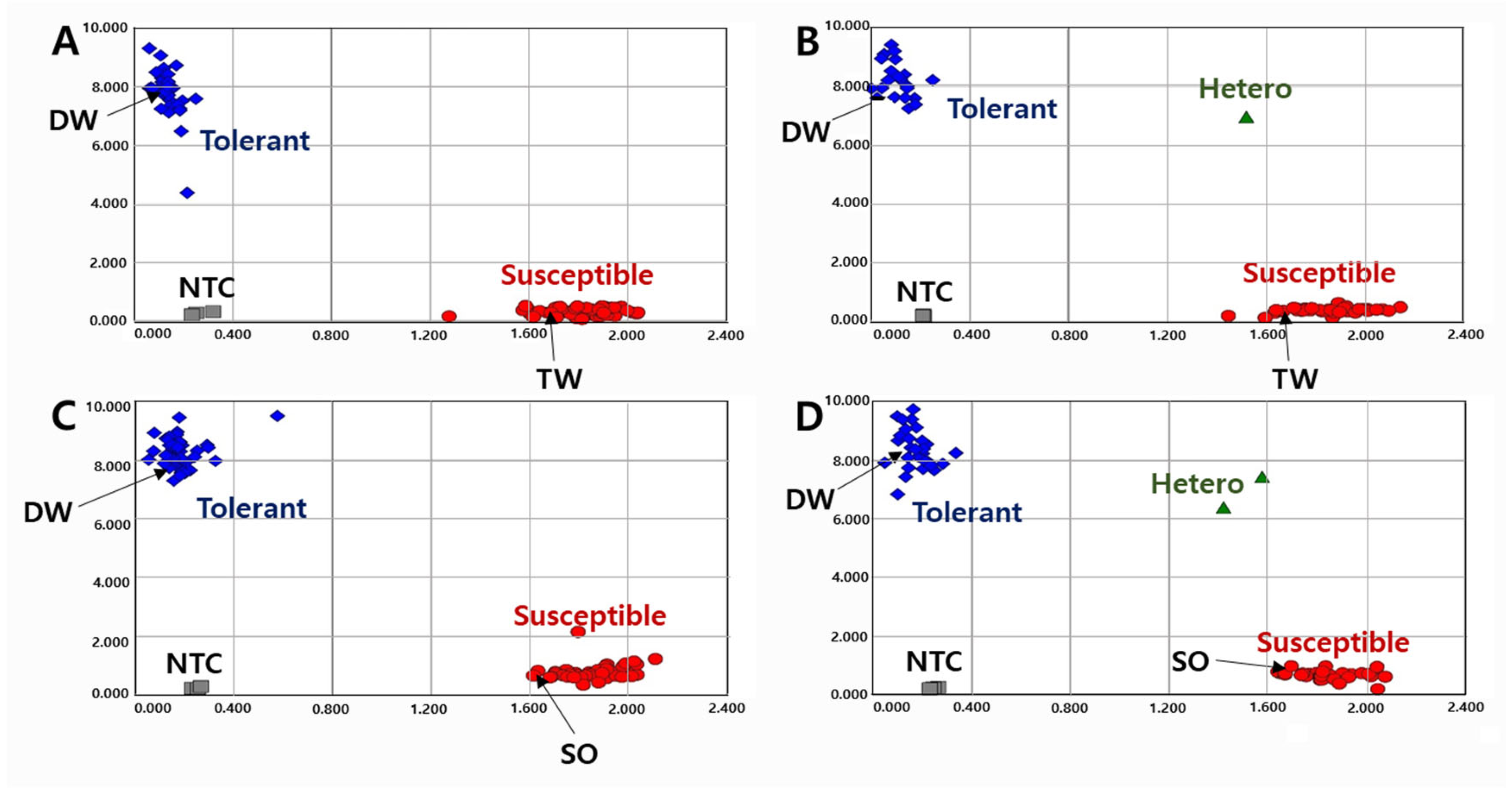
2.1. Development of KASP Markers
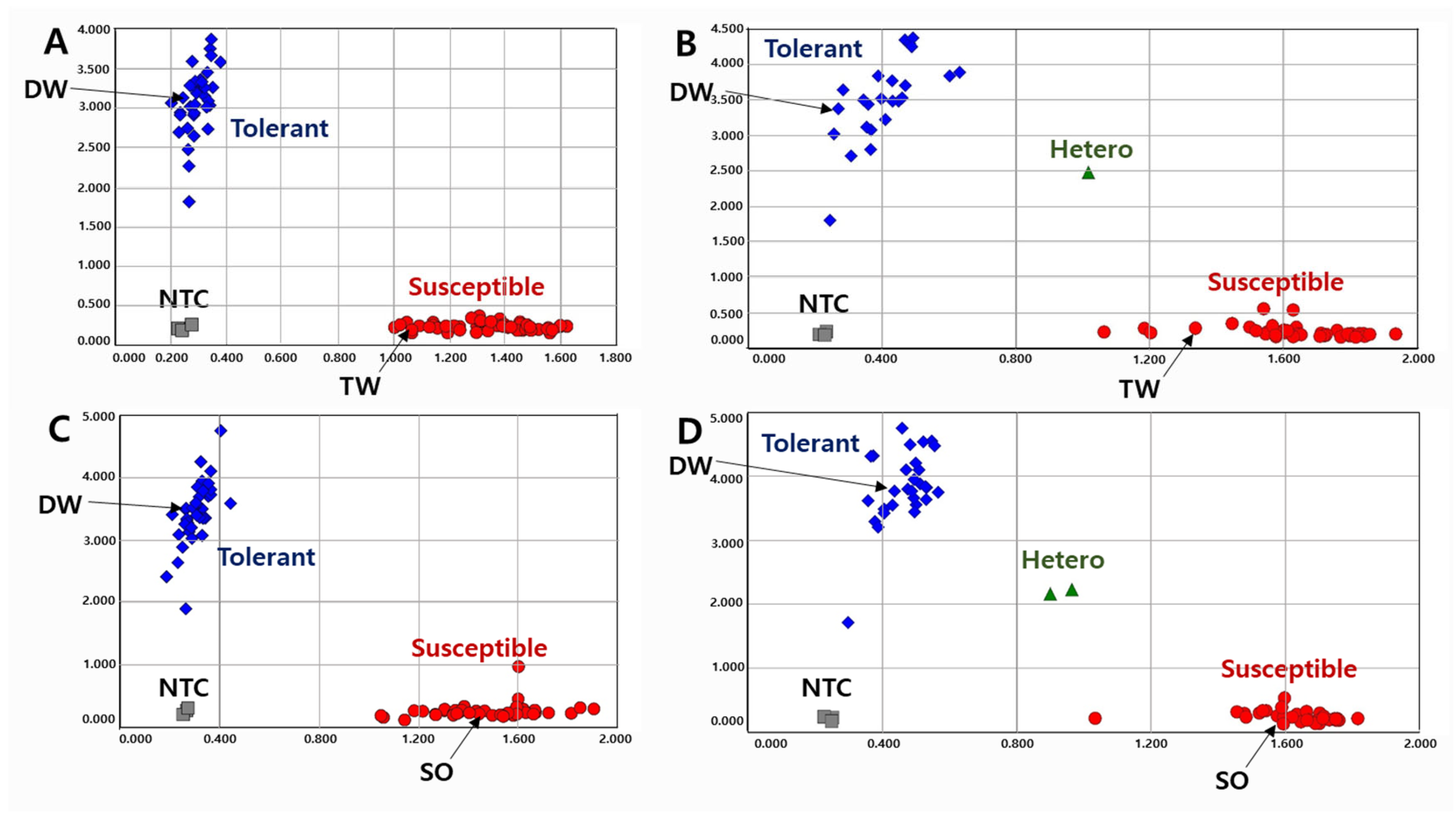
2.2. Development of InDel Markers
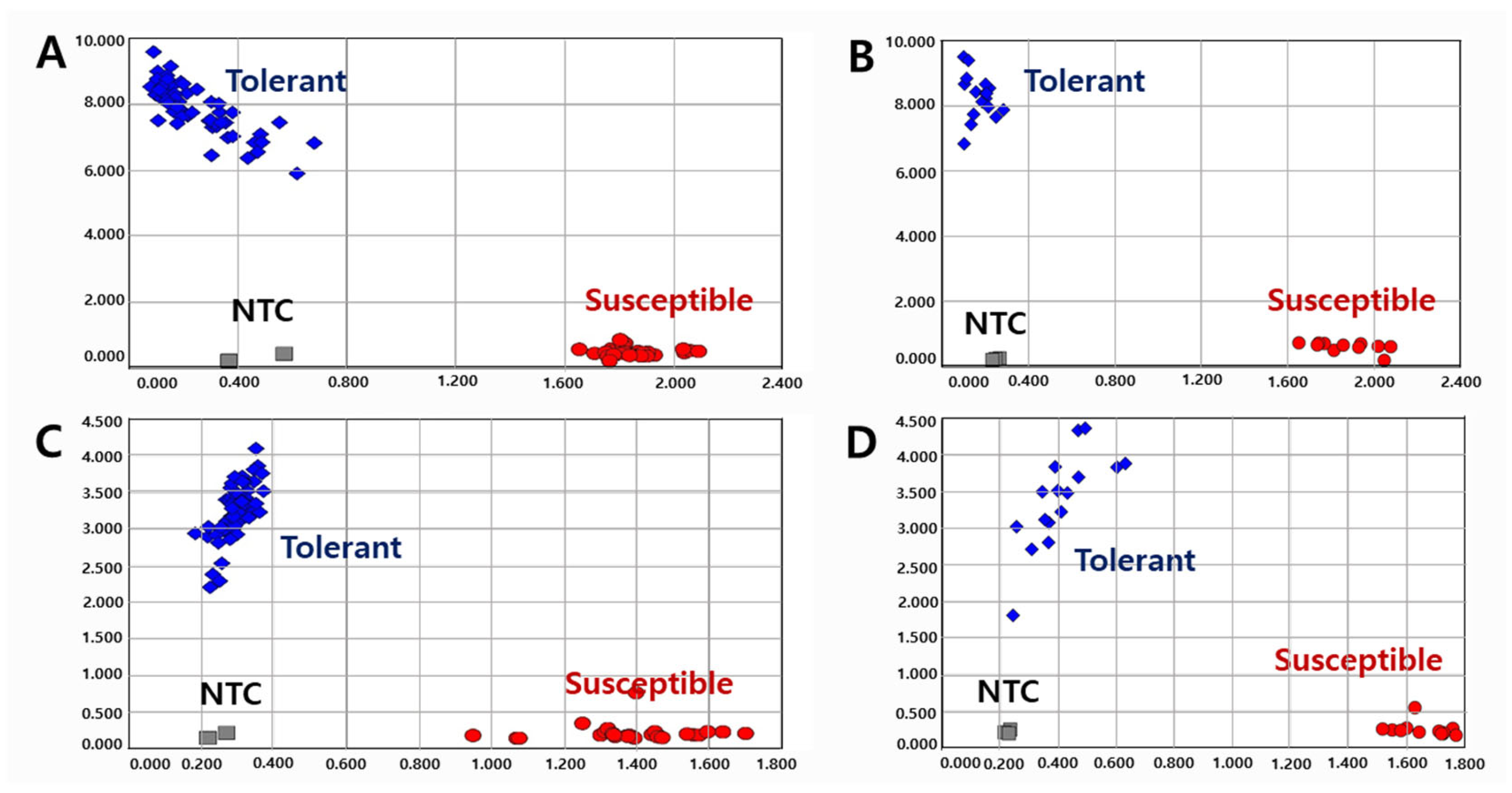
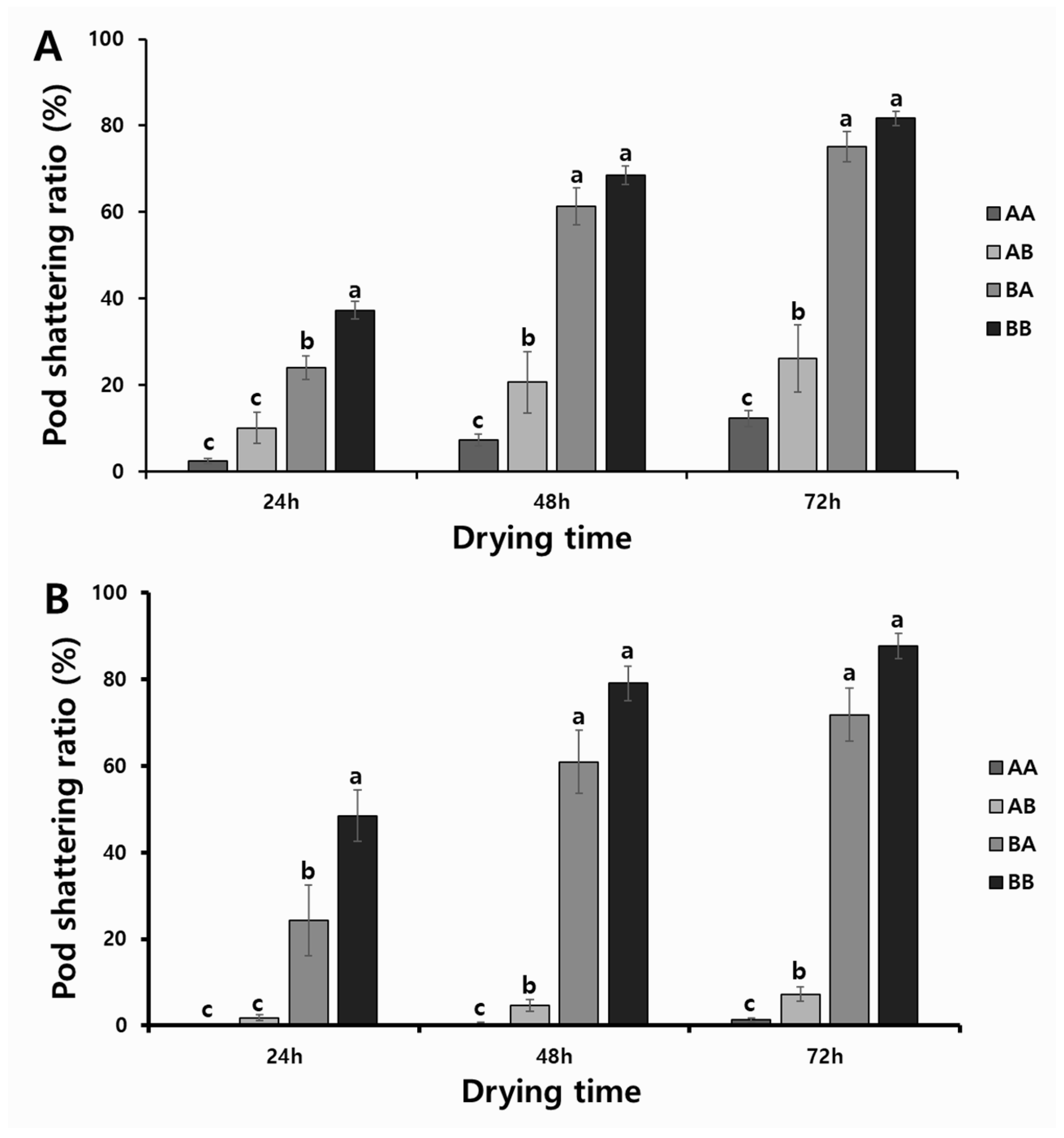
2.3. Validation of KASP and InDel Markers in RIL Populations
2.4. Validation of KASP and InDel Markers in Diverse Varieties and Elite Lines
2.5. Association between Allelic Combination and Phenotypes
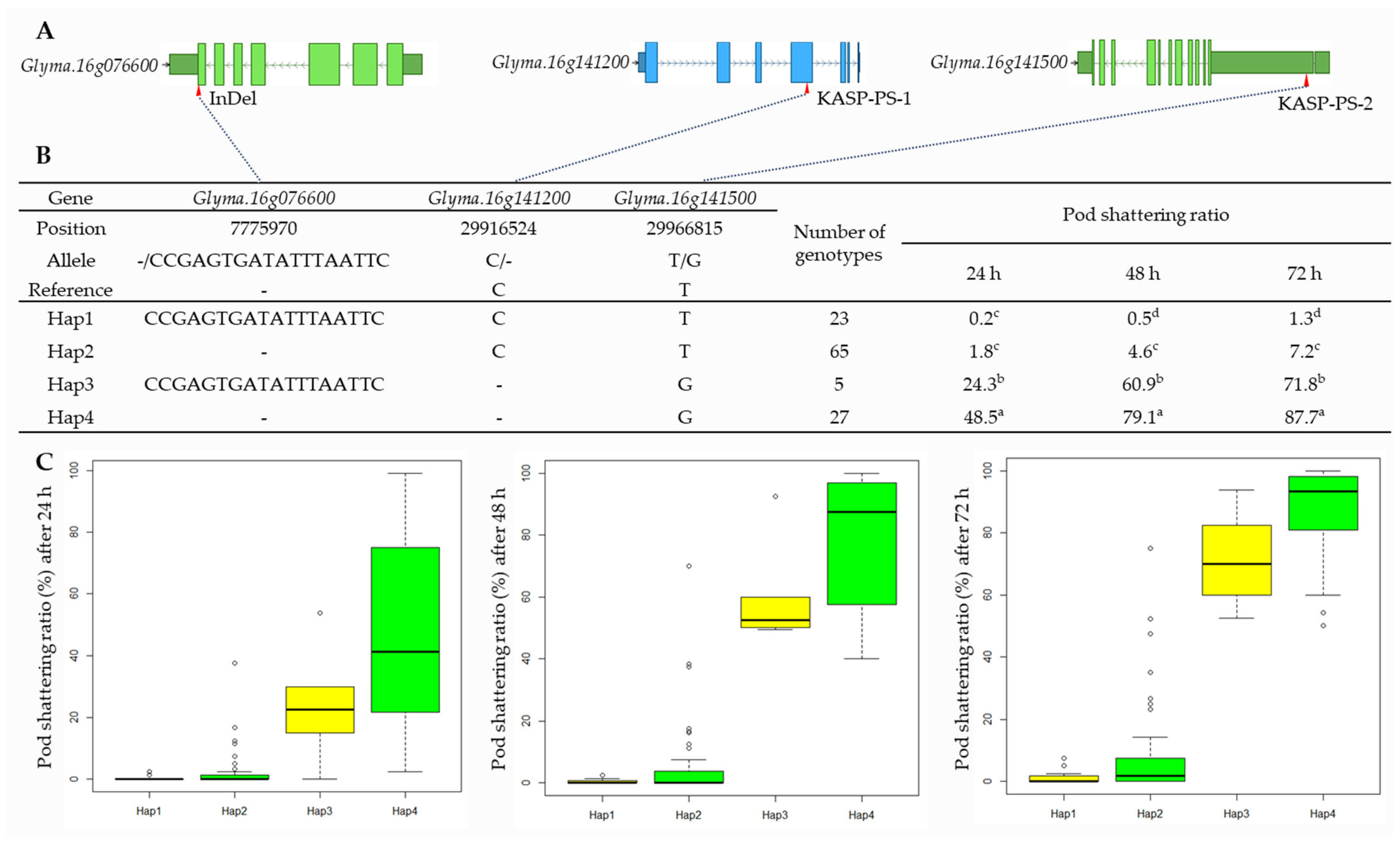
2.6. Haplotype Analysis of Candidate Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Evaluation of Pod-Shattering Tolerance
4.3. DNA Extraction from the Parental Lines and Whole-Genome Resequencing
4.4. Molecular Marker Development and Genotyping
4.5. Determination of Prediction Accuracy
4.6. Haplotype Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rey, L.; Lopez, Y.; Jaramillo, A.; White, J.W. Evaluation of a Growth Simulation Model Applied to Soybean Genotypes (GIycine Max L. Merr) under Tropical Conditions. Corpoica Cienc. Tecnol. Agropecu. 1996, 1, 16. [Google Scholar] [CrossRef] [Green Version]
- Painii Montero, V.F.; Santillán Muñoz, O.B.; Montes Escobar, K.; Garces Fiallos, F.R. Characterization of Soybean Productive Units in the Ecuadorian Coast. Cienc. Tecnol. Agropecu. 2020, 21, 1–20. [Google Scholar] [CrossRef]
- Nilmani, B.; Dhirendra, K.; Shrivastava, A.N. Studies on the Factors Affecting Pod Shattering in Soybean. Indian J. Genet. Plant Breed. 2013, 73, 270–277. [Google Scholar] [CrossRef]
- Funatsuki, H.; Suzuki, M.; Hirose, A.; Inaba, H.; Yamada, T.; Hajika, M.; Komatsu, K.; Katayama, T.; Sayama, T.; Ishimoto, M.; et al. Molecular Basis of a Shattering Resistance Boosting Global Dissemination of Soybean. Proc. Natl. Acad. Sci. USA 2014, 111, 17797–17802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukamuhabwa, P.; Dashiell, K.E.; Rubaihayo, P.; Nabasirye, M. Determination of Field Yield Loss and Effect of Environment on Pod Shattering in Soybean. Afr. Crop Sci. J. 2002, 10, 203–209. [Google Scholar]
- Zhang, L.; Bellaloui, N. Effects of Planting and Maturity Dates on Shattering Patterns under Early Soybean Production System. Am. J. Plant Sci. 2012, 3, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Tu, B.; Liu, C.; Liu, X. Pod Anatomy, Morphology and Dehiscing Forces in Pod Dehiscence of Soybean (Glycine max (L.) Merrill). Flora 2018, 248, 48–53. [Google Scholar] [CrossRef]
- Miles, C.; Wayne, M. Quantitative Trait Locus (QTL) Analysis. Nat. Educ. 2008, 1, 208. [Google Scholar]
- Visscher, P.M.; Andrew, T.; Nyholt, D.R. Genome-Wide Association Studies of Quantitative Traits with Related Individuals: Little (Power) Lost but Much to Be Gained. Eur. J. Hum. Genet. 2008, 16, 387–390. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.A.; Mian, M.A.R.; Carter, T.E.; Ashley, D.A.; Boerma, H.R. Pod Dehiscence of Soybean: Identification of Quantitative Trait Loci. J. Hered. 1997, 88, 152–154. [Google Scholar] [CrossRef]
- Kang, S.-T.; Kwak, M.; Kim, H.-K.; Choung, M.-G.; Han, W.-Y.; Baek, I.-Y.; Kim, M.Y.; Van, K.; Lee, S.-H. Population-Specific QTLs and Their Different Epistatic Interactions for Pod Dehiscence in Soybean [Glycine max (L.) Merr.]. Euphytica 2009, 166, 15–24. [Google Scholar] [CrossRef]
- Funatsuki, H.; Ishimoto, M.; Tsuji, H.; Kawaguchi, K.; Hajika, M.; Fujino, K. Simple Sequence Repeat Markers Linked to a Major QTL Controlling Pod Shattering in Soybean. Plant Breed. 2006, 125, 195–197. [Google Scholar] [CrossRef]
- Suzuki, M.; Fujino, K.; Nakamoto, Y.; Ishimoto, M.; Funatsuki, H. Fine Mapping and Development of DNA Markers for the QPDH1 Locus Associated with Pod Dehiscence in Soybean. Mol. Breed. 2010, 25, 407–418. [Google Scholar] [CrossRef]
- Gao, M.; Zhu, H. Fine Mapping of a Major Quantitative Trait Locus That Regulates Pod Shattering in Soybean. Mol. Breed. 2013, 32, 485–491. [Google Scholar] [CrossRef]
- Hu, D.; Kan, G.; Hu, W.; Li, Y.; Hao, D.; Li, X.; Yang, H.; Yang, Z.; He, X.; Huang, F.; et al. Identification of Loci and Candidate Genes Responsible for Pod Dehiscence in Soybean via Genome-Wide Association Analysis across Multiple Environments. Front. Plant Sci. 2019, 10, 811. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Han, D.; Guo, Y.; Yan, H.; Wei, Z.; Tian, Y.; Qiu, L. QTL Mapping Pod Dehiscence Resistance in Soybean (Glycine max L. Merr.) Using Specific-Locus Amplified Fragment Sequencing. Theor. Appl. Genet. 2019, 132, 2253–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.-H.; Kang, B.-K.; Dhungana, S.K.; Oh, J.-H.; Choi, M.-S.; Park, J.-H.; Shin, S.-O.; Kim, H.-S.; Baek, I.-Y.; Sung, J.-S.; et al. QTL Mapping and Candidate Gene Analysis for Pod Shattering Tolerance in Soybean (Glycine max). Plants 2020, 9, 1163. [Google Scholar] [CrossRef]
- Parker, T.A.; Berny Mier y Teran, J.C.; Palkovic, A.; Jernstedt, J.; Gepts, P. Pod Indehiscence Is a Domestication and Aridity Resilience Trait in Common Bean. New Phytol. 2020, 225, 558–570. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.T.; Kim, H.K.; Baek, I.Y.; Chung, M.G.; Han, W.Y.; Shin, D.C.; Lee, S.-H. Genetic Analysis of Pod Dehiscence in Soybean. Korean J. Crop Sci. 2005, 50, 281–285. [Google Scholar]
- Tukamuhabwa, P.; Rubaihayo, P.; Dashiell, K.E. Genetic Components of Pod Shattering in Soybean. Euphytica 2002, 125, 29–34. [Google Scholar] [CrossRef]
- Zhao, S.; Li, A.; Li, C.; Xia, H.; Zhao, C.; Zhang, Y.; Hou, L.; Wang, X. Development and Application of KASP Marker for High Throughput Detection of AhFAD2 Mutation in Peanut. Electron. J. Biotechnol. 2017, 25, 9–12. [Google Scholar] [CrossRef]
- Wang, D.; Yang, T.; Liu, R.; Li, N.; Wang, X.; Sarker, A.; Zhang, X.; Li, R.; Pu, Y.; Li, G.; et al. RNA-Seq Analysis and Development of SSR and KASP Markers in Lentil (Lens culinaris Medikus Subsp. culinaris). Crop J. 2020, 8, 953–965. [Google Scholar] [CrossRef]
- Hechanova, S.L.; Bhattarai, K.; Simon, E.V.; Clave, G.; Karunarathne, P.; Ahn, E.-K.; Li, C.-P.; Lee, J.-S.; Kohli, A.; Hamilton, N.R.S.; et al. Development of a Genome-Wide InDel Marker Set for Allele Discrimination between Rice (Oryza sativa) and the Other Seven AA-Genome Oryza Species. Sci. Rep. 2021, 11, 8962. [Google Scholar] [CrossRef] [PubMed]
- Paudel, L.; Clevenger, J.; McGregor, C. Refining of the Egusi Locus in Watermelon Using KASP Assays. Sci. Hortic. 2019, 257, 108665. [Google Scholar] [CrossRef]
- Devran, Z.; Göknur, A. Development and Validation of a SNP-Based KASP Assay for Rapid Identification of Aphelenchoides besseyi Christie, 1942. Crop Prot. 2020, 136, 105235. [Google Scholar] [CrossRef]
- Shi, Z.; Liu, S.; Noe, J.; Arelli, P.; Meksem, K.; Li, Z. SNP Identification and Marker Assay Development for High-Throughput Selection of Soybean Cyst Nematode Resistance. BMC Genom. 2015, 16, 314. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.T.; Steketee, C.J.; Boehm, J.D.; Noe, J.; Li, Z. Genome-Wide Association Analysis Pinpoints Additional Major Genomic Regions Conferring Resistance to Soybean Cyst Nematode (Heterodera glycines Ichinohe). Front. Plant Sci. 2019, 10, 401. [Google Scholar] [CrossRef]
- Pham, A.-T.; Harris, D.K.; Buck, J.; Hoskins, A.; Serrano, J.; Abdel-Haleem, H.; Cregan, P.; Song, Q.; Boerma, H.R.; Li, Z. Fine Mapping and Characterization of Candidate Genes That Control Resistance to Cercospora sojina K. Hara in Two Soybean Germplasm Accessions. PLoS ONE 2015, 10, e0126753. [Google Scholar] [CrossRef]
- Cheng, P.; Gedling, C.R.; Patil, G.; Vuong, T.D.; Shannon, J.G.; Dorrance, A.E.; Nguyen, H.T. Genetic Mapping and Haplotype Analysis of a Locus for Quantitative Resistance to Fusarium graminearum in Soybean Accession PI 567516C. Theor. Appl. Genet. 2017, 130, 999–1010. [Google Scholar] [CrossRef]
- Miranda, C.; Culp, C.; Škrabišová, M.; Joshi, T.; Belzile, F.; Grant, D.M.; Bilyeu, K. Molecular Tools for Detecting Pdh1 Can Improve Soybean Breeding Efficiency by Reducing Yield Losses Due to Pod Shatter. Mol. Breed. 2019, 39, 27. [Google Scholar] [CrossRef]
- Kim, J.-M.; Kim, K.-H.; Jung, J.; Kang, B.K.; Lee, J.; Ha, B.-K.; Kang, S. Validation of Marker-Assisted Selection in Soybean Breeding Program for Pod Shattering Resistance. Euphytica 2020, 216, 166. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, K.R.; Ha, B.-K.; Kang, S. Identification of SNPs Tightly Linked to the QTL for Pod Shattering in Soybean. Mol. Breed. 2017, 37, 54. [Google Scholar] [CrossRef]
- Tiwari, S.; Bhatnagar, P. Pod Shattering as Related to Other Agronomic Attributes in Soybean. Trop. Agric. 1991, 68, 102–103. [Google Scholar]
- Umar, F.A.; Mohammed, M.S.; Oyekunle, M.; Usman, I.S.; Ishaq, M.N.; Dachi, S.N. Estimates of Combining Ability for Resistance to Pod Shattering in Soybean (Glycine max (L.) Merrill) Genotypes. J. Plant Breed. Crop Sci. 2017, 9, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Plomin, R.; DeFries, J.C.; McClearn, G.E.; McGuffin, P. Behavioral Genetics; Worth Publishers: New York, NY, USA, 2001. [Google Scholar]
- Tsuchiya, T.; Sunada, K. Breeding Studies on Pod Shattering in Soybeans. II. Methods of Testing for Shattering and Varietal Differences. Soybean Abstr. 1980, 3, 1706. [Google Scholar]
- Tukamuhabwa, P.; Rubaihayo, P.R.; Dashiell, K.; Adipala, E. Inheritance to Pod Shattering in Soybean. Afr. J. Crop Sci. 2000, 8, 203–211. [Google Scholar]
- Tiwari, S.; Bhatnagar, P. Consistent Resistance for Pod Shattering in Soybean Varieties. Indian J. Agric. Sci. 1993, 63, 173–174. [Google Scholar]
- Bhor, T.J.; Chimote, V.P.; Deshmukh, M.P. Inheritance of Pod Shattering in Soybean (Glycine max (L.) Merrill). Electron. J. Plant Breed. 2014, 5, 671–676. [Google Scholar]
- Han, D.; Han, J.; Jiang, S.; Su, B.; Zhang, B.; Liu, Z.; Yan, H.; Qiu, L.-J. Shattering-Resistance of an Elite Soybean Variety ‘Heihe 43’ and Identification of Shattering-Resistant Genes. Euphytica 2021, 217, 120. [Google Scholar] [CrossRef]
- Dong, Y.; Yang, X.; Liu, J.; Wang, B.-H.; Liu, B.-L.; Wang, Y.-Z. Pod Shattering Resistance Associated with Domestication Is Mediated by a NAC Gene in Soybean. Nat. Commun. 2014, 5, 3352. [Google Scholar] [CrossRef] [Green Version]
- Satoh, R.; Fujita, Y.; Nakashima, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. A Novel Subgroup of BZIP Proteins Functions as Transcriptional Activators in Hypoosmolarity-Responsive Expression of the ProDH Gene in Arabidopsis. Plant Cell Physiol. 2004, 45, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavi Kishor, P.B. Role of Proline in Cell Wall Synthesis and Plant Development and Its Implications in Plant Ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihsan, M.Z.; Ahmad, S.J.N.; Shah, Z.H.; Rehman, H.M.; Aslam, Z.; Ahuja, I.; Bones, A.M.; Ahmad, J.N. Gene Mining for Proline Based Signaling Proteins in Cell Wall of Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funck, D.; Eckard, S.; Müller, G. Non-Redundant Functions of Two Proline Dehydrogenase Isoforms in Arabidopsis. BMC Plant Biol. 2010, 10, 70. [Google Scholar] [CrossRef] [Green Version]
- Dong, R.; Dong, D.; Luo, D.; Zhou, Q.; Chai, X.; Zhang, J.; Xie, W.; Liu, W.; Dong, Y.; Wang, Y.; et al. Transcriptome Analyses Reveal Candidate Pod Shattering-Associated Genes Involved in the Pod Ventral Sutures of Common Vetch (Vicia sativa L.). Front. Plant Sci. 2017, 8, 649. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Yan, L.; Dong, R.; Liu, W.; Wang, Y.; Liu, Z. Evaluation and Analysis of Pod Dehiscence Factors in Shatter-Susceptible and Shatter-Resistant Common Vetch. Crop Sci. 2017, 57, 2770–2776. [Google Scholar] [CrossRef]
- Ehlting, J.; Mattheus, N.; Aeschliman, D.S.; Li, E.; Hamberger, B.; Cullis, I.F.; Zhuang, J.; Kaneda, M.; Mansfield, S.D.; Samuels, L.; et al. Global Transcript Profiling of Primary Stems from Arabidopsis thaliana Identifies Candidate Genes for Missing Links in Lignin Biosynthesis and Transcriptional Regulators of Fiber Differentiation. Plant J. 2005, 42, 618–640. [Google Scholar] [CrossRef]
- Tucker, M.L.; Whitelaw, C.A.; Lyssenko, N.N.; Nath, P. Functional Analysis of Regulatory Elements in the Gene Promoter for an Abscission-Specific Cellulase from Bean and Isolation, Expression, and Binding Affinity of Three TGA-Type Basic Leucine Zipper Transcription Factors. Plant Physiol. 2002, 130, 1487–1496. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, J.; Zhang, Z.; Xie, W. Elymus Nutans Genes for Seed Shattering and Candidate Gene-Derived EST-SSR Markers for Germplasm Evaluation. BMC Plant Biol. 2019, 19, 102. [Google Scholar] [CrossRef]
- Meyer, K.; Leube, M.P.; Grill, E. A Protein Phosphatase 2C Involved in ABA Signal Transduction in Arabidopsis thaliana. Science 1994, 264, 1452–1455. [Google Scholar] [CrossRef]
- Saito, S.; Hirai, N.; Matsumoto, C.; Ohigashi, H.; Ohta, D.; Sakata, K.; Mizutani, M. Arabidopsis CYP707A s Encode (+)-Abscisic Acid 8′-Hydroxylase, a Key Enzyme in the Oxidative Catabolism of Abscisic Acid. Plant Physiol. 2004, 134, 1439–1449. [Google Scholar] [CrossRef] [Green Version]
- Jaradat, M.R.; Ruegger, M.; Bowling, A.; Butler, H.; Cutler, A.J. A Comprehensive Transcriptome Analysis of Silique Development and Dehiscence in Arabidopsis and Brassica Integrating Genotypic, Interspecies and Developmental Comparisons. GM Crops Food 2014, 5, 302–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.H.; Kang, B.K.; Kim, H.T.; Kim, H.S.; Choi, M.S.; Oh, J.H.; Shin, S.O.; Baek, I.Y.; Kwak, D.Y. Variation in Pod Shattering in a RIL Population and Selection for Pod Shattering Tolerance in Soybean [Glycine max (L.) Merr]. Korean J. Crop Sci. 2019, 64, 414–421. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome Sequence of the Palaeopolyploid Soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; San, M.L.; Jang, S.-G.; Lee, J.-H.; Kim, N.-E.; Lee, A.-R.; Park, S.-Y.; Cao, F.-Y.; Chin, J.-H.; Kwon, S.-W. Genome-Wide Association Study of Root System Development at Seedling Stage in Rice. Genes 2020, 11, 1395. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Model | SNP Position (bp) | Polymorphic Site | SNP Information | Amino Acid Change |
|---|---|---|---|---|
| Glyma.16g141200 | 29,916,524 | 3′ UTR | Deletion (TC > T) | - |
| Glyma.16g141500 | 29,964,216 | 5′ UTR | A > C | - |
| Glyma.16g141500 | 29,966,815 | 5′ UTR | T > G | - |
| Glyma.16g141600 | 29,970,894 | Coding region | T > C Nonsynonymous | (Asp > Gly) |
| Glyma.16g141600 | 29,970,957 | Coding region | T > C Nonsynonymous | (Asn > Ser) |
| Gene Model | Position (bp) | Polymorphism Site | Codon Change | Amino Acid Change |
|---|---|---|---|---|
| Glyma.16g076600 | 7,775,892 | Coding region | C > T Nonsynonymous | (Glu > Lys) |
| 7,775,945 | Coding region | T > A Nonsynonymous | (Lys > Met) | |
| 7,775,948 | Coding region | A > G Nonsynonymous | (Ile > Thr) | |
| 7,775,970 | Coding region | Insertion (18 bp) | Stop gained and disruptive inframe insertion | |
| 7,776,045 | Coding region | C > T Nonsynonymous | (Met > Ile) | |
| 7,777,575 | Coding region | Deletion (3 bp) Nonsynonymous | Inframe deletion (Asn > -) |
| Marker Type | Population | Genotype | Pod Shattering Ratio (%) | ||
|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | |||
| KASP | DT | P1 | 1.6a | 5.1a | 9.0a |
| P2 | 25.9b | 55.0b | 75.1b | ||
| DS | P1 | 4.9a | 12.3a | 18.3a | |
| P2 | 44.0b | 80.4b | 86.3b | ||
| InDel | DT | P1 | 7.4a | 20.3a | 27.9a |
| P2 | 24.7b | 49.5b | 69.0b | ||
| DS | P1 | 7.6a | 19.7a | 26.4a | |
| P2 | 43.0b | 75.9b | 81.2b | ||
| Marker Type | Genotype | Pod-Shattering Ratio (%) | ||
|---|---|---|---|---|
| 24 h | 48 h | 72 h | ||
| KASP | P1 | 1.4a | 3.6a | 5.7a |
| P2 | 44.6b | 76.2b | 85.1b | |
| InDel | P1 | 4.7a | 11.6a | 14.4a |
| P2 | 15.0b | 26.1b | 30.7b | |
| Genotypes | RIL Populations | Varieties and Elite Lines | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |||||||
| Tol | Sus | Tol | Sus | Tol | Sus | Tol | Sus | Tol | Sus | Tol | Sus | |
| X | 116 | 12 | 101 | 27 | 82 | 46 | 84 | 4 | 80 | 8 | 73 | 12 |
| Y | 31 | 135 | 4 | 162 | 1 | 165 | 4 | 27 | 0 | 31 | 0 | 31 |
| Accuracy (%) | 86.0 | 88.2 | 81.7 | 91.3 | 95.5 | 92.9 | ||||||
| AA | 101 | 7 | 90 | 18 | 73 | 35 | 22 | 0 | 22 | 0 | 22 | 0 |
| BB | 22 | 109 | 2 | 129 | 1 | 130 | 0 | 26 | 0 | 26 | 0 | 26 |
| Accuracy (%) | 88.4 | 90.9 | 83.4 | 100 | 100 | 100 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, J.-H.; Dhungana, S.K.; Kang, B.-K.; Baek, I.-Y.; Sung, J.-S.; Ko, J.-Y.; Jung, C.-S.; Kim, K.-S.; Jun, T.-H. Development and Validation of SNP and InDel Markers for Pod-Shattering Tolerance in Soybean. Int. J. Mol. Sci. 2022, 23, 2382. https://doi.org/10.3390/ijms23042382
Seo J-H, Dhungana SK, Kang B-K, Baek I-Y, Sung J-S, Ko J-Y, Jung C-S, Kim K-S, Jun T-H. Development and Validation of SNP and InDel Markers for Pod-Shattering Tolerance in Soybean. International Journal of Molecular Sciences. 2022; 23(4):2382. https://doi.org/10.3390/ijms23042382
Chicago/Turabian StyleSeo, Jeong-Hyun, Sanjeev Kumar Dhungana, Beom-Kyu Kang, In-Youl Baek, Jung-Sook Sung, Jee-Yeon Ko, Chan-Sik Jung, Ki-Seung Kim, and Tae-Hwan Jun. 2022. "Development and Validation of SNP and InDel Markers for Pod-Shattering Tolerance in Soybean" International Journal of Molecular Sciences 23, no. 4: 2382. https://doi.org/10.3390/ijms23042382
APA StyleSeo, J.-H., Dhungana, S. K., Kang, B.-K., Baek, I.-Y., Sung, J.-S., Ko, J.-Y., Jung, C.-S., Kim, K.-S., & Jun, T.-H. (2022). Development and Validation of SNP and InDel Markers for Pod-Shattering Tolerance in Soybean. International Journal of Molecular Sciences, 23(4), 2382. https://doi.org/10.3390/ijms23042382