
Polyhydroxyalkanoate Production by Caenibius tardaugens from Steroidal Endocrine Disruptors



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strains and Culture Conditions
2.3. Polymer Analysis
2.4. Characterization of PHA Polymers by Nuclear Magnetic Resonance (NMR)
2.5. Transmission Electron Microscopy (TEM)
2.6. Bioinformatics Analysis
3. Results
3.1. Analysis of Genes Related to PHB Synthesis in C. tardaugens
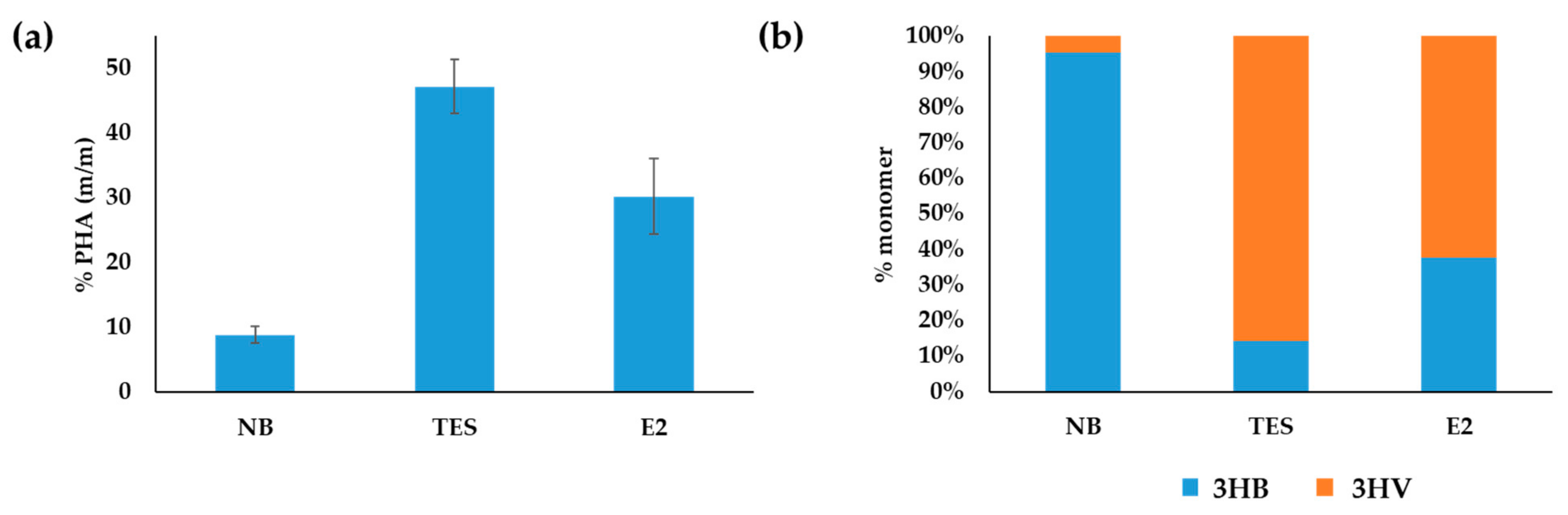
3.2. Production of PHAs from Steroids in C. tardaugens NBRC 16725
3.3. Distribution of PHA Genes in Phylogenetically Related Bacteria
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bradley, P.M.; Chapelle, F.H.; Barber, L.B.; McMahon, P.B.; Gray, J.L.; Kolpin, D.W. Biodegradation of 17β-estradiol, estrone, and testosterone in stream sediments. In Proceedings of the 10th International in Situ and On-Site Bioremediation Symposium, In Situ and On-Site Bioremediation-2009, Baltimore, MD, USA, 5–8 May 2009. [Google Scholar]
- Chen, Y.-L.; Wang, C.-H.; Yang, F.-C.; Ismail, W.; Wang, P.-H.; Shih, C.-J.; Wu, Y.-C.; Chiang, Y.-R. Identification of Comamonas testosteroni as an androgen degrader in sewage. Sci. Rep. 2016, 6, 35386. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Casey, F.X.M.; Hakk, H.; Larsen, G.L. Persistence and fate of 17β-estradiol and testosterone in agricultural soils. Chemosphere 2007, 67, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Wu, S.; Chang, H.; Hu, J. Behaviors of glucocorticoids, androgens and progestogens in a municipal sewage treatment plant: Comparison to estrogens. Environ. Sci. Technol. 2011, 45, 2725–2733. [Google Scholar] [CrossRef] [PubMed]
- Teles, M.; Gravato, C.; Pacheco, M.; Santos, M.A. Juvenile sea bass biotransformation, genotoxic and endocrine responses to β-naphthoflavone, 4-nonylphenol and 17β-estradiol individual and combined exposures. Chemosphere 2004, 57, 147–158. [Google Scholar] [CrossRef]
- Ibero, J.; Galán, B.; Rivero-Buceta, V.; García, J.L. Unraveling the 17β-estradiol degradation pathway in Novosphingobium tardaugens NBRC 16725. Front. Microbiol. 2020, 11, 588300. [Google Scholar] [CrossRef]
- Fujii, K.; Satomi, M.; Morita, N.; Motomura, T.; Tanaka, T.; Kikuchi, S. Novosphingobium tardaugens sp. nov., an oestradiol-degrading bacterium isolated from activated sludge of a sewage treatment plant in Tokyo. Int. J. Syst. Evol. Microbiol. 2003, 53, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Ibero, J.; Sanz, D.; Galán, B.; Díaz, E.; García, J.L. High-quality whole-genome sequence of an estradiol-degrading strain, Novosphingobium tardaugens NBRC 16725. Microb. Resour. Annoucements 2019, 8, e01715-18. [Google Scholar] [CrossRef] [Green Version]
- Ibero, J.; Galán, B.; Díaz, E.; García, J.L. Testosterone degradative pathway of Novosphingobium tardaugens. Genes 2019, 10, 871. [Google Scholar] [CrossRef] [Green Version]
- Ibero, J.; Galán, B.; García, J.L. Identification of the Edcr estrogen-dependent repressor in caenibius tardaugens NBRC 16725: Construction of a cellular estradiol biosensor. Genes 2021, 12, 1846. [Google Scholar] [CrossRef]
- Coombe, R.G.; Tsong, Y.Y.; Hamilton, P.B.; Sih, C.J. Mechanisms of steroid oxidation by microorganisms. X. Oxidative cleavage of estrone. J. Biol. Chem. 1966, 241, 1587–1595. [Google Scholar] [CrossRef]
- Chen, Y.L.; Yu, C.P.; Lee, T.H.; Goh, K.S.; Chu, K.H.; Wang, P.H.; Ismail, W.; Shih, C.J.; Chiang, Y.R. Biochemical mechanisms and catabolic enzymes involved in bacterial estrogen degradation pathways. Cell Chem. Biol. 2017, 24, 712–724.e7. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Lee, T.-H.; Chen, Y.-L.; Wang, Y.-S.; Wang, P.-H.; Yu, C.-P.; Chu, K.-H.; Chiang, Y.-R. Metabolites involved in aerobic degradation of the A and B rings of estrogen. Appl. Environ. Microbiol. 2018, 85, e02223-18. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Liu, J.; Williams, M.A.; Ling, W.; Sun, K.; Lu, C.; Gao, Y.; Waigi, M.G. Metabolism of 17β-estradiol by Novosphingobium sp. ES2-1 as probed via HRMS combined with 13C3-labeling. J. Hazard. Mater. 2020, 389, 121875. [Google Scholar] [CrossRef]
- Madison, L.L.; Huisman, G.W. Metabolic engineering of poly(3-hydroxyalkanoates): From DNA to plastic. Microbiol. Mol. Biol. Rev. 1999, 63, 21–53. [Google Scholar] [CrossRef] [Green Version]
- Prieto, M.A. From oil to bioplastics, a dream come true? J. Bacteriol. 2007, 189, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Raza, Z.A.; Abid, S.; Banat, I.M. Polyhydroxyalkanoates: Characteristics, production, recent developments and applications. Int. Biodeterior. Biodegrad. 2018, 126, 45–56. [Google Scholar] [CrossRef]
- Choi, S.Y.; Cho, I.J.; Lee, Y.; Kim, Y.J.; Kim, K.J.; Lee, S.Y. Microbial Polyhydroxyalkanoates and Nonnatural Polyesters. Adv. Mater. 2020, 32, 1907138. [Google Scholar] [CrossRef]
- Dürre, P.; Eikmanns, B.J. C1-carbon sources for chemical and fuel production by microbial gas fermentation. Curr. Opin. Biotechnol. 2015, 35, 63–72. [Google Scholar] [CrossRef]
- De Smet, M.J.; Eggink, G.; Witholt, B.; Kingma, J.; Wynberg, H. Characterization of intracellular inclusions formed by Pseudomonas oleovorans during growth on octane. J. Bacteriol. 1983, 154, 870–878. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.Y.; Kim, D.Y.; Chung, M.G.; Lee, S.H.; Park, H.Y.; Rhee, Y.H. Biosynthesis of medium-chain-length poly(3-hydroxyalkanoates) by volatile aromatic hydrocarbons-degrading Pseudomonas fulva TY16. Bioresour. Technol. 2010, 101, 8485–8488. [Google Scholar] [CrossRef]
- Tobin, K.M.; O’Connor, K.E. Polyhydroxyalkanoate accumulating diversity of Pseudomonas species utilising aromatic hydrocarbons. FEMS Microbiol. Lett. 2005, 253, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Nikodinovic, J.; Kenny, S.T.; Babu, R.P.; Woods, T.; Blau, W.J.; O’Connor, K.E. The conversion of BTEX compounds by single and defined mixed cultures to medium-chain-length polyhydroxyalkanoate. Appl. Microbiol. Biotechnol. 2008, 80, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Sanz, D.; García, J.L.; Díaz, E. Expanding the current knowledge and biotechnological applications of the oxygen-independent ortho-phthalate degradation pathway. Environ. Microbiol. 2020, 22, 3478–3493. [Google Scholar] [CrossRef] [PubMed]
- Takabatake, H.; Satoh, H.; Mino, T.; Matsuo, T. PHA (polyhydroxyalkanoate) production potential of activated sludge treating wastewater. Water Sci. Technol. 2002, 45, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Teeka, J.; Imai, T.; Reungsang, A.; Cheng, X.; Yuliani, E.; Thiantanankul, J.; Poomipuk, N.; Yamaguchi, J.; Jeenanong, A.; Higuchi, T.; et al. Characterization of polyhydroxyalkanoates (PHAs) biosynthesis by isolated Novosphingobium sp. THA-AIK7 using crude glycerol. J. Ind. Microbiol. Biotechnol. 2012, 39, 749–758. [Google Scholar] [CrossRef]
- Godoy, F.; Vancanneyt, M.; Martínez, M.; Steinbüchel, A.; Swings, J.; Rehm, B.H.A. Sphingopyxis chilensis sp. nov., a chlorophenol-degrading bacterium that accumulates polyhydroxyalkanoate, and transfer of Sphingomonas alaskensis to Sphingopyxis alaskensis comb. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 473–477. [Google Scholar] [CrossRef]
- Addison, S.L.; Foote, S.M.; Reid, N.M.; Lloyd-Jones, G. Novosphingobium nitrogenifigens sp. nov., a polyhydroxyalkanoate-accumulating diazotroph isolated from a New Zealand pulp and paper wastewater. Int. J. Syst. Evol. Microbiol. 2007, 57, 2467–2471. [Google Scholar] [CrossRef]
- Hördt, A.; López, M.G.; Meier-Kolthoff, J.P.; Schleuning, M.; Weinhold, L.M.; Tindall, B.J.; Gronow, S.; Kyrpides, N.C.; Woyke, T.; Göker, M. Analysis of 1000+ type-strain genomes substantially improves taxonomic classification of alphaproteobacteria. Front. Microbiol. 2020, 11, 468. [Google Scholar] [CrossRef]
- Moldes, C.; García, P.; García, J.L.; Prieto, M.A. In vivo immobilization of fusion proteins on bioplastics by the novel tag BioF. Appl. Environ. Microbiol. 2004, 70, 3205–3212. [Google Scholar] [CrossRef] [Green Version]
- Lageveen, R.G.; Huisman, G.W.; Preusting, H.; Ketelaar, P.; Eggink, G.; Witholt, B. Formation of polyesters by Pseudomonas oleovorans: Effect of substrates on formation and composition of poly-(R)-3-hydroxyalkanoates and poly-(R)-3-hydroxyalkenoates. Appl. Environ. Microbiol. 1988, 54, 2924–2932. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Luo, G.; Zhou, X.R.; Chen, G.Q. Biosynthesis of poly(3-hydroxydecanoate) and 3-hydroxydodecanoate dominating polyhydroxyalkanoates by β-oxidation pathway inhibited Pseudomonas putida. Metab. Eng. 2011, 13, 11–17. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Możejko-Ciesielska, J.; Kiewisz, R. Bacterial polyhydroxyalkanoates: Still fabulous? Microbiol. Res. 2016, 192, 271–282. [Google Scholar] [CrossRef]
- Strabala, T.J.; Macdonald, L.; Liu, V.; Smit, A.M. Draft genome sequence of Novosphingobium nitrogenifigens Y88 T. J. Bacteriol. 2012, 194, 201. [Google Scholar] [CrossRef] [Green Version]
- Little, G.T.; Ehsaan, M.; Arenas-López, C.; Jawed, K.; Winzer, K.; Kovacs, K.; Minton, N.P. Complete genome sequence of Cupriavidus necator H16 (DSM 428). Microbiol. Resour. Announc. 2019, 8, e00814-19. [Google Scholar] [CrossRef] [Green Version]
- Pohlmann, A.; Fricke, W.F.; Reinecke, F.; Kusian, B.; Liesegang, H.; Cramm, R.; Eitinger, T.; Ewering, C.; Pötter, M.; Schwartz, E.; et al. Genome sequence of the bioplastic-producing “Knallgas” bacterium Ralstonia eutropha H16. Nat. Biotechnol. 2006, 24, 1257–1262. [Google Scholar] [CrossRef] [Green Version]
- Galán, B.; Dinjaski, N.; Maestro, B.; De Eugenio, L.I.; Escapa, I.F.; Sanz, J.M.; García, J.L.; Prieto, M.A. Nucleoid-associated PhaF phasin drives intracellular location and segregation of polyhydroxyalkanoate granules in Pseudomonas putida KT2442. Mol. Microbiol. 2011, 79, 402–418. [Google Scholar] [CrossRef]
- De Eugenio, L.I.; Escapa, I.F.; Morales, V.; Dinjaski, N.; Galán, B.; García, J.L.; Prieto, M.A. The turnover of medium-chain-length polyhydroxyalkanoates in Pseudomonas putida KT2442 and the fundamental role of PhaZ depolymerase for the metabolic balance. Environ. Microbiol. 2010, 12, 207–221. [Google Scholar] [CrossRef]
- Doi, Y.; Kunioka, M.; Nakamura, Y.; Soga, K. Nuclear magnetic resonance studies on poly(β-hydroxybutyrate) and a copolyester of β-hydroxybutyrate and β-hydroxyvalerate isolated from Alcaligenes eutrophus H16. Macromolecules 1986, 19, 2860–2864. [Google Scholar] [CrossRef]
- Bluhm, T.L.; Hamer, G.K.; Marchessault, R.H.; Fyfe, C.A.; Veregin, R.P. Isodimorphism in bacterial poly(β-hydroxybutyrate-co-β-hydroxyvalerate). Macromolecules 2002, 19, 2871–2876. [Google Scholar] [CrossRef]
- Smit, A.M.; Strabala, T.J.; Peng, L.; Rawson, P.; Lloyd-Jones, G.; Jordan, T.W. Proteomic phenotyping of Novosphingobium nitrogenifigens reveals a robust capacity for simultaneous nitrogen fixation, polyhydroxyalkanoate production, and resistance to reactive oxygen species. Appl. Environ. Microbiol. 2012, 78, 4802–4815. [Google Scholar] [CrossRef] [Green Version]
- Tang, R.; Peng, X.; Weng, C.; Han, Y. The Overexpression of Phasin and Regulator Genes Promoting the Synthesis of Polyhydroxybutyrate in Cupriavidus necator H16 under Nonstress Conditions. Appl. Environ. Microbiol. 2022, 88, AEM-01458. [Google Scholar] [CrossRef]
- Kutralam-Muniasamy, G.; Peréz-Guevara, F. Genome characteristics dictate poly-R-(3)-hydroxyalkanoate production in Cupriavidus necator H16. World J. Microbiol. Biotechnol. 2018, 34, 79. [Google Scholar] [CrossRef]
- Peoples, O.P.; Sinskey, A.J. Poly-β-hydroxybutyrate biosynthesis in Alcaligenes eutrophus H16. Characterization of the genes encoding β-ketothiolase and acetoacetyl-CoA reductase. J. Biol. Chem. 1989, 264, 15293–15297. [Google Scholar] [CrossRef]
- Winnacker, M. Polyhydroxyalkanoates: Recent Advances in Their Synthesis and Applications. Eur. J. Lipid Sci. Technol. 2019, 121, 1900101. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Lee, S.Y. Factors affecting the economics of polyhydroxyalkanoate production by bacterial fermentation. Appl. Microbiol. Biotechnol. 1999, 51, 13–21. [Google Scholar] [CrossRef]
- Leong, Y.K.; Show, P.L.; Ooi, C.W.; Ling, T.C.; Lan, J.C.W. Current trends in polyhydroxyalkanoates (PHAs) biosynthesis: Insights from the recombinant Escherichia coli. J. Biotechnol. 2014, 180, 52–65. [Google Scholar] [CrossRef]
- Portugal-Nunes, D.J.; Pawar, S.S.; Lidén, G.; Gorwa-Grauslund, M.F. Effect of nitrogen availability on the poly-3-D-hydroxybutyrate accumulation by engineered Saccharomyces cerevisiae. AMB Express 2017, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Poblete-Castro, I.; Escapa, I.F.; Jäger, C.; Puchalka, J.; Chi Lam, C.M.; Schomburg, D.; Prieto, M.A.; Martins dos Santos, V.A. The metabolic response of P. putida KT2442 producing high levels of polyhydroxyalkanoate under single- and multiple-nutrient-limited growth: Highlights from a multi-level omics approach. Microb. Cell Fact. 2012, 11, 34. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Function | Gene Locus (EGO55_) |
|---|---|---|
| PhaA | β-ketothiolase | 09255 |
| 20150 | ||
| 05760 | ||
| 05015 | ||
| 13750 | ||
| 13790 | ||
| 05665 | ||
| 03475 | ||
| 14320 | ||
| PhaB | acetoacetyl-CoA reductase | 18540 |
| 09490 | ||
| PhaC | PHB synthase | 12105 |
| 17580 | ||
| 18530 | ||
| PhaZ | PHB depolymerase | 09590 |
| PhaP | Phasin | 12100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibero, J.; Rivero-Buceta, V.; García, J.L.; Galán, B. Polyhydroxyalkanoate Production by Caenibius tardaugens from Steroidal Endocrine Disruptors. Microorganisms 2022, 10, 706. https://doi.org/10.3390/microorganisms10040706
Ibero J, Rivero-Buceta V, García JL, Galán B. Polyhydroxyalkanoate Production by Caenibius tardaugens from Steroidal Endocrine Disruptors. Microorganisms. 2022; 10(4):706. https://doi.org/10.3390/microorganisms10040706
Chicago/Turabian StyleIbero, Juan, Virginia Rivero-Buceta, José Luis García, and Beatriz Galán. 2022. "Polyhydroxyalkanoate Production by Caenibius tardaugens from Steroidal Endocrine Disruptors" Microorganisms 10, no. 4: 706. https://doi.org/10.3390/microorganisms10040706