
From Free Binding Energy Calculations of SARS-CoV-2—Receptor Interactions to Cellular Immune Responses


Abstract
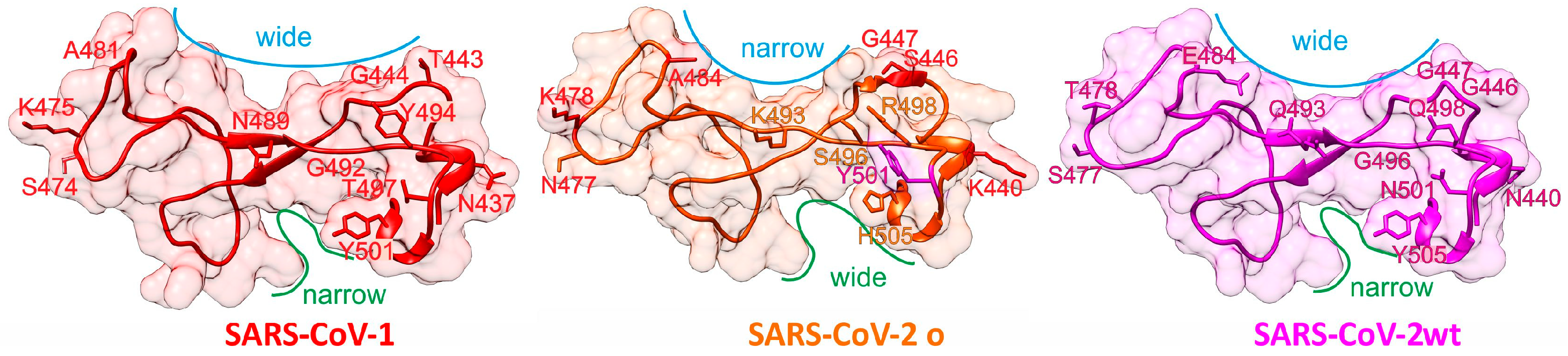
:1. Amino Acid Sequence Alignments Point to a Shift in RBM Characteristics
2. Free Energy Calculations Indicate Weaker Receptor Binding of Omicron’s RBM
3. A Molecular Perspective on Transmissibility and Disease Outcome
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Jhun, H.; Park, H.-Y.; Hisham, Y.; Song, C.-S.; Kim, S. SARS-CoV-2 Delta (B.1.617.2) Variant: A Unique T478K Mutation in Receptor Binding Motif (RBM) of Spike Gene. Immune Netw. 2021, 21, e32. [Google Scholar] [CrossRef] [PubMed]
- ECDC Threat Assessment Brief. Available online: https://www.ecdc.europa.eu/en/publications-data/threatassessment-brief-emergence-sars-cov-2-variant-b.1.1.529 (accessed on 30 November 2021).
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS Coronavirus Spike Receptor-Binding Domain Complexed with Receptor. Science 2005, 390, 1864–1868. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.D.; Dingens, A.S.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C.; et al. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 2020, 182, 1295–1310. [Google Scholar] [CrossRef]
- Guruprasad, L. Human SARS CoV-2 spike protein mutations. Proteins 2020, 89, 569–576. [Google Scholar] [CrossRef]
- Cheng, M.H.; Krieger, J.M.; Kaynak, B.; Arditi, M.; Bahar, I. Impact of South African 501.V2 Variant on SARSCoV-2 Spike Infectivity and Neutralization: A Structure-based Computational Assessment. Bioinformatics, 2021; in press. [Google Scholar] [CrossRef]
- Li, W.; Zhang, C.; Sui, J.; Kuhn, J.H.; Moore, M.J.; Luo, S.; Wong, S.-K.; Huang, I.-C.; Xu, K.; Vasilieva, N.; et al. Receptor and viral determinants of SARS coronavirus adaptation to human ACE2. EMBO J. 2005, 24, 1634–1643. [Google Scholar] [CrossRef] [Green Version]
- Fratev, F. N501Y and K417N Mutations in the Spike Protein of SARS-CoV-2 Alter the Interactions with Both hACE2 and Human-Derived Antibody: A Free Energy of Perturbation Retrospective Study. J. Chem. Inf. Model 2021, 61, 6079–6084. [Google Scholar] [CrossRef]
- Rathnasinghe, R.; Jangra, S.; Cupic, A.; Martínez-Romero, C.; Mulder, L.C.F.; Kehrer, T.; Yildiz, S.; Choi, A.; Mena, I.; De Vrieze, J.; et al. The N501Y mutation in SARS-CoV-2 spike leads to morbidity in obese and aged mice and is neutralized by convalescent and postvaccination human sera. MedRxiv 2021. [Google Scholar] [CrossRef]
- Gu, H.; Chen, Q.; Yang, G.; He, L.; Fan, H.; Deng, Y.-Q.; Wang, Y.; Teng, Y.; Zhao, Z.; Cui, Y.; et al. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science 2020, 369, 1603–1607. [Google Scholar] [CrossRef]
- Opuni, K.F.M.; Solomon, S.; Metzen, F.; Frommholz, D.; Koy, C.; Röwer, C.; Glocker, M.O.; Illges, H.; Anderson, P.C. In silico Epitope Mapping of Glucose-6-Phosphate Isomerase: A Rheumatoid Arthritis Autoantigen. J. Proteom. Bioinform. 2017, 10, 60–72. [Google Scholar] [CrossRef]
- Dehouck, Y.; Kwasigroch, J.M.; Rooman, M.; Gilis, D. BeAtMuSiC: Prediction of changes in protein–protein binding affinity on mutations. Nucleic Acids Res. 2013, 41, W333–W339. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Shi, X.; Jiang, L.; Zhang, S.; Wang, D.; Tong, P.; Guo, D.; Fu, L.; Cui, Y.; Liu, X.; et al. Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res. 2013, 23, 986–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, Z.; Yang, L.; Lian, X.; Xie, Y.; Li, S.; Xin, S.; Cao, P.; Lu, J. The MERS-CoV Receptor DPP4 as a Candidate Binding Target of the SARS-CoV-2 Spike. Science 2020, 23, 101160. [Google Scholar] [CrossRef]
- Kumar, S.; Thambiraja, T.S.; Karuppanan, K.; Subramaniam, G. Omicron and Delta Variant of SARS-CoV-2: A Comparative Computational Study of Spike protein. BioRxiv 2021. [Google Scholar] [CrossRef]
- Wu, L.; Zhou, L.; Mo, M.; Li, Y.; Han, J.; Li, J.; Yang, Y.; Zhang, X.; Gong, C.; Lu, K.; et al. The effect of the multiple mutations in Omicron RBD on its binding to human ACE2 receptor and immune evasion: An investigation of molecular dynamics simulations. Signal Transduct. Target. Ther. 2022, 7. [Google Scholar] [CrossRef]
- Lupala, C.S.; Ye, Y.; Chen, H.; Su, X.-D.; Liu, H. Mutations in RBD of SARS-CoV-2 Omicron variant result stronger binding to human ACE2 protein. Biochem. Biophys. Res. Commun. 2022, 590, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Golcuk, M.; Yildiz, A.; Gur, M. The Omicron Variant Increases the Interactions of SARS-CoV-2 Spike Glycoprotein with ACE2. BioRxiv 2021. [Google Scholar] [CrossRef]
- Shah, M.; Woo, H.G. Omicron: A heavily mutated SARS-CoV-2 variant exhibits stronger binding to ACE2 and potently escape approved COVID-19 therapeutic antibodies. BioRxiv 2021. [Google Scholar] [CrossRef]
- Jawaid, M.Z.; Baidya, A.; Mahboubi-Ardakani, R.; Davis RLCox, D.L. Simulation of the omicron variant of SARS-CoV-2 shows broad antibody escape, weakened ACE2 binding, and modest increase in furin binding. BioRxiv 2021. [Google Scholar] [CrossRef]
- Wu, L.; Zhou, L.; Mo, M.; Liu, T.; Wu, C.; Gong, C.; Lu, K.; Gong, L.; Zhu, W.; Xu, Z. SARS-CoV-2 Omicron RBD shows weaker binding affinity than the currently dominant Delta variant to human ACE2. Signal Transduct. Target. Ther. 2022, 7, 8–9. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Li, L.; Liu, S.; Wang, Q.; Zhang, D.; Xu, Z.; Han, P.; Li, X.; Peng, Q.; Su, C.; et al. Receptor binding and complex structures of human ACE2 to spike RBD from Omicron and Delta SARS-CoV-2. Cell, 2021; in press. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Bertoglio, F.; Steinke, S.; Heine, P.A.; Ynga-Durand, M.A.; Zuo, F.; Du, L.; Korn, J.; Milošević, M.; Wenzel, E.V.; et al. Human serum from SARS-CoV-2 vaccinated and COVID-19 patients shows reduced binding to the RBD of SARS-CoV-2 Omicron variant. MedRxiv 2021. [Google Scholar] [CrossRef]
- Mannar, D.; Saville, J.W.; Zhu, X.; Srivastava, S.S.; Berezuk, A.M.; Tuttle, K.S.; Marquez, C.; Sekirov, I.; Subramaniam, S. SARS-CoV-2 Omicron Variant: ACE2 Binding, Cryo-EM Structure of Spike Protein-ACE2 Complex and Antibody Evasion. BioRxiv 2021. [Google Scholar] [CrossRef]
- Matzinger, P. The Danger Model: A Renewed Sense of Self. Science 2002, 296, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Tsueng, G.; Mullen, J.; Alkuzweny, M.; Cano, M.; Rush, B.; Haag, E.; Latif, A.A.; Zhou, X.; Qian, Z.; Andersen, K.G. Outbreak.info: A standardized, searchable platform to discover and explore COVID-19 resources and data. BioRxiv 2022. [Google Scholar] [CrossRef]
- World of Molecules. Available online: https://www.worldofmolecules.com/3D/B.1.1.529-variant.html (accessed on 30 November 2021).
- Torjesen, I. COVID-19: Omicron may be more transmissible than other variants and partly resistant to existing vaccines, scientists fear. Br. Med. J. 2021, 375, n2943. [Google Scholar] [CrossRef]
- Pulliam, J.R.C.; van Schalkwyk, C.; Govender, N.; von Gottberg, A.; Cohen, C.; Groome, M.J.; Dushoff, J.; Mlisana, K.; Moultrie, H. Increased risk of SARS-CoV-2 reinfection associated with emergence of the Omicron variant in South Africa. Br. Med. J. 2021. [Google Scholar] [CrossRef]
- Willett, B.J.; Grove, J.; MacLean, O.A.; Wilkie, C.; Logan, N.; de Lorenzo, G.; Furnon, W.; Scott, S.; Manali, M.; Szemiel, A.; et al. The hyper-transmissible SARS-CoV-2 Omicron variant exhibits significant antigenic change, vaccine escape and a switch in cell entry mechanism. MedRxiv 2022. [Google Scholar] [CrossRef]
- Olbei, M.; Hautefort, I.; Modos, D.; Treveil, A.; Poletti, M.; Gul, L.; Shannon-Lowe, C.D.; Korcsmaros, T. SARS-CoV-2 Causes a Different Cytokine Response Compared to Other Cytokine Storm-Causing Respiratory Viruses in Severely Ill Patients. Front. Immunol. 2021, 12, 629193. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, F.; Torsiello, E.; Spiezia, F.; Oliva, F.; Tingart, M.; Maffulli, N. Association between HLA genotypes and COVID-19 susceptibility, severity and progression: A comprehensive review of the literature. Eur. J. Med. Res. 2021, 26, 84. [Google Scholar] [CrossRef]
- Naemi, F.M.A.; Al-Adwani, S.; Al-Khatabi, H.; Al-Nazawi, A. Association between the HLA genotype and the severity of COVID-19 infection among South Asians. J. Med. Virol. 2021, 93, 4430–4437. [Google Scholar] [CrossRef] [PubMed]
- Abdool Karim, S.S.; Abdool Karim, Q. Omicron SARS-CoV-2 variant: A new chapter in the COVID-19 pandemic. Lancet 2021, 398, 2126–2128. [Google Scholar] [CrossRef]
- Wolter, N.; Jassat, W.; Walaza, S. Early assessment of the clinical severity of the SARS-CoV-2 Omicron variant in South Africa. MedRxiv 2021. [Google Scholar] [CrossRef]
- Ferguson, N.; Ghani, A.; Hinsley, W.; Volz, E.; on behalf of the Imperial College COVID-19 Response Team. Report 50: Hospitalisation risk for Omicron cases in England 2021; Imperial College London: London, UK, 2021; Volume 2021, pp. 1–12. [Google Scholar] [CrossRef]
- Sheikh, A.; Kerr, S.; Woolhouse, M.; McMenamin, J.; Robertson, C. Severity of Omicron Variant of Concern and Vaccine Effectiveness Against Symptomatic Disease: National Cohort with Nested Test Negative Design Study in Scotland. Available online: https://www.pure.ed.ac.uk/ws/portalfiles/portal/245818096/Severity_of_Omicron_variant_of_concern_and_vaccine_effectiveness_against_symptomatic_disease.pdf (accessed on 23 December 2021).
- Lewnard, J.A.; Hong, V.X.; Patel, M.M.; Kahn, R.; Lipsitch, M.; Tartof, S.Y. Clinical outcomes among patients infected with Omicron (B.1.1.529) SARS-CoV-2 variant in southern California. MedRxiv 2022. [Google Scholar] [CrossRef]
- Wang, L.; Berger, N.A.; Kaelber, D.C.; Davis, P.B.; Volkow, N.D.; Xu, R. Comparison of outcomes from COVID infection in pediatric and adult patients before and after the emergence of Omicron. MedRxiv 2022. [Google Scholar] [CrossRef]
- Chan Chi-wai, M. Available online: https://www.med.hku.hk/en/news/press/20211215-omicron-sars-cov-2-infection?utm_medium=social&utm_source=twitter&utm_campaign=press_release (accessed on 23 December 2021).
- Peacock, T.P.; Brown, J.C.; Zhou, J.; Thakur, N.; Newman, J.; Kugathasan, R.; Sukhova, K.; Kaforou, M.; Bailey, D.; Barclay, W.S. The SARS-CoV-2 variant, Omicron, shows enhanced replication in human primary nasal epithelial cells. BioRxiv 2021. [Google Scholar] [CrossRef]
- Telenti, A.; Arvin, A.; Corey, L. After the pandemic: Perspectives on the future trajectory of COVID-19. Nature 2021, 596, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.A.; McBride, S.K.; Winders, B.; Schoen, D.; Trautmann, L.; Curlin, M.E.; Tafesse, F.G. Antibody Response and Variant Cross-Neutralization After SARS-CoV-2 Breakthrough Infection. JAMA 2022, 327, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Karim, F.; Cele, S.; San, J.E.; Lustig, G.; Tegally, H.; Bernstein, M.; Ganga, Y.; Jule, Z.; Reedoy, K.; et al. Omicron infection enhances neutralizing immunity against the Delta variant. Medrxiv 2021. [Google Scholar] [CrossRef]


{kind=link}
| Virus | Receptor Binding Motif/Amino Acid Sequence (a,b,c,d) |
|---|---|
| SARS-2 wt | 437NSNNLDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPY508 |
| SARS-2 α | 437NSNNLDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQAGSTPCNGVKGFNCYFPLQPYGFQPTYGVGYQPY508 |
| SARS-2 δ | 437NSNNLDSKVGGNYNYRYRLFRKSNLKPFERDISTEIYQAGSKPCNGVEGFNCYFPLQSYGFQPTNGVGYQPY508 |
| SARS-2 ο | 437NSNKLDSKVSGNYNYLYRLFRKSNLKPFERDISTEIYQAGNKPCNGVAGFNCYFPLKSYSFRPTYGVGHQPY508 |
| SARS-1 wt | 434NTRNIDATSTGNYNYKYRYLRHGKLRPFERDISNVPFSPDGKPCTPPA-LNCYWPLNDYGFYTTTGIGYQPY504 |
| BM48-31 | NTNSLDS--SNEFEYR-R-FRHGKIKPYGRDLSNVLFNPSGGTC-SAEGLNCYKPLASYGFTQSSGIGFQPY |
| Rp3 | NTAKQDQG-----QYYYRSHRKTKLKPFERDLSSDE-NGV-RT-LS-----------TYDFYPSVPVAYQAT |
| Amino Acid Residue Exchange | Variant of Concern and Human Binding Partner/Receptor Complex | |||||||
|---|---|---|---|---|---|---|---|---|
| Alpha (b,c) | Delta (b,c) | Omicron (b,c) | ||||||
| hACE2 | hDPP-IV | hACE2 | hDPP-IV | hACE2 | hDPP-IV | |||
| N440K | n.a. | n.a. | n.b. | n.a. | n.b. | n.b. | ||
| G446S | n.a. | n.a. | n.a. | n.b. | +1.05 | n.b. | ||
| L452R | n.a. | n.a. | n.b. | n.b. | n.a. | n.a. | ||
| S477N | n.a. | n.a. | n.a. | n.a. | n.b. | +0.39 | ||
| T478K | n.a. | n.a. | n.b. | +1.00 | n.b. | +1.00 | ||
| E484K/A (d) | +0.07 | −0.05 | n.a. | n.a. | +0.10 | −0.20 | ||
| Q493K (e) | n.a. | n.a. | n.a. | n.a. | +0.97 | +0.83 | ||
| S494P | n.b. | n.b. | n.a. | n.a. | n.a. | n.a. | ||
| G496S | n.a. | n.a. | n.a. | n.a. | +0.44 | n.b. | ||
| Q498R | n.a. | n.a. | n.a. | n.a. | +0.84 | n.b. | ||
| N501Y | −0.08 | +0.19 | n.a. | n.a. | −0.08 | +0.19 | ||
| Y505H | n.a. | n.a. | n.a. | n.a. | +1.09 | +1.20 | ||
| sum | −0.01 | +0.14 | ±0.00 | +1.00 | +4.41 | +3.41 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glocker, M.O.; Opuni, K.F.M.; Thiesen, H.-J. From Free Binding Energy Calculations of SARS-CoV-2—Receptor Interactions to Cellular Immune Responses. Medicina 2022, 58, 226. https://doi.org/10.3390/medicina58020226
Glocker MO, Opuni KFM, Thiesen H-J. From Free Binding Energy Calculations of SARS-CoV-2—Receptor Interactions to Cellular Immune Responses. Medicina. 2022; 58(2):226. https://doi.org/10.3390/medicina58020226
Chicago/Turabian StyleGlocker, Michael O., Kwabena F. M. Opuni, and Hans-Juergen Thiesen. 2022. "From Free Binding Energy Calculations of SARS-CoV-2—Receptor Interactions to Cellular Immune Responses" Medicina 58, no. 2: 226. https://doi.org/10.3390/medicina58020226
APA StyleGlocker, M. O., Opuni, K. F. M., & Thiesen, H.-J. (2022). From Free Binding Energy Calculations of SARS-CoV-2—Receptor Interactions to Cellular Immune Responses. Medicina, 58(2), 226. https://doi.org/10.3390/medicina58020226