
Eribulin Mesylate Improves Cisplatin-Induced Cytotoxicity of Triple-Negative Breast Cancer by Extracellular Signal-Regulated Kinase 1/2 Activation

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Lines and Culture
2.3. Cell Viability Assay
2.4. Combination Index
2.5. Caspase-3/CPP32 Colorimetric Assay
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
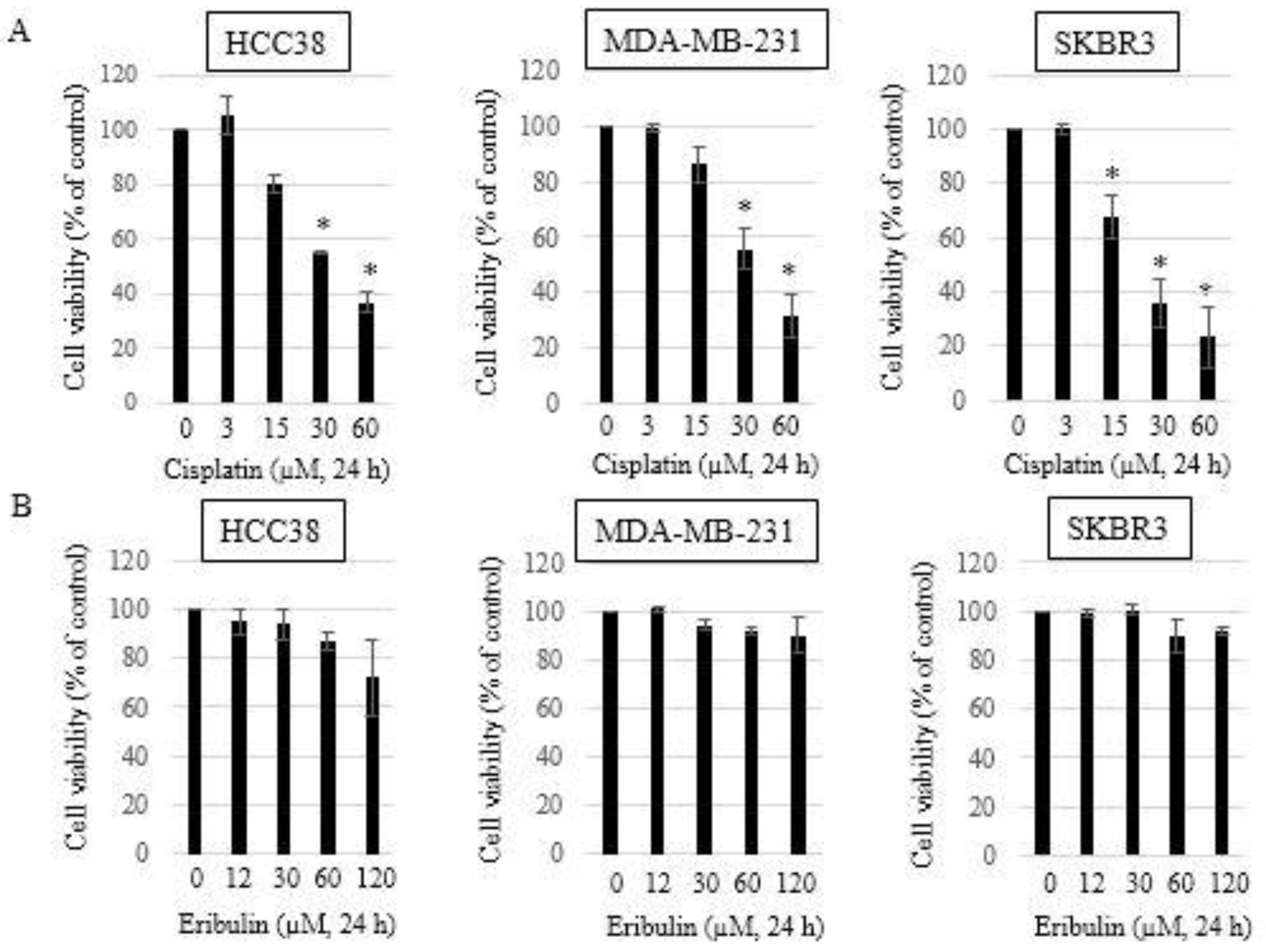
3.1. Growth Inhibitory Effects of Single-Agent Eribulin or Cisplatin in HCC38, MDA-MB-231, and SKBR3 Breast Cancer Cell Lines
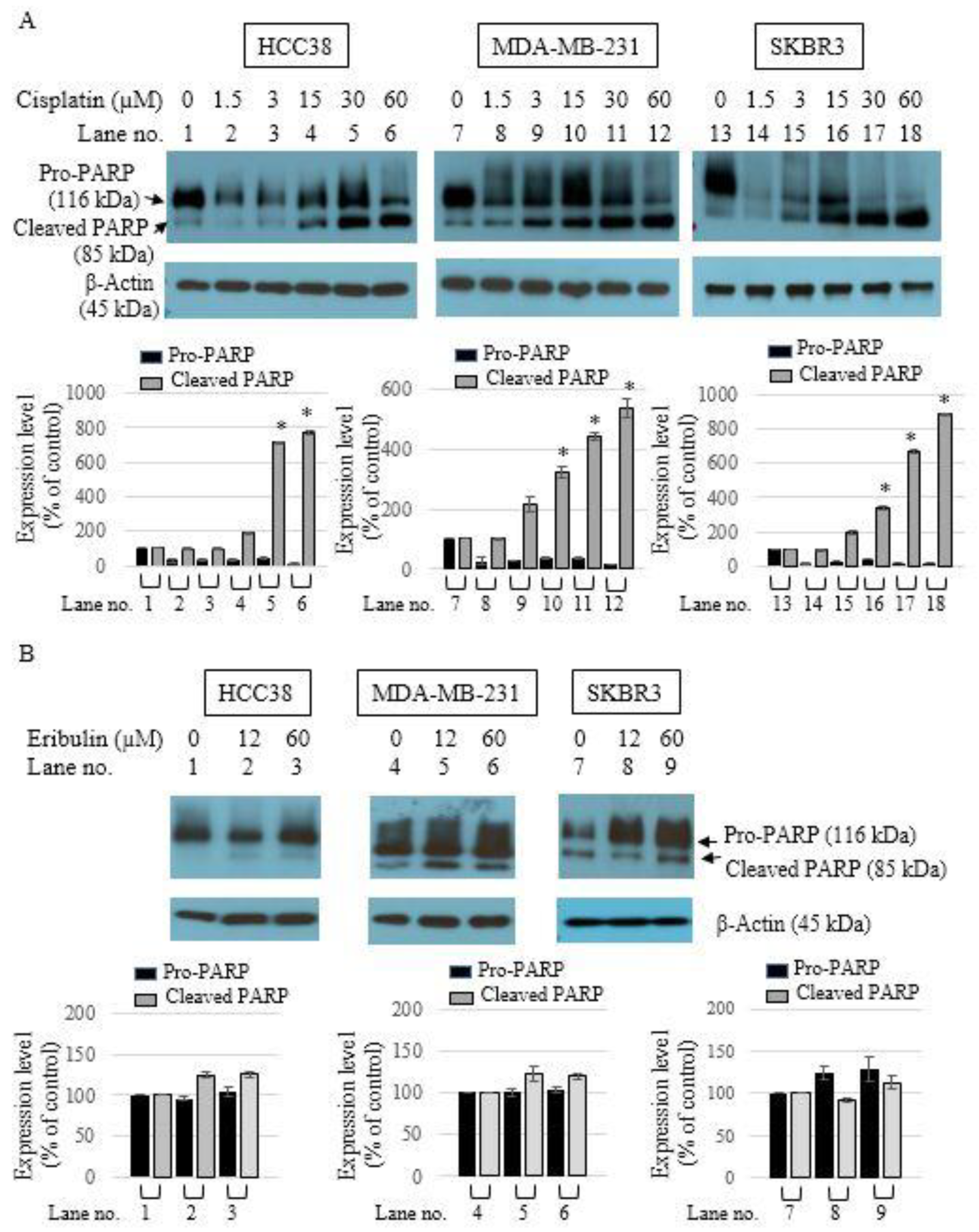
3.2. Apoptotic Activity of Single-Agent Eribulin or Cisplatin in HCC38, MDA-MB-231, and SKBR3 Breast Cancer Cell Lines
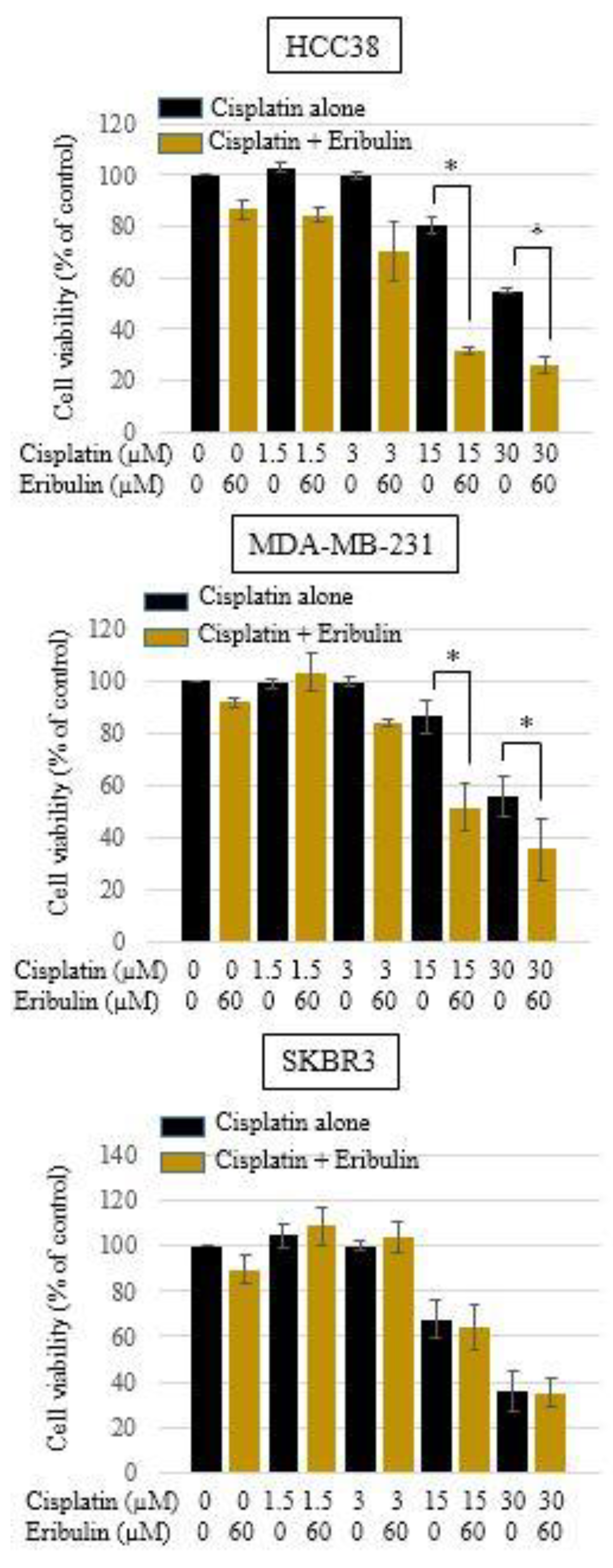
3.3. Effect of Cisplatin and Eribulin Combination on HCC38, MDA-MB-231, and SKBR3 Cells
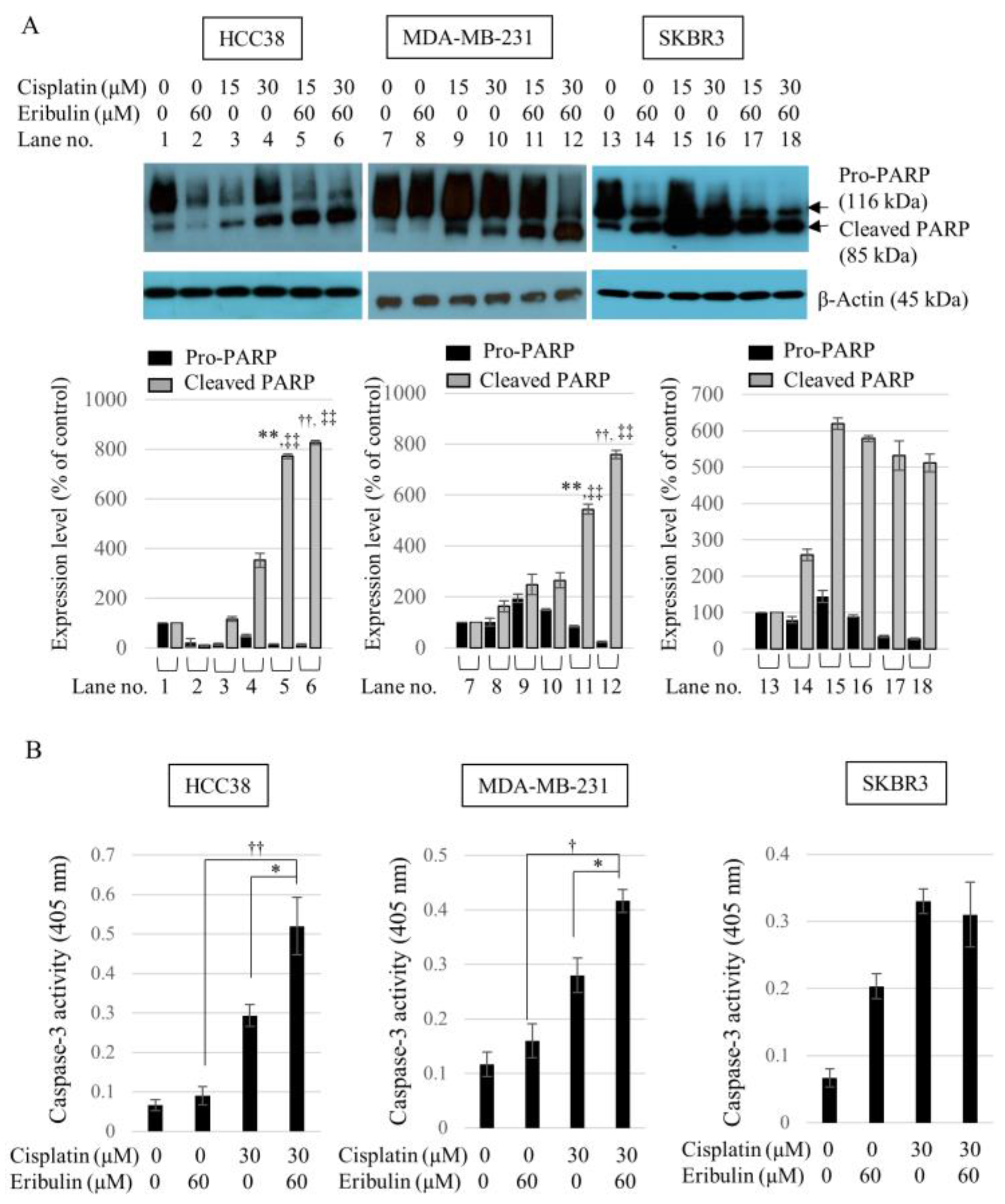
3.4. Combination of Cisplatin and Eribulin Significantly Enhances the Apoptosis of HCC38 and MDA-MB-231 TNBC Cells
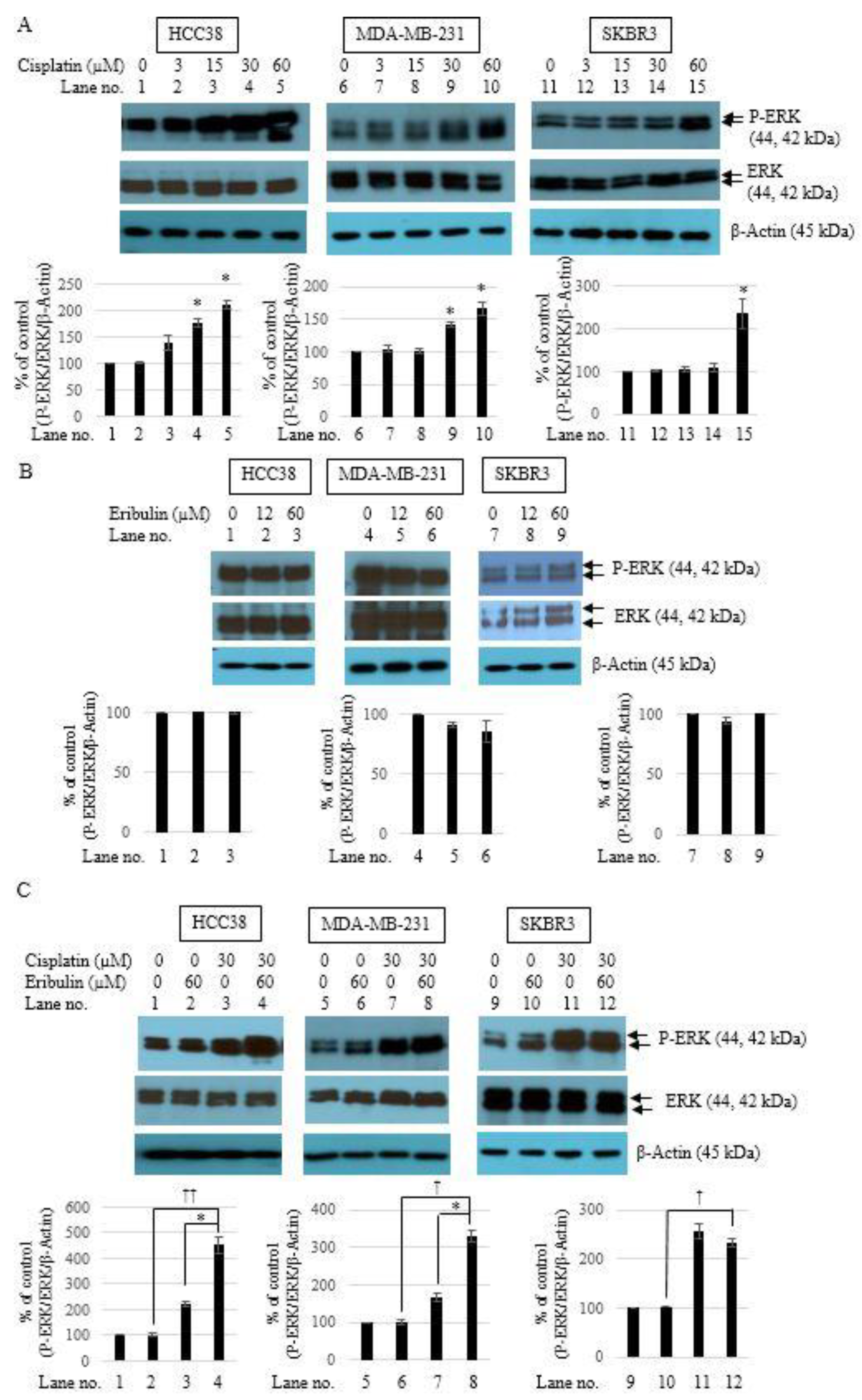
3.5. Combination of Cisplatin and Eribulin Synergistically Activates ERK1/2 in TNBC Cells
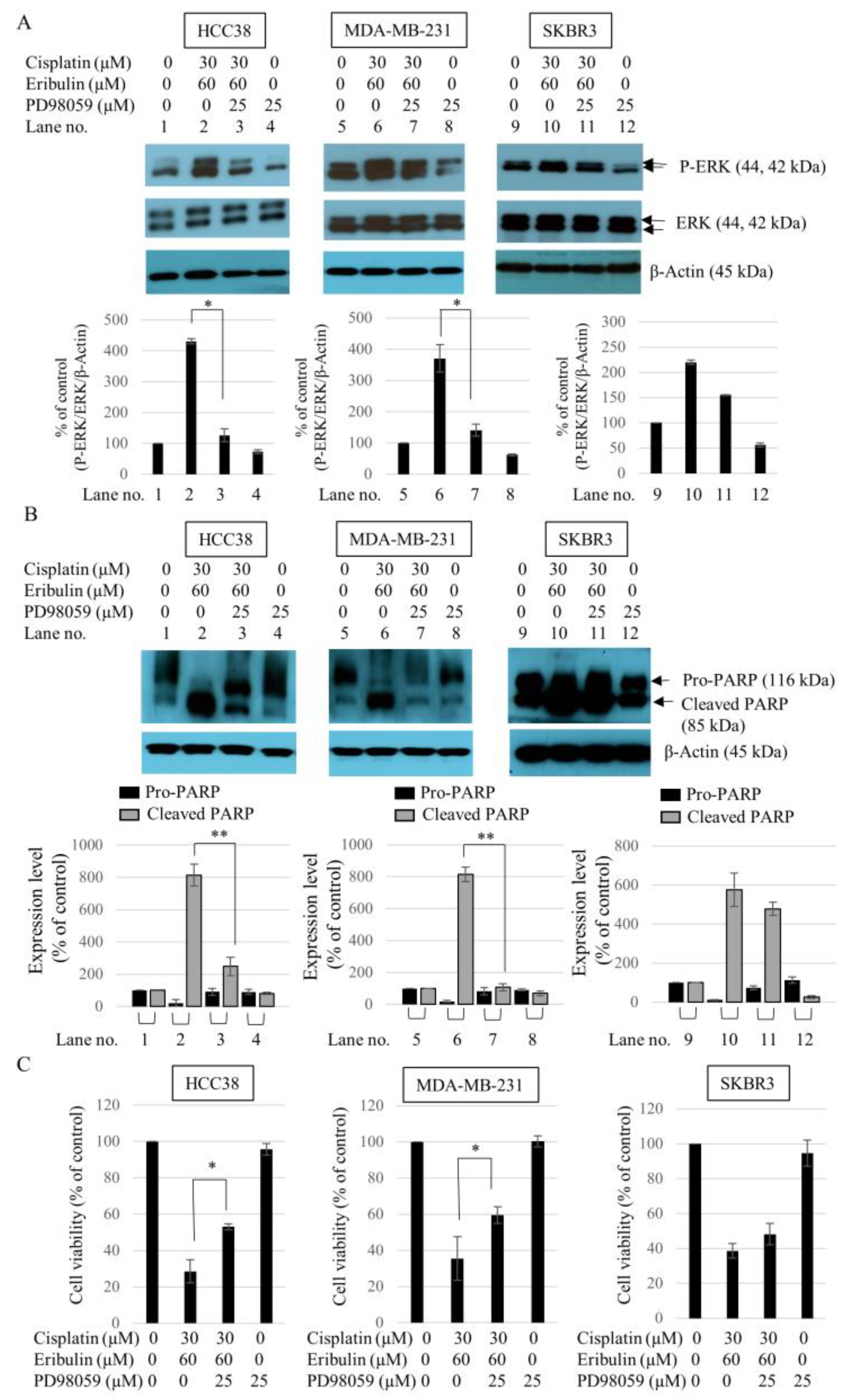
3.6. Effect of ERK1/2 Inhibitor PD98059 on the Sensitivity of TNBC to the Cisplatin and Eribulin Combination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haffty, B.G.; Yang, Q.; Reiss, M.; Kearney, T.; Higgins, S.A.; Weidhaas, J.; Harris, L.; Hait, W.; Toppmeyer, D. Locoregional relapse and distant metastasis in conservatively managed triple negative early-stage breast cancer. J. Clin. Oncol. 2006, 24, 5652–5657. [Google Scholar] [CrossRef] [PubMed]
- Tischkowitz, M.; Brunet, J.S.; Bégin, L.R.; Huntsman, D.G.; Cheang, M.C.; Akslen, L.A.; Nielsen, T.O.; Foulkes, W.D. Use of immunohistochemical markers can refine prognosis in triple negative breast cancer. BMC Cancer 2007, 7, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, K.R.; Brown, M.; Cress, R.D.; Parise, C.A.; Caggiano, V. Descriptive analysis of estrogen receptor (ER)-negative, progesterone receptor (PR)-negative, and HER2-negative invasive breast cancer, the so-called triple-negative phenotype: A population-based study from the California cancer Registry. Cancer 2007, 109, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.N.; Broadwater, G.; Lin, N.U.; Miron, A.; Schnitt, S.J.; Cowan, D.; Lara, J.; Bleiweiss, I.; Berry, D.; Ellis, M.; et al. Molecular subtypes of breast cancer in relation to paclitaxel response and outcomes in women with metastatic disease: Results from CALGB 9342. Breast Cancer Res. BCR 2006, 8, R66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, G.J.; Naidu, S.; Topham, A.K.; Guiles, F.; Xu, Y.; McCue, P.; Schwartz, G.F.; Park, P.K.; Rosenberg, A.L.; Brill, K.; et al. Differences in breast carcinoma characteristics in newly diagnosed African-American and Caucasian patients: A single-institution compilation compared with the National Cancer Institute’s Surveillance, Epidemiology, and End Results database. Cancer 2007, 110, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Lakhani, S.R.; Van De Vijver, M.J.; Jacquemier, J.; Anderson, T.J.; Osin, P.P.; McGuffog, L.; Easton, D.F. The pathology of familial breast cancer: Predictive value of immunohistochemical markers estrogen receptor, progesterone receptor, HER-2, and p53 in patients with mutations in BRCA1 and BRCA2. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 2310–2318. [Google Scholar] [CrossRef] [Green Version]
- Foulkes, W.D.; Stefansson, I.M.; Chappuis, P.O.; Bégin, L.R.; Goffin, J.R.; Wong, N.; Trudel, M.; Akslen, L.A. Germline BRCA1 mutations and a basal epithelial phenotype in breast cancer. J. Natl. Cancer Inst. 2003, 95, 1482–1485. [Google Scholar] [CrossRef]
- Carey, L.A.; Dees, E.C.; Sawyer, L.; Gatti, L.; Moore, D.T.; Collichio, F.; Ollila, D.W.; Sartor, C.I.; Graham, M.L.; Perou, C.M. The triple negative paradox: Primary tumor chemosensitivity of breast cancer subtypes. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 2329–2334. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P. Biology and Management of Patients with Triple-Negative Breast Cancer. Oncologist 2016, 21, 1050–1062. [Google Scholar] [CrossRef] [Green Version]
- Mani, S.; Swami, U. Eribulin mesilate, a halichondrin B analogue, in the treatment of breast cancer. Drugs Today 2010, 46, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Twelves, C.; Cortes, J.; Vahdat, L.; Olivo, M.; He, Y.; Kaufman, P.A.; Awada, A. Efficacy of eribulin in women with metastatic breast cancer: A pooled analysis of two phase 3 studies. Breast Cancer Res. Treat. 2014, 148, 553–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S. Cisplatin: The first metal based anticancer drug. Bioorg. Chem. 2019, 88, 102925. [Google Scholar] [CrossRef]
- Chen, S.-H.; Chang, J.-Y. New Insights into Mechanisms of Cisplatin Resistance: From Tumor Cell to Microenvironment. Int. J. Mol. Sci. 2019, 20, 4136. [Google Scholar] [CrossRef] [Green Version]
- Jovanović, B.; Mayer, I.A.; Mayer, E.L.; Abramson, V.G.; Bardia, A.; Sanders, M.E.; Kuba, M.G.; Estrada, M.V.; Beeler, J.S.; Shaver, T.M.; et al. A Randomized Phase II Neoadjuvant Study of Cisplatin, Paclitaxel with or without Everolimus in Patients with Stage II/III Triple-Negative Breast Cancer (TNBC): Responses and Long-term Outcome Correlated with Increased Frequency of DNA Damage Response Gene Mutations, TNBC Subtype, AR Status, and Ki67. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 4035–4045. [Google Scholar] [CrossRef] [Green Version]
- Sirohi, B.; Arnedos, M.; Popat, S.; Ashley, S.; Nerurkar, A.; Walsh, G.; Johnston, S.; Smith, I.E. Platinum-based chemotherapy in triple-negative breast cancer. Ann. Oncol. 2008, 19, 1847–1852. [Google Scholar] [CrossRef]
- Silver, D.P.; Richardson, A.L.; Eklund, A.C.; Wang, Z.C.; Szallasi, Z.; Li, Q.; Juul, N.; Leong, C.O.; Calogrias, D.; Buraimoh, A.; et al. Efficacy of neoadjuvant Cisplatin in triple-negative breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2010, 28, 1145–1153. [Google Scholar] [CrossRef]
- Bhattacharyya, G.; Basu, S.; Agarwal, V.; Malhotra, H.; Pareekh, P.; Babu, K.; Aggarwala, D. 41LBA Single institute phase II study of weekly cisplatinum and metronomic dosing of endoxan and methotrexate in second line metastatic breast cancer triple-negative. Ejc Suppl. 2009, 7, 18–19. [Google Scholar] [CrossRef]
- Hahnen, E.; Lederer, B.; Hauke, J.; Loibl, S.; Kröber, S.; Schneeweiss, A.; Denkert, C.; Fasching, P.A.; Blohmer, J.U.; Jackisch, C.; et al. Germline Mutation Status, Pathological Complete Response, and Disease-Free Survival in Triple-Negative Breast Cancer: Secondary Analysis of the GeparSixto Randomized Clinical Trial. JAMA Oncol. 2017, 3, 1378–1385. [Google Scholar] [CrossRef]
- Poggio, F.; Bruzzone, M.; Ceppi, M.; Pondé, N.F.; La Valle, G.; Del Mastro, L.; de Azambuja, E.; Lambertini, M. Platinum-based neoadjuvant chemotherapy in triple-negative breast cancer: A systematic review and meta-analysis. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2018, 29, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- Peshkin, B.N.; Alabek, M.L.; Isaacs, C. BRCA1/2 mutations and triple negative breast cancers. Breast Dis 2010, 32, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, A.K.; Nandy, M.; Sharma, G. Cisplatin as neoadjuvant chemotherapy in triple negative breast cancer: Exciting early results. Indian J. Med. Paediatr. Oncol. 2010, 31, 76–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.J.; Mann, E.; Wright, G.; Piett, C.G.; Nagel, Z.D.; Gassman, N.R. Exploiting DNA repair defects in triple negative breast cancer to improve cell killing. Ther. Adv. Med. Oncol. 2020, 12, 1758835920958354. [Google Scholar] [CrossRef] [PubMed]
- Koczywas, M.; Frankel, P.H.; Synold, T.W.; Lenz, H.J.; Mortimer, J.E.; El-Khoueiry, A.B.; Gandara, D.R.; Cristea, M.C.; Chung, V.M.; Lim, D.; et al. Phase I study of the halichondrin B analogue eribulin mesylate in combination with cisplatin in advanced solid tumors. Br. J. Cancer 2014, 111, 2268–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, M.; Matsui, J.; Towle, M.J.; Wu, J.; McGonigle, S.; MH, D.E.B.; Uenaka, T.; Nomoto, K.; Littlefield, B.A. Broad-spectrum Preclinical Antitumor Activity of Eribulin (Halaven®): Combination with Anticancer Agents of Differing Mechanisms. Anticancer Res. 2018, 38, 3375–3385. [Google Scholar] [CrossRef] [PubMed]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-activated protein (MAP) kinase pathways: Regulation and physiological functions. Endocr. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Oba, T.; Ito, K.I. Combination of two anti-tubulin agents, eribulin and paclitaxel, enhances anti-tumor effects on triple-negative breast cancer through mesenchymal-epithelial transition. Oncotarget 2018, 9, 22986–23002. [Google Scholar] [CrossRef] [Green Version]
- Chalakur-Ramireddy, N.K.R.; Pakala, S.B. Combined drug therapeutic strategies for the effective treatment of Triple Negative Breast Cancer. Biosci. Rep. 2018, 38, BSR20171357. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.O.; Kang, M.J.; Byun, W.S.; Kim, S.A.; Seo, I.H.; Han, J.A.; Moon, J.W.; Kim, J.H.; Kim, S.J.; Lee, E.J.; et al. Metformin overcomes resistance to cisplatin in triple-negative breast cancer (TNBC) cells by targeting RAD51. Breast Cancer Res. BCR 2019, 21, 115. [Google Scholar] [CrossRef] [Green Version]
- Gohr, K.; Hamacher, A.; Engelke, L.H.; Kassack, M.U. Inhibition of PI3K/Akt/mTOR overcomes cisplatin resistance in the triple negative breast cancer cell line HCC38. BMC Cancer 2017, 17, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.L.; Wen, X.M.; Lai, Q.H.; Li, J.; Wang, X.W. Lenalidomide improvement of cisplatin antitumor efficacy on triple-negative breast cancer cells in vitro. Oncol. Lett. 2018, 15, 6469–6474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czarnomysy, R.; Surażyński, A.; Popławska, B.; Rysiak, E.; Pawłowska, N.; Czajkowska, A.; Bielawski, K.; Bielawska, A. Synergistic action of cisplatin and echistatin in MDA-MB-231 breast cancer cells. Mol. Cell. Biochem. 2017, 427, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Tsai, T.F.; Lin, J.F.; Lin, Y.C.; Chou, K.Y.; Chen, H.E.; Ho, C.Y.; Chen, P.C.; Hwang, T.I. Cisplatin contributes to programmed death-ligand 1 expression in bladder cancer through ERK1/2-AP-1 signaling pathway. Biosci. Rep. 2019, 39, BSR20190362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Hou, X.; Wang, X.; Shi, Y.; Xu, L.; Zheng, X.; Liu, N.; Qiu, A.; Zhuang, S. Correction: 3-deazaneplanocin A protects against cisplatin-induced renal tubular cell apoptosis and acute kidney injury by restoration of E-cadherin expression. Cell Death Dis. 2019, 10, 543. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.K.; Choi, C.H.; Oh, H.L.; Kim, Y.K. Role of ERK activation in cisplatin-induced apoptosis in A172 human glioma cells. Neurotoxicology 2004, 25, 915–924. [Google Scholar] [CrossRef]
- Cagnol, S.; Chambard, J.C. ERK and cell death: Mechanisms of ERK-induced cell death--apoptosis, autophagy and senescence. FEBS J. 2010, 277, 2–21. [Google Scholar] [CrossRef]
- Cui, W.; Yazlovitskaya, E.M.; Mayo, M.S.; Pelling, J.C.; Persons, D.L. Cisplatin-induced response of c-jun N-terminal kinase 1 and extracellular signal--regulated protein kinases 1 and 2 in a series of cisplatin-resistant ovarian carcinoma cell lines. Mol. Carcinog. 2000, 29, 219–228. [Google Scholar] [CrossRef]
- Liu, Y.; Bi, T.; Wang, G.; Dai, W.; Wu, G.; Qian, L.; Gao, Q.; Shen, G. Lupeol inhibits proliferation and induces apoptosis of human pancreatic cancer PCNA-1 cells through AKT/ERK pathways. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 295–304. [Google Scholar] [CrossRef]
- Song, H.; Wei, M.; Liu, W.; Shen, S.; Li, J.; Wang, L. Cisplatin induced apoptosis of ovarian cancer A2780s cells by activation of ERK/p53/PUMA signals. Histol. Histopathol. 2018, 33, 73–79. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Cispatin (µM) | Eribulin (µM) | CI | DRI (Cisplatin) | DRI (Eribulin) |
|---|---|---|---|---|---|
| HCC38 | 3 | 12 | 0.45 | 13.91 | 2.62 |
| 15 | 30 | 0.40 | 8.05 | 3.56 | |
| 30 | 60 | 0.49 | 4.06 | 24.65 | |
| 60 | 120 | 0.45 | 2.37 | 34.13 | |
| MDA-MB-231 | 3 | 12 | 8.15 | 1.0 | 0.14 |
| 15 | 30 | 0.58 | 4.8 | 2.67 | |
| 30 | 60 | 0.53 | 3.0 | 12.19 | |
| 60 | 120 | 0.44 | 2.5 | 23.29 | |
| SKBR3 | 3 | 12 | 4.68 | ND | ND |
| 15 | 30 | 5.56 | ND | ND | |
| 30 | 60 | 1.27 | ND | ND | |
| 60 | 120 | 1.07 | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, H.; Lee, M.; Cha, E.; Sul, J.; Park, J.; Lee, J. Eribulin Mesylate Improves Cisplatin-Induced Cytotoxicity of Triple-Negative Breast Cancer by Extracellular Signal-Regulated Kinase 1/2 Activation. Medicina 2022, 58, 547. https://doi.org/10.3390/medicina58040547
Ko H, Lee M, Cha E, Sul J, Park J, Lee J. Eribulin Mesylate Improves Cisplatin-Induced Cytotoxicity of Triple-Negative Breast Cancer by Extracellular Signal-Regulated Kinase 1/2 Activation. Medicina. 2022; 58(4):547. https://doi.org/10.3390/medicina58040547
Chicago/Turabian StyleKo, Hyemi, Myungsun Lee, Eunyoung Cha, Jiyoung Sul, Junbeom Park, and Jinsun Lee. 2022. "Eribulin Mesylate Improves Cisplatin-Induced Cytotoxicity of Triple-Negative Breast Cancer by Extracellular Signal-Regulated Kinase 1/2 Activation" Medicina 58, no. 4: 547. https://doi.org/10.3390/medicina58040547
APA StyleKo, H., Lee, M., Cha, E., Sul, J., Park, J., & Lee, J. (2022). Eribulin Mesylate Improves Cisplatin-Induced Cytotoxicity of Triple-Negative Breast Cancer by Extracellular Signal-Regulated Kinase 1/2 Activation. Medicina, 58(4), 547. https://doi.org/10.3390/medicina58040547