
Microbiota-Accessible Boron-Containing Compounds in Complex Regional Pain Syndrome
 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Dysbiosis and Degradation of the Mucus Gel Layer in Complex Regional Pain Syndrome
3. Boron: Essential Element in Host–Microbiota Symbiosis
4. Autoinducer-2–Borate Deficiency and Dysbiosis
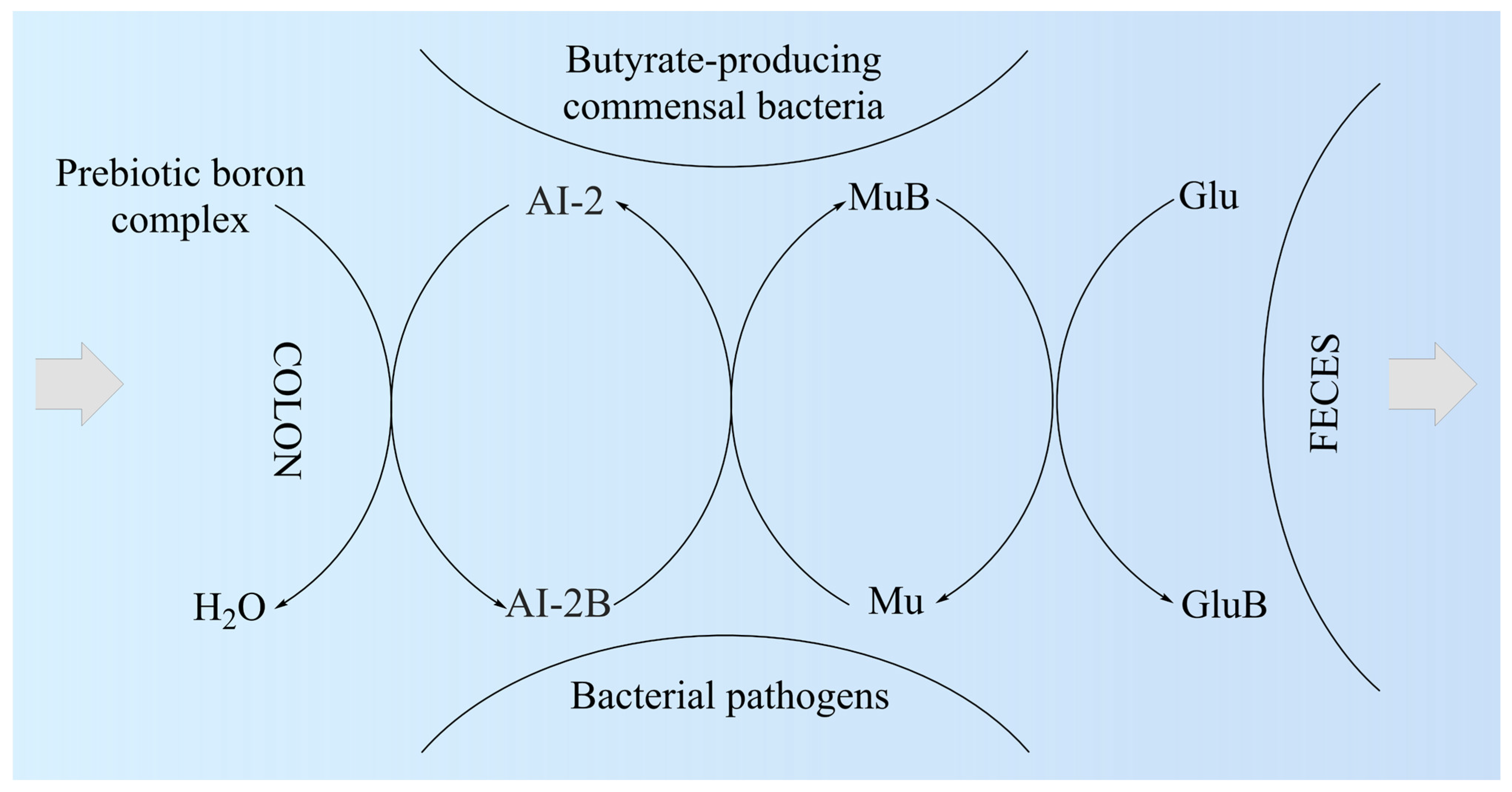
5. Perspectives to Use the Prebiotic Boron-Containing Compounds in Complex Regional Pain Syndrome
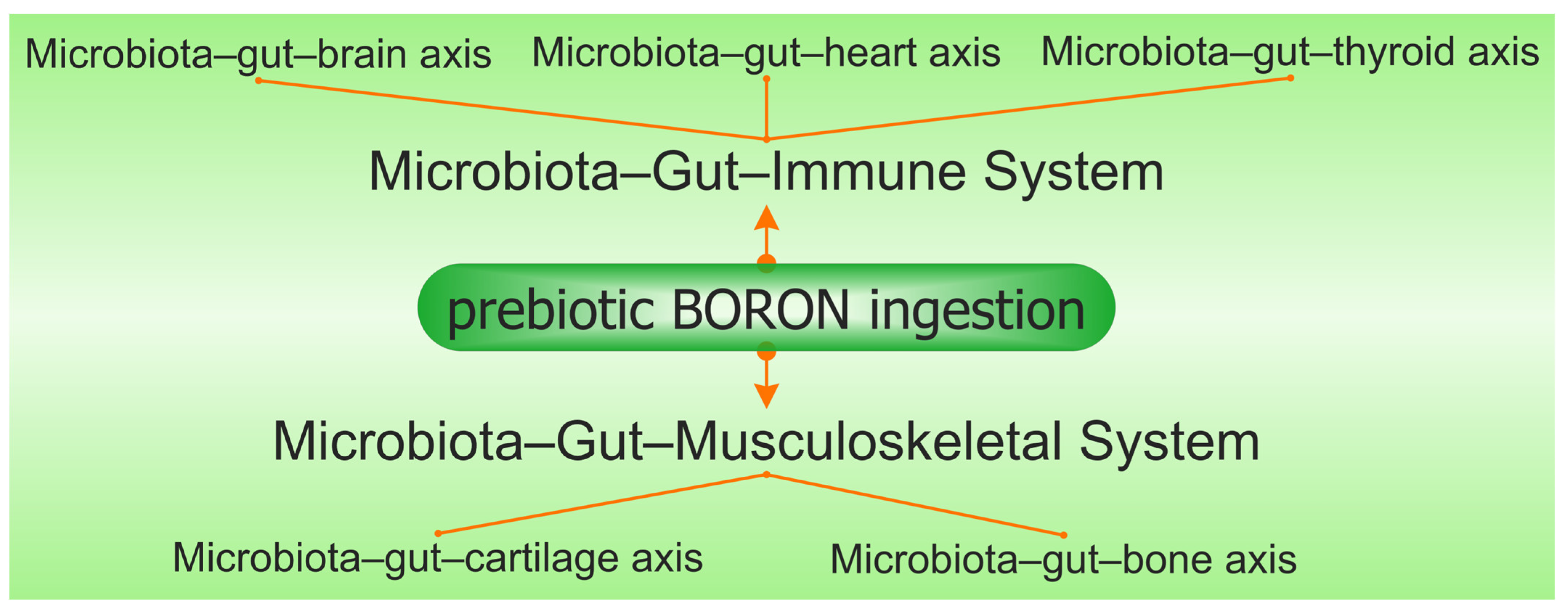
6. Microbiota–Gut–Immune System
7. Microbiota–Gut–Musculoskeletal System
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taylor, S.S.; Noor, N.; Urits, I.; Paladini, A.; Sadhu, M.S.; Gibb, C.; Carlson, T.; Myrcik, D.; Varrassi, G.; Viswanath, O. Complex regional pain syndrome: A comprehensive review. Pain Ther. 2021, 10, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Crock, L.W.; Baldridge, M.T. A role for the microbiota in complex regional pain syndrome? Neurobiol. Pain 2020, 8, 100054. [Google Scholar] [CrossRef]
- Birklein, F.; Ajit, S.K.; Goebel, A.; Perez, R.S.G.M.; Sommer, C. Complex regional pain syndrome—Phenotypic characteristics and potential biomarkers. Nat. Rev. Neurol. 2018, 14, 272–284. [Google Scholar] [CrossRef]
- Birklein, F.; Dimova, V. Complex regional pain syndrome—Up-to-date. Pain Rep. 2017, 2, e624. [Google Scholar] [CrossRef] [PubMed]
- Mekhail, N.; Deer, T.R.; Poree, L.; Staats, P.S.; Burton, A.W.; Connolly, A.T.; Karst, E.; Mehanny, D.S.; Saweris, Y.; Levy, R.M. Cost-effectiveness of dorsal root ganglion stimulation or spinal cord stimulation for complex regional pain syndrome. Neuromodulation 2021, 24, 708–718. [Google Scholar] [CrossRef]
- Li, W.W.; Guo, T.Z.; Shi, X.; Birklein, F.; Schlereth, T.; Kingery, W.S.; Clark, J.D. Neuropeptide regulation of adaptive immunity in the tibia fracture model of complex regional pain syndrome. J. Neuroinflamm. 2018, 15, 105. [Google Scholar] [CrossRef] [PubMed]
- Dirckx, M.; Stronks, D.L.; van Bodegraven-Hof, E.A.M.; Wesseldijk, F.; Groeneweg, J.G.; Huygen, F.J.P.M. Inflammation in cold complex regional pain syndrome. Acta Anaesthesiol. Scand. 2015, 59, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.; Leinders, M.; Üçeyler, N. Inflammation in the pathophysiology of neuropathic pain. Pain 2018, 159, 595–602. [Google Scholar] [CrossRef]
- Baronio, M.; Sadia, H.; Paolacci, S.; Prestamburgo, D.; Miotti, D.; Guardamagna, V.A.; Natalini, G.; Sullivan, S.G.B.; Bertelli, M. Molecular aspects of regional pain syndrome. Pain Res. Manag. 2020, 2020, 7697214. [Google Scholar] [CrossRef]
- Wehmeyer, C.; Pap, T.; Buckley, C.D.; Naylor, A.J. The role of stromal cells in inflammatory bone loss. Clin. Exp. Immunol. 2017, 189, 1–11. [Google Scholar] [CrossRef]
- Littlejohn, G. Neurogenic neuroinflammation in fibromyalgia and complex regional pain syndrome. Nat. Rev. Rheumatol. 2015, 11, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Tonelli Enrico, V.; Vo, N.; Methe, B.; Morris, A.; Sowa, G. An unexpected connection: A narrative review of the associations between gut microbiome and musculoskeletal pain. Eur. Spine J. 2022, 31, 3603–3615. [Google Scholar] [CrossRef] [PubMed]
- Dworsky-Fried, Z.; Kerr, B.J.; Taylor, A.M.W. Microbes, microglia, and pain. Neurobiol. Pain 2020, 7, 100045. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.Z.; Wei, T.; Tajerian, M.; Clark, J.D.; Birklein, F.; Goebel, A.; Li, W.W.; Sahbaie, P.; Escolano, F.L.; Herrnberger, M.; et al. Complex regional pain syndrome patient immunoglobulin M has pronociceptive effects in the skin and spinal cord of tibia fracture mice. Pain 2020, 161, 797–809. [Google Scholar] [CrossRef]
- Luczynski, P.; Tramullas, M.; Viola, M.; Shanahan, F.; Clarke, G.; O’Mahony, S.; Dinan, T.G.; Cryan, J.F. Microbiota regulates visceral pain in the mouse. eLife 2017, 6, e25887. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, C.; Corleto, J.; Ruegger, P.M.; Logan, G.D.; Peacock, B.B.; Mendonca, S.; Yamaki, S.; Adamson, T.; Ermel, R.; McKemy, D.; et al. Dominant role of the gut microbiota in chemotherapy induced neuropathic pain. Sci. Rep. 2019, 9, 20324. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Lim, G.; You, Z.; Ding, W.; Huang, P.; Ran, C.; Doheny, J.; Caravan, P.; Tate, S.; Hu, K.; et al. Gut microbiota is critical for the induction of chemotherapy-induced pain. Nat. Neurosci. 2017, 20, 1213–1216. [Google Scholar] [CrossRef]
- Yang, C.; Fang, X.; Zhan, G.; Huang, N.; Li, S.; Bi, J.; Jiang, R.; Yang, L.; Miao, L.; Zhu, B.; et al. Key role of gut microbiota in anhedonia-like phenotype in rodents with neuropathic pain. Transl. Psychiatry 2019, 9, 57. [Google Scholar] [CrossRef]
- Guo, T.Z.; Shi, X.; Li, W.W.; Wei, T.; Clark, J.D.; Kingery, W.S. Passive transfer autoimmunity in a mouse model of complex regional pain syndrome. Pain 2017, 158, 2410–2421. [Google Scholar] [CrossRef]
- Clos-Garcia, M.; Andrés-Marin, N.; Fernández-Eulate, G.; Abecia, L.; Lavín, J.L.; van Liempd, S.; Cabrera, D.; Royo, F.; Valero, A.; Errazquin, N.; et al. Gut microbiome and serum metabolome analyses identify molecular biomarkers and altered glutamate metabolism in fibromyalgia. eBioMedicine 2019, 46, 499–511. [Google Scholar] [CrossRef]
- Boer, C.G.; Radjabzadeh, D.; Medina-Gomez, C.; Garmaeva, S.; Schiphof, D.; Arp, P.; Koet, T.; Kurilshikov, A.; Fu, J.; Ikram, M.A.; et al. Intestinal microbiome composition and its relation to joint pain and inflammation. Nat. Commun. 2019, 10, 4881. [Google Scholar] [CrossRef]
- Ware, M.A.; Bennett, G.J. Case report: Long-standing complex regional pain syndrome relieved by a cephalosporin antibiotic. Pain 2014, 155, 1412–1415. [Google Scholar] [CrossRef]
- Weinstock, L.B.; Myers, T.L.; Walters, A.S.; Schwartz, O.A.; Younger, J.W.; Chopra, P.J.; Guarino, A.H. Identification and treatment of new inflammatory triggers for complex regional pain syndrome. A A Case Rep. 2016, 6, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, A. Neuroinflammation in fibromyalgia and CRPS is multifactorial. Nat. Rev. Rheumatol. 2016, 12, 242. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.; Wallace, D.; Hallegua, D.; Chow, E.; Kong, Y.; Park, S.; Lin, H.C. A link between irritable bowel syndrome and fibromyalgia may be related to findings on lactulose breath testing. Ann. Rheum. Dis. 2004, 63, 450–452. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, T.T.; Monteleone, G. Immunity, inflammation, and allergy in the gut. Science 2005, 307, 1920–1925. [Google Scholar] [CrossRef]
- Holden, W.; Orchard, T.; Wordsworth, P. Enteropathic arthritis. Rheum. Dis. Clin. N. Am. 2003, 29, 513–530. [Google Scholar] [CrossRef]
- Maes, M.; Mihaylova, I.; Leunis, J.C. Increased serum IgA and IgM against LPS of enterobacteria in chronic fatigue syndrome (CFS): Indication for the involvement of Gram-negative enterobacteria in the etiology of CFS and for the presence of an increased gut–intestinal permeability. J. Affect. Disord. 2007, 99, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Üçeyler, N.; Valenza, R.; Stock, M.; Schedel, R.; Sprotte, G.; Sommer, C. Reduced levels of antiinflammatory cytokines in patients with chronic widespread pain. Arthritis Rheum. 2006, 54, 2656–2664. [Google Scholar] [CrossRef]
- Üçeyler, N.; Eberle, T.; Rolke, R.; Birklein, F.; Sommer, C. Differential expression patterns of cytokines in complex regional pain syndrome. Pain 2007, 132, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.J.; Linker-Israeli, M.; Hallegua, D.; Silverman, S.; Silver, D.; Weisman, M.H. Cytokines play an aetiopathogenetic role in fibromyalgia: A hypothesis and pilot study. Rheumatology 2001, 40, 743–749. [Google Scholar] [CrossRef]
- Hart, R.P.; Wade, J.B.; Martelli, M.F. Cognitive impairment in patients with chronic pain: The significance of stress. Curr. Pain Headache Rep. 2003, 7, 116–126. [Google Scholar] [CrossRef]
- Wallon, C.; Yang, P.C.; Keita, A.V.; Ericson, A.C.; McKay, D.M.; Sherman, P.M.; Perdue, M.H.; Söderholm, J.D. Corticotropin-releasing hormone (CRH) regulates macromolecular permeability via mast cells in normal human colonic biopsies in vitro. Gut 2008, 57, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Buret, A.G. How stress induces intestinal hypersensitivity. Am. J. Pathol. 2006, 168, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Goebel, A.; Buhner, S.; Schedel, R.; Lochs, H.; Sprotte, G. Altered intestinal permeability in patients with primary fibromyalgia and in patients with complex regional pain syndrome. Rheumatology 2008, 47, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Nair, G.B. Homeostasis and dysbiosis of the gut microbiome in health and disease. J. Biosci. 2019, 44, 117. [Google Scholar] [CrossRef]
- Sommer, F.; Anderson, J.M.; Bharti, R.; Raes, J.; Rosenstiel, P. The resilience of the intestinal microbiota influences health and disease. Nat. Rev. Microbiol. 2017, 15, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, K.M.; Hackshaw, K.V. Nutritional supplements for the treatment of neuropathic pain. Biomedicines 2021, 9, 674. [Google Scholar] [CrossRef]
- Pham, V.T.; Dold, S.; Rehman, A.; Bird, J.K.; Steinert, R.E. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr. Res. 2021, 95, 35–53. [Google Scholar] [CrossRef]
- Wan, Z.; Zheng, J.; Zhu, Z.; Sang, L.; Zhu, J.; Luo, S.; Zhao, Y.; Wang, R.; Zhang, Y.; Hao, K.; et al. Intermediate role of gut microbiota in vitamin B nutrition and its influences on human health. Front. Nutr. 2022, 9, 1031502. [Google Scholar] [CrossRef]
- Beane, K.E.; Redding, M.C.; Wang, X.; Pan, J.H.; Le, B.; Cicalo, C.; Jeon, S.; Kim, Y.J.; Lee, J.H.; Shin, E.C.; et al. Effects of dietary fibers, micronutrients, and phytonutrients on gut microbiome: A review. Appl. Biol. Chem. 2021, 64, 36. [Google Scholar] [CrossRef]
- Hossain, K.S.; Amarasena, S.; Mayengbam, S. B vitamins and their roles in gut health. Microorganisms 2022, 10, 1168. [Google Scholar] [CrossRef] [PubMed]
- Tangestani, H.; Boroujeni, H.K.; Djafarian, K.; Emamat, H.; Shab-Bidar, S. Vitamin D and the gut microbiota: A narrative literature review. Clin. Nutr. Res. 2021, 10, 181–191. [Google Scholar] [CrossRef]
- Habib, A.M.; Nagi, K.; Thillaiappan, N.B.; Sukumaran, V.; Akhtar, S. Vitamin D and its potential interplay with pain signaling pathways. Front. Immunol. 2020, 11, 820. [Google Scholar] [CrossRef]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Regulation of the intestinal barrier by nutrients: The role of tight junctions. Anim. Sci. J. 2020, 91, e13357. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Ma, J.; Piao, X.; Mahfuz, S.; Long, S.; Wang, J. The interaction among gut microbes, the intestinal barrier and short chain fatty acids. Anim. Nutr. 2022, 9, 159–174. [Google Scholar] [CrossRef]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Pickard, J.M.; Núñez, G. Pathogen colonization resistance in the gut and its manipulation for improved health. Am. J. Pathol. 2019, 189, 1300–1310. [Google Scholar] [CrossRef]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-chain fatty-acid-producing bacteria: Key components of the human gut microbiota. Nutrients 2023, 15, 2211. [Google Scholar] [CrossRef]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short chain fatty acids and its producing organisms: An overlooked therapy for IBD? eBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut–brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Martel, J.; Chang, S.H.; Ko, Y.F.; Hwang, T.L.; Young, J.D.; Ojcius, D.M. Gut barrier disruption and chronic disease. Trends Endocrinol. Metab. 2022, 33, 247–265. [Google Scholar] [CrossRef]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L.R.; Scaldaferri, F. Gut microbiota, intestinal permeability, and systemic inflammation: A narrative review. Intern. Emerg. Med. 2023. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.B.S.; Meng, J.; Zhang, J. Does low grade systemic inflammation have a role in chronic pain? Front. Mol. Neurosci. 2021, 14, 785214. [Google Scholar] [CrossRef] [PubMed]
- Stacy, A.; Andrade-Oliveira, V.; McCulloch, J.A.; Hild, B.; Oh, J.H.; Perez-Chaparro, P.J.; Sim, C.K.; Lim, A.I.; Link, V.M.; Enamorado, M.; et al. Infection trains the host for microbiota-enhanced resistance to pathogens. Cell 2021, 184, 615–627.e17. [Google Scholar] [CrossRef] [PubMed]
- Sethi, V.; Kurtom, S.; Tarique, M.; Lavania, S.; Malchiodi, Z.; Hellmund, L.; Zhang, L.; Sharma, U.; Giri, B.; Garg, B.; et al. Gut microbiota promotes tumor growth in mice by modulating immune response. Gastroenterology 2018, 155, 33–37.e6. [Google Scholar] [CrossRef]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; ter Horst, R.; Jansen, T.; Jacobs, L.; Bonder, M.J.; et al. Linking the human gut microbiome to inflammatory cytokine production capacity. Cell 2016, 167, 1125–1136.e8. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut microbiota: The neglected endocrine organ. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Yao, Z.; Yang, T. Intestinal microbiota-mediated dietary fiber bioavailability. Front. Nutr. 2022, 9, 1003571. [Google Scholar] [CrossRef]
- Feng, W.; Ao, H.; Peng, C. Gut microbiota, short-chain fatty acids, and herbal medicines. Front. Pharmacol. 2018, 9, 1354. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Kuenstner, J.T.; Chamberlin, W.; Naser, S.A.; Collins, M.T.; Dow, C.T.; Aitken, J.M.; Weg, S.; Telega, G.; John, K.; Haas, D.; et al. Resolution of Crohn’s disease and complex regional pain syndrome following treatment of paratuberculosis. World J. Gastroenterol. 2015, 21, 4048–4062. [Google Scholar] [CrossRef] [PubMed]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.H.; Sperandio, M.; Di Ciaula, A. Gut microbiota and short chain fatty acids: Implications in glucose homeostasis. Int. J. Mol. Sci. 2022, 23, 1105. [Google Scholar] [CrossRef]
- Visekruna, A.; Luu, M. The role of short-chain fatty acids and bile acids in intestinal and liver function, inflammation, and carcinogenesis. Front. Cell Dev. Biol. 2021, 9, 703218. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, R.; Suk, K.T. Therapeutic potential of human microbiome-based short-chain fatty acids and bile acids in liver disease. Livers 2022, 2, 139–145. [Google Scholar] [CrossRef]
- Akhtar, M.; Naqvi, S.U.A.S.; Liu, Q.; Pan, H.; Ma, Z.; Kong, N.; Chen, Y.; Shi, D.; Kulyar, M.F.E.A.; Khan, J.A.; et al. Short chain fatty acids (SCFAs) are the potential immunomodulatory metabolites in controlling Staphylococcus aureus-mediated mastitis. Nutrients 2022, 14, 3687. [Google Scholar] [CrossRef]
- Fock, E.; Parnova, R. Mechanisms of blood–brain barrier protection by microbiota-derived short-chain fatty acids. Cells 2023, 12, 657. [Google Scholar] [CrossRef]
- Barcelon, E.E.; Cho, W.H.; Jun, S.B.; Lee, S.J. Brain microglial activation in chronic pain-associated affective disorder. Front. Neurosci. 2019, 13, 213. [Google Scholar] [CrossRef] [PubMed]
- Beggs, S.; Trang, T.; Salter, M.W. P2X4R+ microglia drive neuropathic pain. Nat. Neurosci. 2012, 15, 1068–1073. [Google Scholar] [CrossRef]
- Keller, A.F.; Beggs, S.; Salter, M.W.; De Koninck, Y. Transformation of the output of spinal lamina I neurons after nerve injury and microglia stimulation underlying neuropathic pain. Mol. Pain 2007, 3, 27. [Google Scholar] [CrossRef] [PubMed]
- Tanga, F.Y.; Raghavendra, V.; DeLeo, J.A. Quantitative real-time RT-PCR assessment of spinal microglial and astrocytic activation markers in a rat model of neuropathic pain. Neurochem. Int. 2004, 45, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.W.; Castonguay, A.; Taylor, A.J.; Murphy, N.P.; Ghogha, A.; Cook, C.; Xue, L.; Olmstead, M.C.; De Koninck, Y.; Evans, C.J.; et al. Microglia disrupt mesolimbic reward circuitry in chronic pain. J. Neurosci. 2015, 35, 8442–8450. [Google Scholar] [CrossRef]
- Zhang, R.X.; Liu, B.; Wang, L.; Ren, K.; Qiao, J.T.; Berman, B.M.; Lao, L. Spinal glial activation in a new rat model of bone cancer pain produced by prostate cancer cell inoculation of the tibia. Pain 2005, 118, 125–136. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhou, L.J.; Ren, W.J.; Xin, W.J.; Li, Y.Y.; Zhang, T.; Liu, X.G. The direction of synaptic plasticity mediated by C-fibers in spinal dorsal horn is decided by Src-family kinases in microglia: The role of tumor necrosis factor-α. Brain. Behav. Immun. 2010, 24, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Walter, L.; Neumann, H. Role of microglia in neuronal degeneration and regeneration. Semin. Immunopathol. 2009, 31, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.K. Microglia as a source and target of cytokines. Glia 2002, 40, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, L.; Schwartzman, R.J.; Alexander, G. Spinal cord histopathological alterations in a patient with longstanding complex regional pain syndrome. Brain. Behav. Immun. 2009, 23, 85–91. [Google Scholar] [CrossRef]
- Jeon, S.Y.; Seo, S.; Lee, J.S.; Choi, S.H.; Lee, D.H.; Jung, Y.H.; Song, M.K.; Lee, K.J.; Kim, Y.C.; Kwon, H.W.; et al. [11C]-(R)-PK11195 positron emission tomography in patients with complex regional pain syndrome: A pilot study. Medicine 2017, 96, e5735. [Google Scholar] [CrossRef]
- Reichenberger, E.R.; Alexander, G.M.; Perreault, M.J.; Russell, J.A.; Schwartzman, R.J.; Hershberg, U.; Rosen, G. Establishing a relationship between bacteria in the human gut and complex regional pain syndrome. Brain. Behav. Immun. 2013, 29, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, S.; Robbe-Masselot, C.; Ait-Belgnaoui, A.; Mancuso, A.; Mercade-Loubière, M.; Salvador-Cartier, C.; Gillet, M.; Ferrier, L.; Loubière, P.; Dague, E.; et al. Stress disrupts intestinal mucus barrier in rats via mucin O-glycosylation shift: Prevention by a probiotic treatment. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G420–G429. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef]
- Defaye, M.; Gervason, S.; Altier, C.; Berthon, J.Y.; Ardid, D.; Filaire, E.; Carvalho, F.A. Microbiota: A novel regulator of pain. J. Neural Transm. 2020, 127, 445–465. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef]
- Kamada, N.; Seo, S.U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Costigan, M.; Scholz, J.; Woolf, C.J. Neuropathic pain: A maladaptive response of the nervous system to damage. Annu. Rev. Neurosci. 2009, 32, 1–32. [Google Scholar] [CrossRef]
- Watkins, L.R.; Maier, S.F. Immune regulation of central nervous system functions: From sickness responses to pathological pain. J. Intern. Med. 2005, 257, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Manichanh, C.; Rigottier-Gois, L.; Bonnaud, E.; Gloux, K.; Pelletier, E.; Frangeul, L.; Nalin, R.; Jarrin, C.; Chardon, P.; Marteau, P.; et al. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut 2006, 55, 205–211. [Google Scholar] [CrossRef]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Sanderson, J.D.; Churcher, C.; Parkes, G.C.; Hudspith, B.N.; Rayment, N.; Brostoff, J.; Parkhill, J.; Dougan, G.; Petrovska, L. High-throughput clone library analysis of the mucosa-associated microbiota reveals dysbiosis and differences between inflamed and non-inflamed regions of the intestine in inflammatory bowel disease. BMC Microbiol. 2011, 11, 7. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef]
- Cariello, R.; Federico, A.; Sapone, A.; Tuccillo, C.; Scialdone, V.R.; Tiso, A.; Miranda, A.; Portincasa, P.; Carbonara, V.; Palasciano, G.; et al. Intestinal permeability in patients with chronic liver diseases: Its relationship with the aetiology and the entity of liver damage. Dig. Liver Dis. 2010, 42, 200–204. [Google Scholar] [CrossRef]
- Varatharaj, A.; Galea, I. The blood–brain barrier in systemic inflammation. Brain. Behav. Immun. 2017, 60, 1–12. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, W.; Fang, H. Inflammation caused by peripheral immune cells across into injured mouse blood brain barrier can worsen postoperative cognitive dysfunction induced by isoflurane. BMC Cell Biol. 2018, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Piche, T.; Barbara, G.; Aubert, P.; Bruley des Varannes, S.; Dainese, R.; Nano, J.L.; Cremon, C.; Stanghellini, V.; De Giorgio, R.; Galmiche, J.P.; et al. Impaired intestinal barrier integrity in the colon of patients with irritable bowel syndrome: Involvement of soluble mediators. Gut 2009, 58, 196–201. [Google Scholar] [CrossRef]
- Ernberg, M.; Christidis, N.; Ghafouri, B.; Bileviciute-Ljungar, I.; Löfgren, M.; Bjersing, J.; Palstam, A.; Larsson, A.; Mannerkorpi, K.; Gerdle, B.; et al. Plasma cytokine levels in fibromyalgia and their response to 15 weeks of progressive resistance exercise or relaxation therapy. Mediat. Inflamm. 2018, 2018, 3985154. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, T.; Huys, G.; Brandt, E.; Swings, J. Temporal stability analysis of the microbiota in human feces by denaturing gradient gel electrophoresis using universal and group-specific 16S rRNA gene primers. FEMS Microbiol. Ecol. 2004, 48, 437–446. [Google Scholar] [CrossRef]
- Zoetendal, E.G.; Collier, C.T.; Koike, S.; Mackie, R.I.; Gaskins, H.R. Molecular ecological analysis of the gastrointestinal microbiota: A review. J. Nutr. 2004, 134, 465–472. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef]
- Collado, M.C.; Isolauri, E.; Salminen, S.; Sanz, Y. The impact of probiotic on gut health. Curr. Drug Metab. 2009, 10, 68–78. [Google Scholar] [CrossRef]
- Biţă, A.; Scorei, I.R.; Bălşeanu, T.A.; Ciocîlteu, M.V.; Bejenaru, C.; Radu, A.; Bejenaru, L.E.; Rău, G.; Mogoşanu, G.D.; Neamţu, J.; et al. New insights into boron essentiality in humans and animals. Int. J. Mol. Sci. 2022, 23, 9147. [Google Scholar] [CrossRef]
- Mitruţ, I.; Scorei, I.R.; Manolea, H.O.; Biţă, A.; Mogoantă, L.; Neamţu, J.; Bejenaru, L.E.; Ciocîlteu, M.V.; Bejenaru, C.; Rău, G.; et al. Boron-containing compounds in Dentistry: A narrative review. Rom. J. Morphol. Embryol. 2022, 63, 477–483. [Google Scholar] [CrossRef]
- Biţă, A.; Scorei, I.R.; Rangavajla, N.; Bejenaru, L.E.; Rău, G.; Bejenaru, C.; Ciocîlteu, M.V.; Dincă, L.; Neamţu, J.; Bunaciu, A.; et al. Diester chlorogenoborate complex: A new naturally occurring boron-containing compound. Inorganics 2023, 11, 112. [Google Scholar] [CrossRef]
- Scorei, I.R.; Bita, A.; Dinca, L.; Mogosanu, G.D.; Rangavajla, N. Borate Complexes of Chlorogenic Acid and Uses Thereof. United States Patent and Trademark Office (USPTO) Patent Application No. PCT/US22/78488, 21 October 2022. Available online: https://www.uspto.gov/patents (accessed on 10 September 2023).
- Das, B.C.; Thapa, P.; Karki, R.; Schinke, C.; Das, S.; Kambhampati, S.; Banerjee, S.K.; Van Veldhuizen, P.; Verma, A.; Weiss, L.M.; et al. Boron chemicals in diagnosis and therapeutics. Future Med. Chem. 2013, 5, 653–676. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.F.S.; Denny, W.A.; Dos Santos, J.L. Boron in drug design: Recent advances in the development of new therapeutic agents. Eur. J. Med. Chem. 2019, 179, 791–804. [Google Scholar] [CrossRef]
- Messner, K.; Vuong, B.; Tranmer, G.K. The boron advantage: The evolution and diversification of boron’s applications in medicinal chemistry. Pharmaceuticals 2022, 15, 264. [Google Scholar] [CrossRef]
- Ji, Y.C.; Sun, Q.; Fu, C.Y.; She, X.; Liu, X.C.; He, Y.; Ai, Q.; Li, L.Q.; Wang, Z.L. Exogenous autoinducer-2 rescues intestinal dysbiosis and intestinal inflammation in a neonatal mouse necrotizing enterocolitis model. Front. Cell. Infect. Microbiol. 2021, 11, 694395. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, S.; Han, Y.; Zhang, F.; Ren, M.; Hu, Q.; Ye, P.; Li, X.; Jin, E.; Li, S. Proteomic analysis of rat duodenum reveals the modulatory effect of boron supplementation on immune activity. Genes 2023, 14, 1560. [Google Scholar] [CrossRef]
- Fu, C.Y.; Li, L.Q.; Yang, T.; She, X.; Ai, Q.; Wang, Z.L. Autoinducer-2 may be a new biomarker for monitoring neonatal necrotizing enterocolitis. Front. Cell. Infect. Microbiol. 2020, 10, 140. [Google Scholar] [CrossRef]
- Zhang, L.; Li, S.; Liu, X.; Wang, Z.; Jiang, M.; Wang, R.; Xie, L.; Liu, Q.; Xie, X.; Shang, D.; et al. Sensing of autoinducer-2 by functionally distinct receptors in prokaryotes. Nat. Commun. 2020, 11, 5371. [Google Scholar] [CrossRef]
- Xavier, K.B.; Bassler, B.L. Interference with AI-2-mediated bacterial cell–cell communication. Nature 2005, 437, 750–753. [Google Scholar] [CrossRef]
- Chen, H.; Yan, C.H.; Zhan, Y.F.; Geng, L.T.; Zhu, L.L.; Gong, L.C.; Wang, J. Boron derivatives accelerate biofilm formation of recombinant Escherichia coli via increasing quorum sensing system autoinducer-2 activity. Int. J. Mol. Sci. 2022, 23, 8059. [Google Scholar] [CrossRef] [PubMed]
- Keizers, M.; Dobrindt, U.; Berger, M. A simple biosensor-based assay for quantitative autoinducer-2 analysis. ACS Synth. Biol. 2022, 11, 747–759. [Google Scholar] [CrossRef]
- Hunter, J.M.; Nemzer, B.V.; Rangavajla, N.; Biţă, A.; Rogoveanu, O.C.; Neamţu, J.; Scorei, I.R.; Bejenaru, L.E.; Rău, G.; Bejenaru, C.; et al. The fructoborates: Part of a family of naturally occurring sugar–borate complexes—Biochemistry, physiology, and impact on human health: A review. Biol. Trace Elem. Res. 2019, 188, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Estevez-Fregoso, E.; Farfán-García, E.D.; García-Coronel, I.H.; Martínez-Herrera, E.; Alatorre, A.; Scorei, R.I.; Soriano-Ursúa, M.A. Effects of boron-containing compounds in the fungal kingdom. J. Trace Elem. Med. Biol. 2021, 65, 126714. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, T.; Ishii, T.; Matsunaga, T. Isolation and structural characterization of rhamnogalacturonan II–borate complex from Pinus densiflora. J. Wood Sci. 1999, 45, 435–439. [Google Scholar] [CrossRef]
- Sun, A.; Xu, Q.; Ren, L.; Cao, G.; Gou, D. Non-equilibrium ultrasound-assisted solid–liquid extraction of boron present in different phases within plants by ICP-OES. RSC Adv. 2017, 7, 49890–49894. [Google Scholar] [CrossRef]
- Donoiu, I.; Militaru, C.; Obleagă, O.; Hunter, J.M.; Neamţu, J.; Biţă, A.; Scorei, I.R.; Rogoveanu, O.C. Effects of boron-containing compounds on cardiovascular disease risk factors—A review. J. Trace Elem. Med. Biol. 2018, 50, 47–56. [Google Scholar] [CrossRef]
- Ascenso, O.S.; Torcato, I.M.; Miguel, A.S.; Marques, J.C.; Xavier, K.B.; Ventura, M.R.; Maycock, C.D. Synthesis and biological activity of a potent optically pure autoinducer-2 quorum sensing agonist. Bioorg. Chem. 2019, 85, 75–81. [Google Scholar] [CrossRef]
- Vijay, A.; Valdes, A.M. Role of the gut microbiome in chronic diseases: A narrative review. Eur. J. Clin. Nutr. 2022, 76, 489–501. [Google Scholar] [CrossRef]
- Kim, Y.S.; Unno, T.; Kim, B.Y.; Park, M.S. Sex differences in gut microbiota. World J. Mens Health 2020, 38, 48–60. [Google Scholar] [CrossRef]
- Prejac, J.; Skalny, A.A.; Grabeklis, A.R.; Uzun, S.; Mimica, N.; Momčilović, B. Assessing the boron nutritional status by analyzing its cumulative frequency distribution in the hair and whole blood. J. Trace Elem. Med. Biol. 2018, 45, 50–56. [Google Scholar] [CrossRef]
- Ahmed, S.; Spence, J.D. Sex differences in the intestinal microbiome: Interactions with risk factors for atherosclerosis and cardiovascular disease. Biol. Sex Differ. 2021, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, L.J.; Monga, M.; Miller, A.W. Defining dysbiosis for a cluster of chronic diseases. Sci. Rep. 2019, 9, 12918. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Wang, H.; Zhou, Y.; Zhang, H.; Zhou, H.; Zhang, X. Slimy partners: The mucus barrier and gut microbiome in ulcerative colitis. Exp. Mol. Med. 2021, 53, 772–787. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Kostanski, L.; Ketelson, H.; Meadows, D.; Pelton, R. Hydroxypropyl guar−borate interactions with tear film mucin and lysozyme. Langmuir 2005, 21, 10032–10037. [Google Scholar] [CrossRef]
- Wang, X.; Xia, N.; Liu, L. Boronic acid-based approach for separation and immobilization of glycoproteins and its application in sensing. Int. J. Mol. Sci. 2013, 14, 20890–20912. [Google Scholar] [CrossRef]
- Thompson, J.A.; Oliveira, R.A.; Djukovic, A.; Ubeda, C.; Xavier, K.B. Manipulation of the quorum sensing signal AI-2 affects the antibiotic-treated gut microbiota. Cell Rep. 2015, 10, 1861–1871. [Google Scholar] [CrossRef] [PubMed]
- Ragonnaud, E.; Biragyn, A. Gut microbiota as the key controllers of “healthy” aging of elderly people. Immun. Ageing 2021, 18, 2. [Google Scholar] [CrossRef]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef]
- Saraswati, S.; Sitaraman, R. Aging and the human gut microbiota—From correlation to causality. Front. Microbiol. 2015, 5, 764. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Jeffery, I.B. Gut microbiota and aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Catalkaya, G.; Venema, K.; Lucini, L.; Rocchetti, G.; Delmas, D.; Daglia, M.; De Filippis, A.; Xiao, H.; Quiles, J.L.; Xiao, J.; et al. Interaction of dietary polyphenols and gut microbiota: Microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 2020, 1, 109–133. [Google Scholar] [CrossRef]
- Plamada, D.; Vodnar, D.C. Polyphenols–gut microbiota interrelationship: A transition to a new generation of prebiotics. Nutrients 2021, 14, 137. [Google Scholar] [CrossRef]
- Ray, S.K.; Mukherjee, S. Evolving interplay between dietary polyphenols and gut microbiota—An emerging importance in healthcare. Front. Nutr. 2021, 8, 634944. [Google Scholar] [CrossRef] [PubMed]
- Amiri, P.; Hosseini, S.A.; Ghaffari, S.; Tutunchi, H.; Ghaffari, S.; Mosharkesh, E.; Asghari, S.; Roshanravan, N. Role of butyrate, a gut microbiota derived metabolite, in cardiovascular diseases: A comprehensive narrative review. Front. Pharmacol. 2022, 12, 837509. [Google Scholar] [CrossRef] [PubMed]
- Amiri, P.; Arefhosseini, S.; Bakhshimoghaddam, F.; Jamshidi Gurvan, H.; Hosseini, S.A. Mechanistic insights into the pleiotropic effects of butyrate as a potential therapeutic agent on NAFLD management: A systematic review. Front. Nutr. 2022, 9, 1037696. [Google Scholar] [CrossRef]
- Paone, P.; Cani, P.D. Mucus barrier, mucins and gut microbiota: The expected slimy partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Suriano, F.; Nyström, E.E.L.; Sergi, D.; Gustafsson, J.K. Diet, microbiota, and the mucus layer: The guardians of our health. Front. Immunol. 2022, 13, 953196. [Google Scholar] [CrossRef]
- Zhu, L.B.; Zhang, Y.C.; Huang, H.H.; Lin, J. Prospects for clinical applications of butyrate-producing bacteria. World J. Clin. Pediatr. 2021, 10, 84–92. [Google Scholar] [CrossRef]
- Pietrzkowski, Z. Compositions and Methods Related to Calcitriol. United States Patent and Trademark Office (USPTO) Patent Application No. US 2014/0349951 A1, 27 November 2014. Available online: https://patents.google.com/patent/US20140349951 (accessed on 12 September 2023).
- Thomas, R.L.; Jiang, L.; Adams, J.S.; Xu, Z.Z.; Shen, J.; Janssen, S.; Ackermann, G.; Vanderschueren, D.; Pauwels, S.; Knight, R.; et al. Vitamin D metabolites and the gut microbiome in older men. Nat. Commun. 2020, 11, 5997. [Google Scholar] [CrossRef] [PubMed]
- Turkez, H.; Arslan, M.E.; Tatar, A.; Mardinoglu, A. Promising potential of boron compounds against glioblastoma: In vitro antioxidant, anti-inflammatory and anticancer studies. Neurochem. Int. 2021, 149, 105137. [Google Scholar] [CrossRef] [PubMed]
- Ahlawat, S.; Asha; Sharma, K.K. Gut–organ axis: A microbial outreach and networking. Lett. Appl. Microbiol. 2021, 72, 636–668. [Google Scholar] [CrossRef] [PubMed]
- Córdova-Chávez, R.I.; Carrasco-Ruiz, M.F.; Rodríguez-Vera, D.; Pérez-Capistran, T.; Tamay-Cach, F.; Scorei, I.R.; Abad-García, A.; Soriano-Ursúa, M.A. Boron-containing compounds for prevention, diagnosis, and treatment of human metabolic disorders. Biol. Trace Elem. Res. 2023, 201, 2222–2239. [Google Scholar] [CrossRef]
- Sizmaz, O.; Koksal, B.H.; Yildiz, G. Rumen microbial fermentation, protozoan abundance and boron availability in yearling rams fed diets with different boron concentrations. J. Anim. Feed Sci. 2017, 26, 59–64. [Google Scholar] [CrossRef]
- Mayorga-Ramos, A.; Barba-Ostria, C.; Simancas-Racines, D.; Guamán, L.P. Protective role of butyrate in obesity and diabetes: New insights. Front. Nutr. 2022, 9, 1067647. [Google Scholar] [CrossRef]
- Bivar Xavier, K. Bacterial interspecies quorum sensing in the mammalian gut microbiota. Comptes Rendus Biol. 2018, 341, 297–299. [Google Scholar] [CrossRef]
- Hsiao, A.; Ahmed, A.M.S.; Subramanian, S.; Griffin, N.W.; Drewry, L.L.; Petri, W.A., Jr.; Haque, R.; Ahmed, T.; Gordon, J.I. Members of the human gut microbiota involved in recovery from Vibrio cholerae infection. Nature 2014, 515, 423–426. [Google Scholar] [CrossRef]
- Chen, J.; Vitetta, L. The role of butyrate in attenuating pathobiont-induced hyperinflammation. Immune Netw. 2020, 20, e15. [Google Scholar] [CrossRef]
- Rettner, R.E.; Saier, M.H., Jr. The autoinducer-2 exporter superfamily. J. Mol. Microbiol. Biotechnol. 2010, 18, 195–205. [Google Scholar] [CrossRef]
- Bridgeman, S.C.; Northrop, W.; Melton, P.E.; Ellison, G.C.; Newsholme, P.; Mamotte, C.D.S. Butyrate generated by gut microbiota and its therapeutic role in metabolic syndrome. Pharmacol. Res. 2020, 160, 105174. [Google Scholar] [CrossRef] [PubMed]
- Jourova, L.; Anzenbacherova, E.; Dostal, Z.; Anzenbacher, P.; Briolotti, P.; Rigal, E.; Daujat-Chavanieu, M.; Gerbal-Chaloin, S. Butyrate, a typical product of gut microbiome, affects function of the AhR gene, being a possible agent of crosstalk between gut microbiome, and hepatic drug metabolism. J. Nutr. Biochem. 2022, 107, 109042. [Google Scholar] [CrossRef]
- Kyner, D.; Zabos, P.; Christman, J.; Acs, G. Effect of sodium butyrate on lymphocyte activation. J. Exp. Med. 1976, 144, 1674–1678. [Google Scholar] [CrossRef]
- Kibbie, J.J.; Dillon, S.M.; Thompson, T.A.; Purba, C.M.; McCarter, M.D.; Wilson, C.C. Butyrate directly decreases human gut lamina propria CD4 T cell function through histone deacetylase (HDAC) inhibition and GPR43 signaling. Immunobiology 2021, 226, 152126. [Google Scholar] [CrossRef]
- Siddiqui, M.T.; Cresci, G.A.M. The immunomodulatory functions of butyrate. J. Inflamm. Res. 2021, 14, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, S.J.A.; Oliveira, M.M.B.; Ribeiro, S.B.; de Medeiros, C.A.C.X.; Lima, M.L.S.; Guerra, G.C.B.; de Araújo Júnior, R.F.; de Sousa Junior, F.C.; Martins, A.A.; Paiva, D.F.F.; et al. The beneficial effects of Lacticaseibacillus casei on the small intestine and colon of Swiss mice against the deleterious effects of 5-fluorouracil. Front. Immunol. 2022, 13, 954885. [Google Scholar] [CrossRef]
- Duncan, S.H.; Holtrop, G.; Lobley, G.E.; Calder, A.G.; Stewart, C.S.; Flint, H.J. Contribution of acetate to butyrate formation by human faecal bacteria. Br. J. Nutr. 2004, 91, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Belzer, C.; Goossens, M.; Kleerebezem, M.; De Vos, W.M.; Thas, O.; De Weirdt, R.; Kerckhof, F.M.; Van de Wiele, T. Butyrate-producing Clostridium cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. 2013, 7, 949–961. [Google Scholar] [CrossRef]
- Corrêa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The two-way polyphenols–microbiota interactions and their effects on obesity and related metabolic diseases. Front. Nutr. 2019, 6, 188. [Google Scholar] [CrossRef]
- Wang, X.; Qi, Y.; Zheng, H. Dietary polyphenol, gut microbiota, and health benefits. Antioxidants 2022, 11, 1212. [Google Scholar] [CrossRef] [PubMed]
- Schlechte, J.; Zucoloto, A.Z.; Yu, I.L.; Doig, C.J.; Dunbar, M.J.; McCoy, K.D.; McDonald, B. Dysbiosis of a microbiota–immune metasystem in critical illness is associated with nosocomial infections. Nat. Med. 2023, 29, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Sizmaz, Ö.; Köksal, B.H.; Yildiz, G. Rumen fermentation characteristics of rams fed supplemental boric acid and humic acid diets. Ankara Üniv. Vet. Fak. Derg. 2022, 69, 337–340. [Google Scholar] [CrossRef]
- Özkoç, M.; Can, B.; Şentürk, H.; Burukoğlu Dönmez, D.; Kanbak, G. Possible curative effects of boric acid and Bacillus clausii treatments on TNBS-induced ulcerative colitis in rats. Biol. Trace Elem. Res. 2023, 201, 1237–1251. [Google Scholar] [CrossRef]
- Thompson, J.A.; Oliveira, R.A.; Xavier, K.B. Chemical conversations in the gut microbiota. Gut Microbes 2016, 7, 163–170. [Google Scholar] [CrossRef]
- Nicholson, L.B. The immune system. Essays Biochem. 2016, 60, 275–301. [Google Scholar] [CrossRef]
- Delves, P.J.; Roitt, I.M. The immune system. First of two parts. N. Engl. J. Med. 2000, 343, 37–49. [Google Scholar] [CrossRef]
- Delves, P.J.; Roitt, I.M. The immune system. Second of two parts. N. Engl. J. Med. 2000, 343, 108–117. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota–gut–brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Góralczyk-Bińkowska, A.; Szmajda-Krygier, D.; Kozłowska, E. The microbiota–gut–brain axis in psychiatric disorders. Int. J. Mol. Sci. 2022, 23, 11245. [Google Scholar] [CrossRef]
- Thangaleela, S.; Sivamaruthi, B.S.; Kesika, P.; Bharathi, M.; Chaiyasut, C. Role of the gut–brain axis, gut microbial composition, diet, and probiotic intervention in Parkinson’s disease. Microorganisms 2022, 10, 1544. [Google Scholar] [CrossRef]
- Rutsch, A.; Kantsjö, J.B.; Ronchi, F. The gut–brain axis: How microbiota and host inflammasome influence brain physiology and pathology. Front. Immunol. 2020, 11, 604179. [Google Scholar] [CrossRef]
- Nandwana, V.; Nandwana, N.K.; Das, Y.; Saito, M.; Panda, T.; Das, S.; Almaguel, F.; Hosmane, N.S.; Das, B.C. The role of microbiome in brain development and neurodegenerative diseases. Molecules 2022, 27, 3402. [Google Scholar] [CrossRef] [PubMed]
- Bekker, R.A.; Bykov, Y.V. Boron preparations in psychiatry and neurology: Their rise, fall and renewed interest. Acta Biomed. Sci. 2018, 3, 85–100. [Google Scholar] [CrossRef]
- Estevez-Fregoso, E.; Kilic, A.; Rodríguez-Vera, D.; Nicanor-Juárez, L.E.; Romero-Rizo, C.E.M.; Farfán-García, E.D.; Soriano-Ursúa, M.A. Effects of boron-containing compounds on liposoluble hormone functions. Inorganics 2023, 11, 84. [Google Scholar] [CrossRef]
- Penland, J.G. Dietary boron, brain function, and cognitive performance. Environ. Health Perspect. 1994, 102, 65–72. [Google Scholar] [CrossRef]
- Penland, J.G. The importance of boron nutrition for brain and psychological function. Biol. Trace Elem. Res. 1998, 66, 299–317. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, M.W.; Alim, I.; Bultman, S.J.; Ratan, R.R. Butyrate, neuroepigenetics and the gut microbiome: Can a high fiber diet improve brain health? Neurosci. Lett. 2016, 625, 56–63. [Google Scholar] [CrossRef]
- Fernando, W.M.A.D.B.; Martins, I.J.; Morici, M.; Bharadwaj, P.; Rainey-Smith, S.R.; Lim, W.L.F.; Martins, R.N. Sodium butyrate reduces brain amyloid-β levels and improves cognitive memory performance in an Alzheimer’s disease transgenic mouse model at an early disease stage. J. Alzheimers Dis. 2020, 74, 91–99. [Google Scholar] [CrossRef]
- Alpino, G.C.Á.; Pereira-Sol, G.A.; Dias, M.M.E.; Aguiar, A.S.; Peluzio, M.D.C.G. Beneficial effects of butyrate on brain functions: A view of epigenetic. Crit. Rev. Food Sci. Nutr. 2022, 1–10. [Google Scholar] [CrossRef]
- Tang, J.; Zheng, X.; Xiao, K.; Wang, K.; Wang, J.; Wang, Y.; Wang, K.; Wang, W.; Lu, S.; Yang, K.; et al. Effect of boric acid supplementation on the expression of BDNF in African ostrich chick brain. Biol. Trace Elem. Res. 2016, 170, 208–215. [Google Scholar] [CrossRef]
- Turkez, H.; Yıldırım, S.; Sahin, E.; Arslan, M.E.; Emsen, B.; Tozlu, O.O.; Alak, G.; Ucar, A.; Tatar, A.; Hacimuftuoglu, A.; et al. Boron compounds exhibit protective effects against aluminum-induced neurotoxicity and genotoxicity: In vitro and in vivo study. Toxics 2022, 10, 428. [Google Scholar] [CrossRef]
- Alak, G.; Turkez, H.; Ucar, A.; Yeltekin, A.C.; Ozgeris, F.B.; Parlak, V.; Atamanalp, M. Neuroprotective properties of borax against aluminum hydroxide-induced neurotoxicity: Possible role of Nrf-2/BDNF/AChE pathways in fish brain. Brain Res. 2023, 1803, 148241. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, S.; Wu, J.; Ye, T.; Wang, S.; Wang, P.; Xing, D. Butyrate-producing bacteria and the gut–heart axis in atherosclerosis. Clin. Chim. Acta 2020, 507, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Gao, X.; Peng, Y.; Wu, Q.; Zhu, J.; Tan, C.; Xia, G.; You, C.; Xu, R.; Pan, S.; et al. Higher risk of stroke is correlated with increased opportunistic pathogen load and reduced levels of butyrate-producing bacteria in the gut. Front. Cell. Infect. Microbiol. 2019, 9, 4. [Google Scholar] [CrossRef]
- Bouchareb, R.; Katz, M.; Saadallah, N.; Sassi, Y.; Ali, S.; Lebeche, D. Boron improves cardiac contractility and fibrotic remodeling following myocardial infarction injury. Sci. Rep. 2020, 10, 17138. [Google Scholar] [CrossRef]
- Weber, K.S.; Ratjen, I.; Enderle, J.; Seidel, U.; Rimbach, G.; Lieb, W. Plasma boron concentrations in the general population: A cross-sectional analysis of cardio-metabolic and dietary correlates. Eur. J. Nutr. 2022, 61, 1363–1375. [Google Scholar] [CrossRef]
- Nishii, Y.; Ichikawa, K.; Miyamoto, T.; Takeda, T.; Kobayashi, M.; Suzuki, S.; Sakurai, A.; Hashizume, K. n-Butyrate enhances induction of thyroid hormone-responsive nuclear protein. Endocr. J. 1993, 40, 515–521. [Google Scholar] [CrossRef]
- Su, X.; Zhao, Y.; Li, Y.; Ma, S.; Wang, Z. Gut dysbiosis is associated with primary hypothyroidism with interaction on gut–thyroid axis. Clin. Sci. 2020, 134, 1521–1535. [Google Scholar] [CrossRef]
- Kuru, R.; Yilmaz, S.; Balan, G.; Tuzuner, B.A.; Tasli, P.N.; Akyuz, S.; Yener Ozturk, F.; Altuntas, Y.; Yarat, A.; Sahin, F. Boron-rich diet may regulate blood lipid profile and prevent obesity: A non-drug and self-controlled clinical trial. J. Trace Elem. Med. Biol. 2019, 54, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Evariste, L.; Flahaut, E.; Baratange, C.; Barret, M.; Mouchet, F.; Pinelli, E.; Galibert, A.M.; Soula, B.; Gauthier, L. Ecotoxicological assessment of commercial boron nitride nanotubes toward Xenopus laevis tadpoles and host-associated gut microbiota. Nanotoxicology 2021, 15, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Fort, D.J.; Rogers, R.L.; McLaughlin, D.W.; Sellers, C.M.; Schlekat, C.L. Impact of boron deficiency on Xenopus laevis: A summary of biological effects and potential biochemical roles. Biol. Trace Elem. Res. 2002, 90, 117–142. [Google Scholar] [CrossRef]
- Ibrahim, T.B.; Abdel-Wahab, A.; Abdel Aziz, R.L.; El-Anwar, A.H.; Ibrahim, S.S. Dietary boron supplementation and its impact on testicular function, thyroid activity and serum calcium in rams. Small Rumin. Res. 2019, 174, 156–162. [Google Scholar] [CrossRef]
- Kan, F.; Kucukkurt, I. Investigation of the effect of boron on thyroid functions and biochemical parameters in hypothyroid induced-rats. J. Biochem. Mol. Toxicol. 2022, 36, e23186. [Google Scholar] [CrossRef]
- Knezevic, J.; Starchl, C.; Tmava Berisha, A.; Amrein, K. Thyroid–gut–axis: How does the microbiota influence thyroid function? Nutrients 2020, 12, 1769. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Wang, C.; Meng, F.; Wang, H.; Song, B.; Yang, Y.; Shan, Z. Association between gut microbiota and autoimmune thyroid disease: A systematic review and meta-analysis. Front. Endocrinol. 2021, 12, 774362. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Anh, N.D.Q.; Bossier, P.; Defoirdt, T. Norepinephrine and dopamine increase motility, biofilm formation, and virulence of Vibrio harveyi. Front. Microbiol. 2014, 5, 584. [Google Scholar] [CrossRef] [PubMed]
- Hamamah, S.; Aghazarian, A.; Nazaryan, A.; Hajnal, A.; Covasa, M. Role of microbiota–gut–brain axis in regulating dopaminergic signaling. Biomedicines 2022, 10, 436. [Google Scholar] [CrossRef]
- Jiang, W.; Lu, G.; Gao, D.; Lv, Z.; Li, D. The relationships between the gut microbiota and its metabolites with thyroid diseases. Front. Endocrinol. 2022, 13, 943408. [Google Scholar] [CrossRef]
- Jung, G.S.; Buehler, M.J. Multiscale modeling of muscular-skeletal systems. Annu. Rev. Biomed. Eng. 2017, 19, 435–457. [Google Scholar] [CrossRef]
- Newnham, R.E. Essentiality of boron for healthy bones and joints. Environ. Health Perspect. 1994, 102, 83–85. [Google Scholar] [CrossRef]
- Newnham, R. Discovering the cure for arthritis. Nutr. Health 2004, 17, 281–284. [Google Scholar] [CrossRef]
- Newnham, R.E. Agricultural practices affect arthritis. Nutr. Health 1991, 7, 89–100. [Google Scholar] [CrossRef]
- Gök, S.; Ozan, F.; Akay, E.; Yamak, K.; Kayalı, C.; Altay, T. Effect of boric acid on cartilage formation of osteochondral defects in rabbit knee: An experimental study. Ulus. Travma Acil Cerrahi Derg. 2021, 27, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Baqeri, M.; Kaka, G.; Sadraie, H.; Mofid, M.; Rezaei, S.; Naghii, M.R. Boron effect on articular cartilage repair in rat. Koomesh 1394, 17, 170–178. Available online: http://koomeshjournal.semums.ac.ir/article-1-2463-en.html (accessed on 15 September 2023).
- Scorei, I.D.; Scorei, R.I. Calcium fructoborate helps control inflammation associated with diminished bone health. Biol. Trace Elem. Res. 2013, 155, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.H.; Na, H.S.; Jhun, J.; Woo, J.S.; Lee, A.R.; Lee, S.Y.; Lee, J.S.; Um, I.G.; Kim, S.J.; Park, S.H.; et al. Lactobacillus (LA-1) and butyrate inhibit osteoarthritis by controlling autophagy and inflammatory cell death of chondrocytes. Front. Immunol. 2022, 13, 930511. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, G.; Wang, Y.; Jiang, R.; Li, Y.; Wang, H.; Wang, F.; Ma, H.; Cao, L. Microbial metabolite sodium butyrate attenuates cartilage degradation by restoring impaired autophagy and autophagic flux in osteoarthritis development. Front. Pharmacol. 2021, 12, 659597. [Google Scholar] [CrossRef]
- Cooney, O.D.; Nagareddy, P.R.; Murphy, A.J.; Lee, M.K.S. Healthy gut, healthy bones: Targeting the gut microbiome to promote bone health. Front. Endocrinol. 2021, 11, 620466. [Google Scholar] [CrossRef]
- Chen, C.; Dong, B.; Wang, Y.; Zhang, Q.; Wang, B.; Feng, S.; Zhu, Y. The role of Bacillus acidophilus in osteoporosis and its roles in proliferation and differentiation. J. Clin. Lab. Anal. 2020, 34, e23471. [Google Scholar] [CrossRef]
- Wallimann, A.; Magrath, W.; Pugliese, B.; Stocker, N.; Westermann, P.; Heider, A.; Gehweiler, D.; Zeiter, S.; Claesson, M.J.; Richards, R.G.; et al. Butyrate inhibits osteoclast activity in vitro and regulates systemic inflammation and bone healing in a murine osteotomy model compared to antibiotic-treated mice. Mediators Inflamm. 2021, 2021, 8817421. [Google Scholar] [CrossRef] [PubMed]
- Zemanova, N.; Omelka, R.; Mondockova, V.; Kovacova, V.; Martiniakova, M. Roles of gut microbiome in bone homeostasis and its relationship with bone-related diseases. Biology 2022, 11, 1402. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, S.; Musmeci, E.; Candeliere, F.; Amaretti, A.; Rossi, M. Identification of mucin degraders of the human gut microbiota. Sci. Rep. 2021, 11, 11094. [Google Scholar] [CrossRef] [PubMed]
- Glover, J.S.; Ticer, T.D.; Engevik, M.A. Characterizing the mucin-degrading capacity of the human gut microbiota. Sci. Rep. 2022, 12, 8456. [Google Scholar] [CrossRef]
- Xu, H.M.; Huang, H.L.; Xu, J.; He, J.; Zhao, C.; Peng, Y.; Zhao, H.L.; Huang, W.Q.; Cao, C.Y.; Zhou, Y.J.; et al. Cross-talk between butyric acid and gut microbiota in ulcerative colitis following fecal microbiota transplantation. Front. Microbiol. 2021, 12, 658292. [Google Scholar] [CrossRef] [PubMed]
- Stachowska, E.; Wiśniewska, M.; Dzieżyc, A.; Bohatyrewicz, A. Could the use of butyric acid have a positive effect on microbiota and treatment of type 2 diabetes? Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 4570–4578. [Google Scholar] [CrossRef] [PubMed]
- Dessordi, R.; Spirlandeli, A.L.; Zamarioli, A.; Volpon, J.B.; Navarro, A.M. Boron supplementation improves bone health of non-obese diabetic mice. J. Trace Elem. Med. Biol. 2017, 39, 169–175. [Google Scholar] [CrossRef]
- Chummun, N.H. The Relationship between Boron, Stress Hormones and Bone Markers in Humans of Different Ages, Life Style and Health Status. Ph.D. Thesis, University of Greenwich, London, UK, 2000. Available online: https://gala.gre.ac.uk/id/eprint/8728/ (accessed on 16 September 2023).
- Tyagi, A.M.; Yu, M.; Darby, T.M.; Vaccaro, C.; Li, J.Y.; Owens, J.A.; Hsu, E.; Adams, J.; Weitzmann, M.N.; Jones, R.M.; et al. The microbial metabolite butyrate stimulates bone formation via T regulatory cell-mediated regulation of WNT10B expression. Immunity 2018, 49, 1116–1131.e7. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biţă, C.E.; Scorei, I.R.; Vreju, A.F.; Muşetescu, A.E.; Mogoşanu, G.D.; Biţă, A.; Dinescu, V.C.; Dinescu, Ş.C.; Criveanu, C.; Bărbulescu, A.L.; et al. Microbiota-Accessible Boron-Containing Compounds in Complex Regional Pain Syndrome. Medicina 2023, 59, 1965. https://doi.org/10.3390/medicina59111965
Biţă CE, Scorei IR, Vreju AF, Muşetescu AE, Mogoşanu GD, Biţă A, Dinescu VC, Dinescu ŞC, Criveanu C, Bărbulescu AL, et al. Microbiota-Accessible Boron-Containing Compounds in Complex Regional Pain Syndrome. Medicina. 2023; 59(11):1965. https://doi.org/10.3390/medicina59111965
Chicago/Turabian StyleBiţă, Cristina Elena, Ion Romulus Scorei, Ananu Florentin Vreju, Anca Emanuela Muşetescu, George Dan Mogoşanu, Andrei Biţă, Venera Cristina Dinescu, Ştefan Cristian Dinescu, Cristina Criveanu, Andreea Lili Bărbulescu, and et al. 2023. "Microbiota-Accessible Boron-Containing Compounds in Complex Regional Pain Syndrome" Medicina 59, no. 11: 1965. https://doi.org/10.3390/medicina59111965