
Transcriptomic Profiling of Pleural Effusions: Differences in Malignant and Infectious Fluids
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Ethics Approval
2.2. Specimen Collection
2.3. RNA Sequencing
2.4. Bioinformatics Analysis and Statistics
3. Results
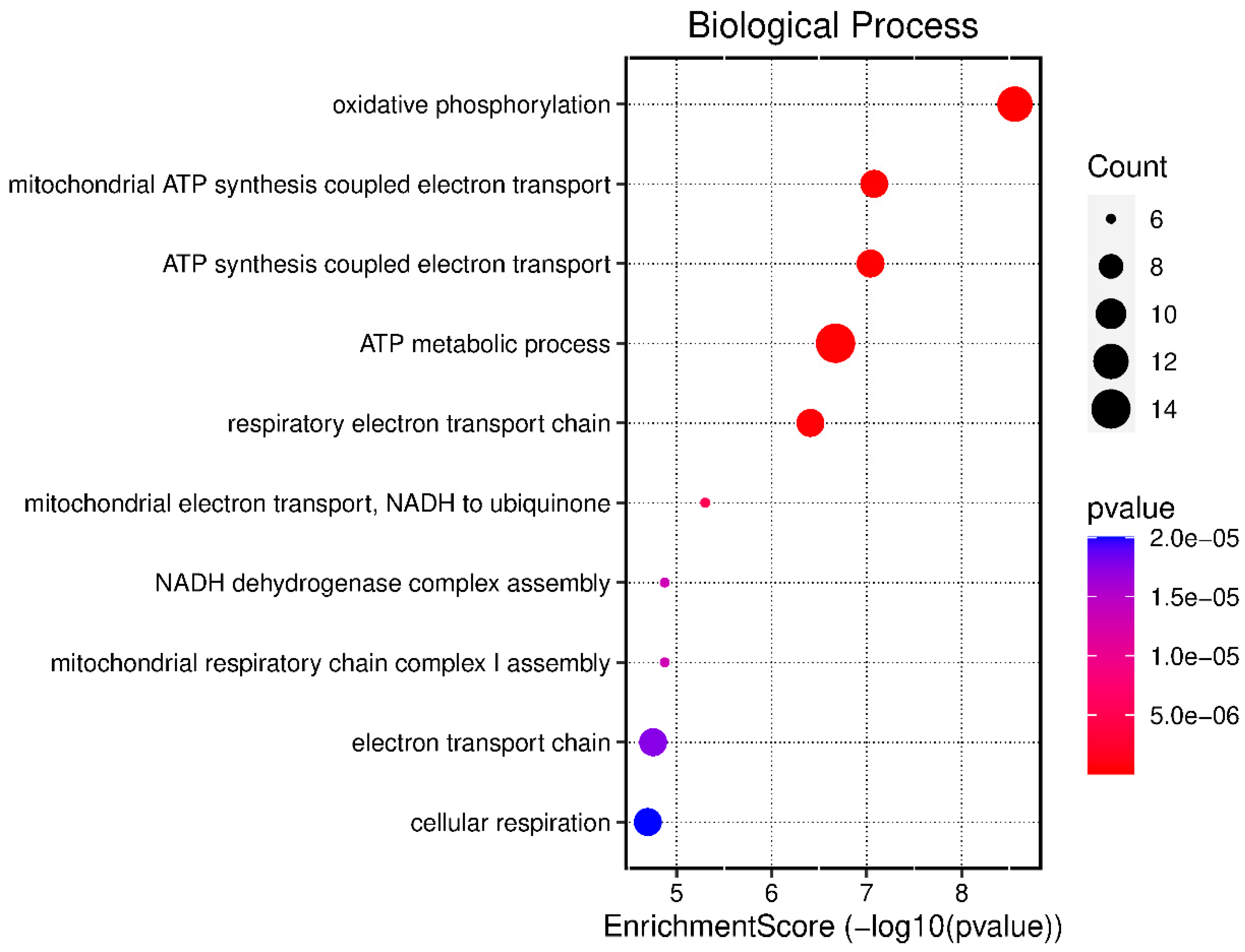
3.1. Biological Processes Expressed in Infectious Pleural Effusions
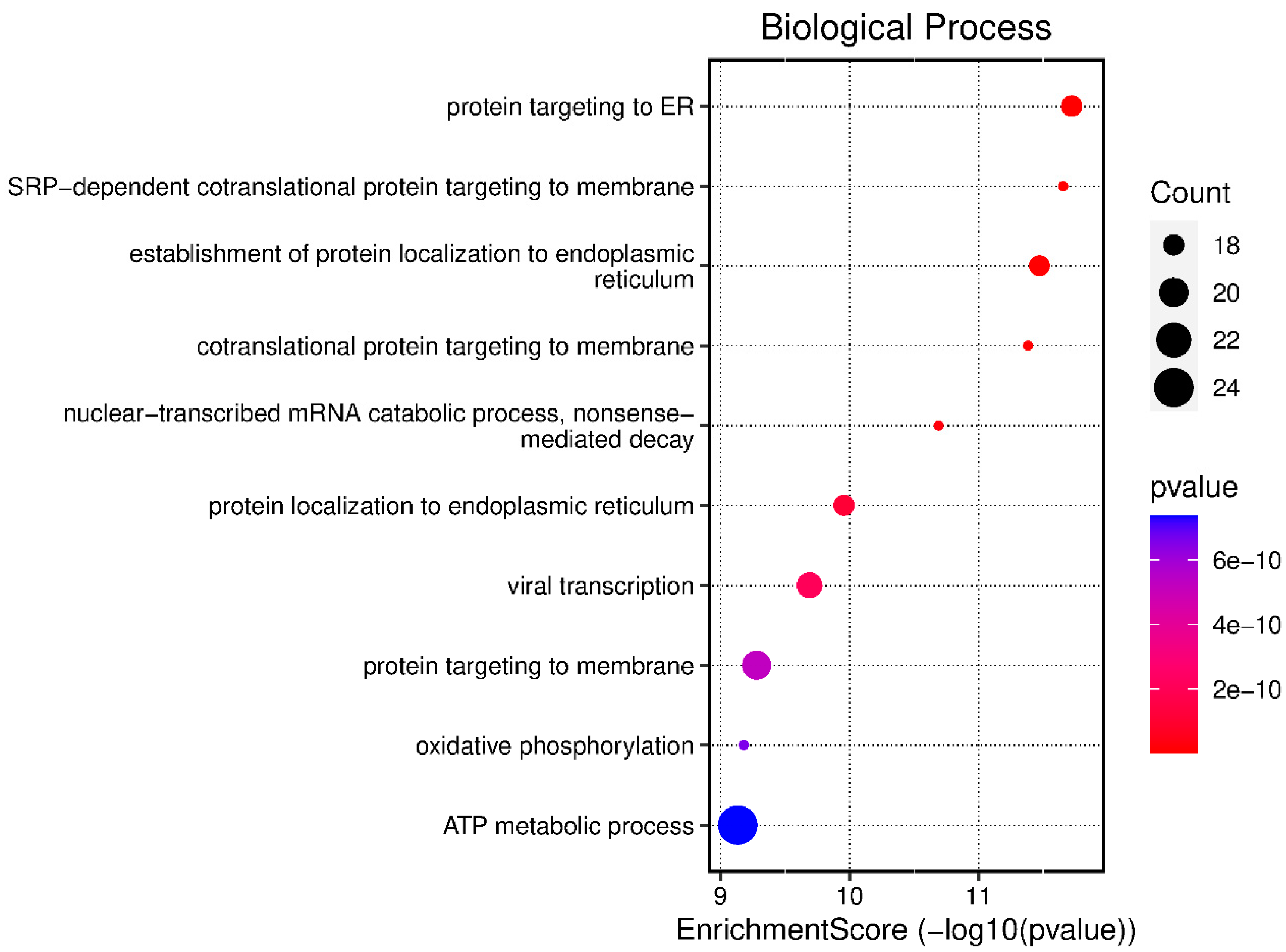
3.2. Biological Processes Expressed in Malignant Pleural Effusions
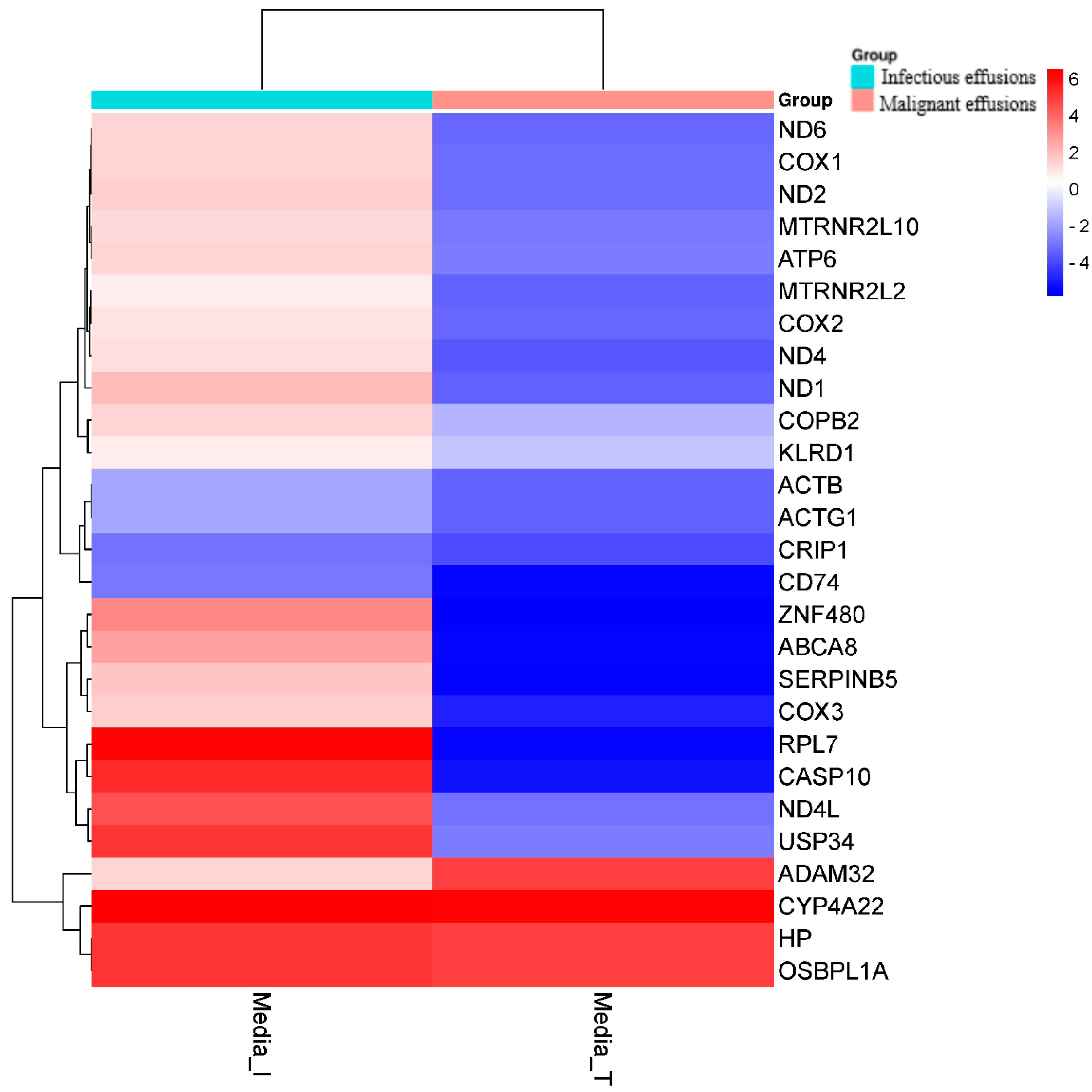
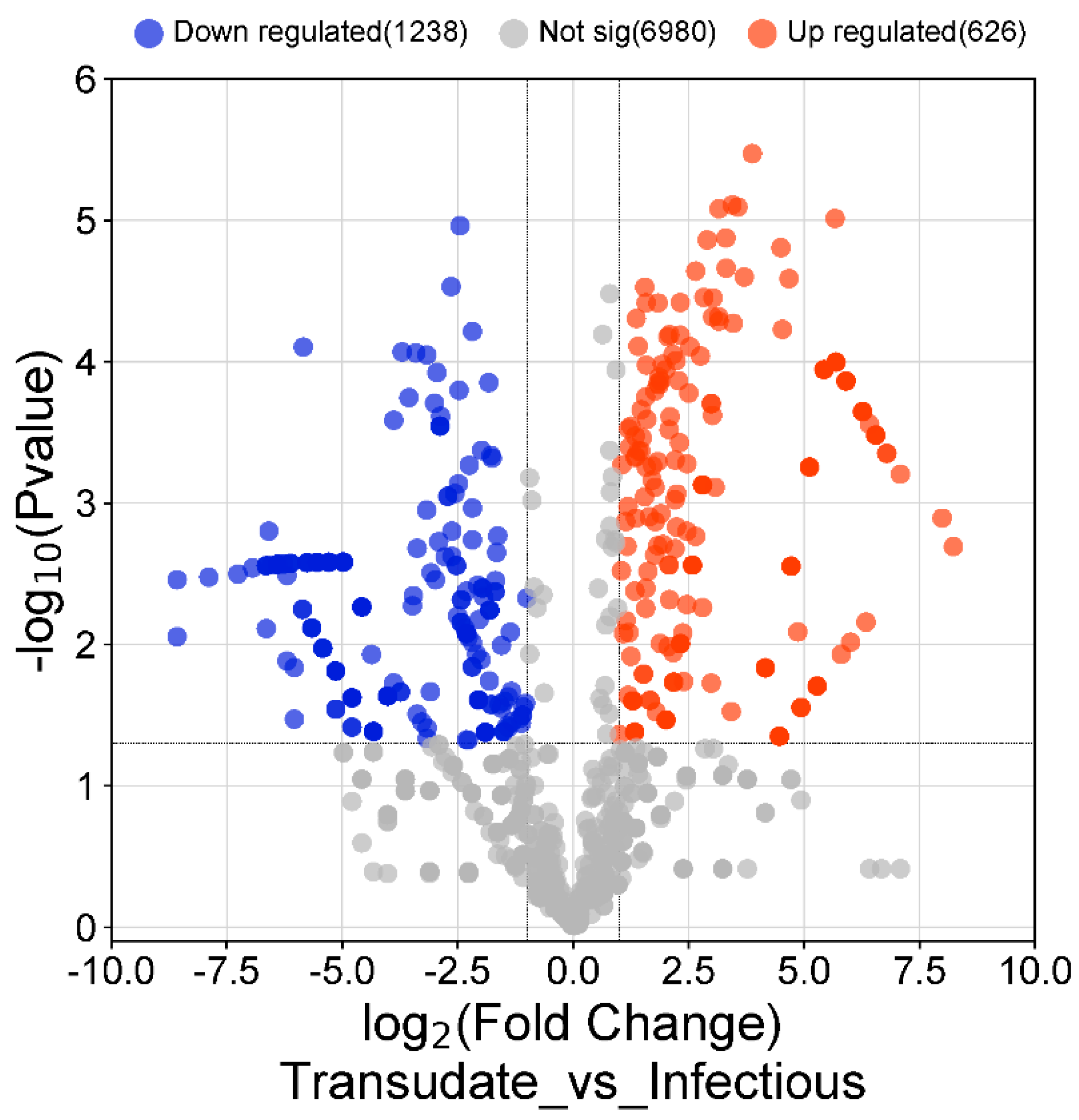
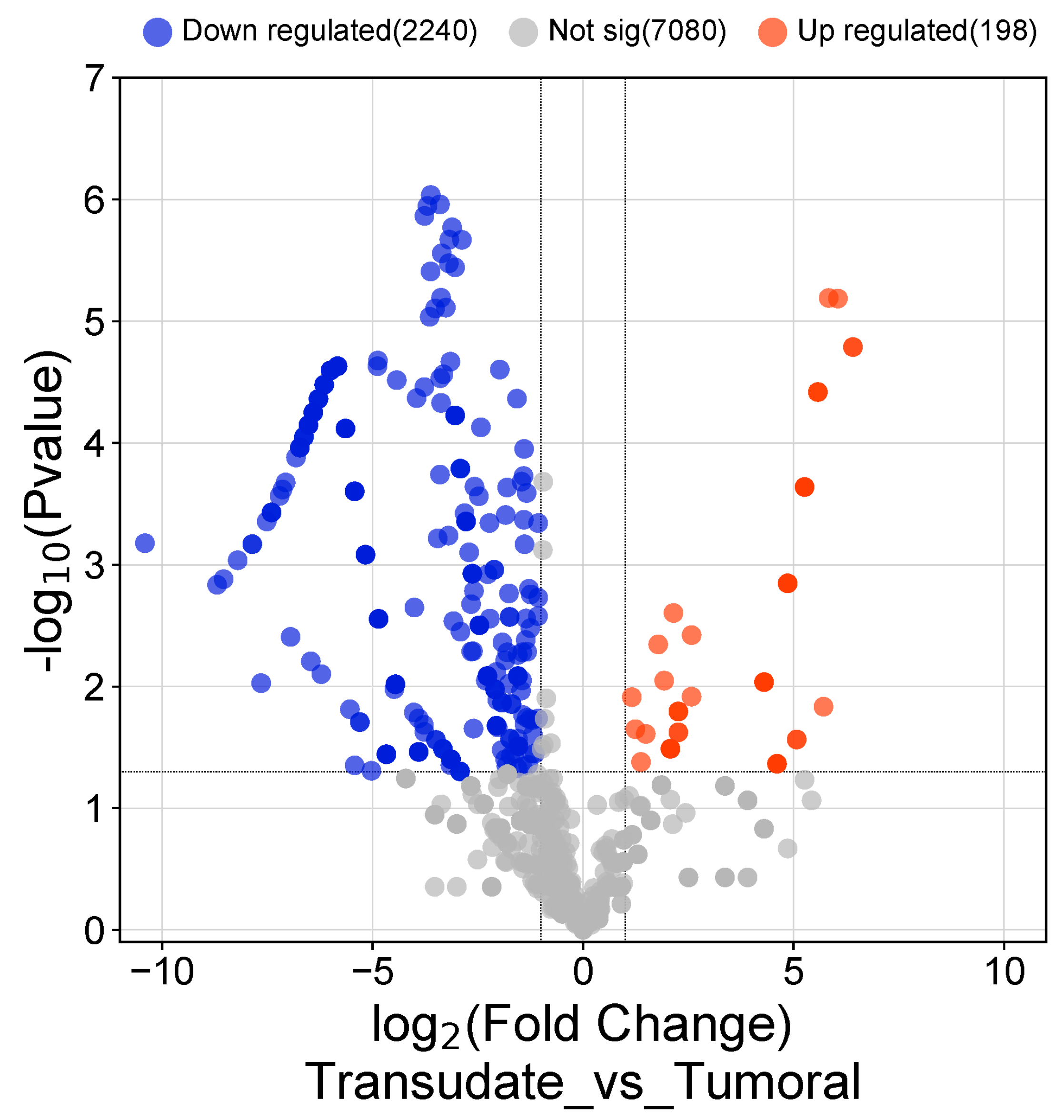
3.3. Differentially Expressed Genes in Malignant vs. Infectious Effusions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Villena-Garrido, V.; Ferrer-Sancho, J.; Hernández-Blasco, L.; de Pablo-Gafas, A.; Pérez-Rodríguez, E.; Rodríguez-Panadero, F.; Candeira, S.R.; Velázquez, Á.S.; Cuadrado, L.V.; Separ, T. Diagnóstico y tratamiento del derrame pleural. Arch. Bronconeumol. 2006, 42, 349–372. [Google Scholar] [CrossRef]
- Feller-Kopman, D.; Light, R. Pleural Disease. N. Engl. J. Med. 2018, 378, 740–751. [Google Scholar] [CrossRef]
- Porcel, J.M. Biomarkers in the diagnosis of pleural diseases: A 2018 update. Ther. Adv. Respir. Dis. 2018, 12, 1753466618808660. [Google Scholar] [CrossRef]
- Maskell, N.A.; Batt, S.; Hedley, E.L.; Davies, C.W.; Gillespie, S.H.; Davies, R.J. The bacteriology of pleural infection by genetic and standard methods and its mortality significance. Am. J. Respir Crit. Care Med. 2006, 174, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Supplitt, S.; Karpinski, P.; Sasiadek, M.; Laczmanska, I. Current achievements and applications of transcriptomics in personalized cancer medicine. Int. J. Mol. Sci. 2021, 22, 1422. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods. 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Bustin, S.A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol. 2000, 25, 169–193. [Google Scholar] [CrossRef]
- Kozian, D.H.; Kirschbaum, B.J. Comparative gene-expression analysis. Trends Biotechnol. 1999, 17, 73–78. [Google Scholar] [CrossRef]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial electron transport chain: Oxidative phosphorylation, oxidant production, and methods of measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef] [PubMed]
- Pool, M.R. Targeting of Proteins for Translocation at the Endoplasmic Reticulum. Int. J. Mol. Sci. 2022, 23, 3773. [Google Scholar] [CrossRef]
- Egea, P.F.; Stroud, R.M.; Walter, P. Targeting proteins to membranes: Structure of the signal recognition particle. Curr. Opin. Struct. Biol. 2005, 15, 213–220. [Google Scholar] [CrossRef]
- Cui, X.A.; Palazzo, A.F. Localization of mRNAs to the endoplasmic reticulum. Wiley Interdiscip. Rev. RNA 2014, 5, 481–492. [Google Scholar] [CrossRef]
- Bao, Q.L.; Li, J.; Sun, W.; Jiang, H.G.; Zhu, L.R.; Wang, Y. Diagnostic utility of LUNX mRNA and VEGF mRNA in pleural fluid for differentiating benign from malignant origin. Jpn. J. Clin. Oncol. 2014, 44, 1198–1205. [Google Scholar] [CrossRef]
- Hsu, S.C.; Chang, S.Y.; Hwang, Y.T.; Terng, H.J.; Tsai, C.L.; Shen, C.H.; Huang, S.K.; Chian, C.F. mRNA markers associated with malignant pleural effusion. Sci. Rep. 2023, 13, 6677. [Google Scholar] [CrossRef]
- Łukaszewicz-Zając, M.; Pączek, S.; Mroczko, B. A disintegrin and metalloproteinase (ADAM) family-novel biomarkers of selected gastrointestinal (GI) malignancies? Cancers 2022, 14, 2307. [Google Scholar] [CrossRef] [PubMed]
- Hida, Y.; Hamada, J. Differential expressions of matrix metalloproteinases, a disintegrin and metalloproteinases, and a disintegrin and metalloproteinases with thrombospondin motifs and their endogenous inhibitors among histologic subtypes of lung cancers. Anticancer. Agents Med. Chem. 2012, 12, 744–752. [Google Scholar] [CrossRef] [PubMed]
- DiGiusto, D.L.; Cannon, P.M.; Holmes, M.C.; Li, L.; Rao, A.; Wang, J.; Lee, G.; Gregory, P.D.; A Kim, K.; Hayward, S.B.; et al. Preclinical development and qualification of ZFN-mediated CCR5 disruption in human hematopoietic stem/progenitor cells. Mol. Ther. Methods Clin. Dev. 2016, 3, 16067. [Google Scholar] [CrossRef]
- Villet, R.A.; Truong-Bolduc, Q.C.; Wang, Y.; Estabrooks, Z.; Medeiros, H.; Hooper, D.C. Regulation of expression of abcA and its response to environmental conditions. J. Bacteriol. 2014, 196, 1532–1539. [Google Scholar] [CrossRef]
- Bao, J.; Pan, G.; Poncz, M.; Wei, J.; Ran, M.; Zhou, Z. Serpin functions in host-pathogen interactions. PeerJ 2018, 6, e4557. [Google Scholar] [CrossRef]
- Nakamura, A.; Fukuda, S.; Kusuki, M.; Watari, H.; Shimura, S.; Kimura, K.; Nishi, I.; Komatsu, M.; The Study of Bacterial Resistance in the Kinki region of japan (SBRK) group. Evaluation of five Legionella urinary antigen detection kits including new Ribotest Legionella for simultaneous detection of ribosomal protein L7/L12. J. Infect. Chemother. 2021, 27, 1533–1535. [Google Scholar] [CrossRef]
- Runthala, A. Protein structure prediction: Challenging targets for CASP10. J. Biomol. Struct. Dyn. 2012, 30, 607–615. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Štibrániová, I.; Bartíková, P.; Holíková, V.; Kazimírová, M. Deciphering biological processes at the tick-host interface opens new strategies for treatment of human diseases. Front. Physiol. 2019, 1, 830. [Google Scholar] [CrossRef] [PubMed]
- Leone, R.D.; Powell, J.D. Metabolism of immune cells in cancer. Nat. Rev. Cancer. 2020, 20, 516–531. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Makkar, R.; Anwer, K.; Hassani, R.; Khuwaja, G.; Khalid, A.; Mohan, S.; Alhazmi, H.A.; Sachdeva, M.; Rachamalla, M. Mitochondrial dysfunction: A cellular and molecular hub in pathology of metabolic diseases and infection. J. Clin. Med. 2023, 12, 2882. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CHRACTERISTICS | Transudates (11) | Infectious (16) | Malignant (19) |
|---|---|---|---|
| Male, n(%) | 3 (27.27%) | 9 (56.25%) | 11 (57.89%) |
| Age, year | 79.18 ± 11.6 | 63.38 ± 12.3 | 65.89 ± 13.9 |
| DISEASES | Congestive heart failure (7) | Paraneumonic (13) | Lung cáncer (6) |
| Another transudate causes (4) | Another infectious (3) | Breast cancer (4) | |
| Another cáncer (9) |
| ID | Description | p Value | Gene ID |
|---|---|---|---|
| GO:0006119 | oxidative phosphorylation | 2.75522 × 10−9 | ATP8/ND4L/ND1/COX3/PPIF/ND4/ND6/COX1/ATP6/ND3/COX2/ND2 |
| GO:0042775 | mitochondrial ATP synthesis coupled electron transport | 8.32541 × 10−8 | ND4L/ND1/COX3/ND4/ND6/COX1/ND3/COX2/ND2 |
| GO:0042773 | ATP synthesis coupled electron transport | 9.10531 × 10−8 | ND4L/ND1/COX3/ND4/ND6/COX1/ND3/COX2/ND2 |
| GO:0046034 | ATP metabolic process | 2.1265 × 10−7 | ATP8/ND4L/ND1/GAPDH/COX3/PPIF/ND4/ND6/COX1/ATP6/ND3/COX2/AK9/ND2 |
| GO:0022904 | respiratory electron transport chain | 3.90402 × 10−7 | ND4L/ND1/COX3/ND4/ND6/COX1/ND3/COX2/ND2 |
| GO:0006120 | mitochondrial electron transport, NADH to ubiquinone | 4.9925 × 10−6 | ND4L/ND1/ND4/ND6/ND3/ND2 |
| GO:0010257 | NADH dehydrogenase complex assembly | 1.33194× 10−5 | ND1/ND4/ND6/ND3/NDUFAF8/ND2 |
| GO:0032981 | mitochondrial respiratory chain complex I assembly | 1.33194 × 10−5 | ND1/ND4/ND6/ND3/NDUFAF8/ND2 |
| GO:0022900 | electron transport chain | 1.76432 × 10−5 | ND4L/ND1/COX3/ND4/ND6/COX1/ND3/COX2/ND2 |
| GO:0045333 | cellular respiration | 2.00624 × 10−5 | ND4L/ND1/COX3/ND4/ND6/COX1/ND3/COX2/ND2 |
| ID | Description | p Value | Gene ID |
|---|---|---|---|
| O:0045047 | protein targeting to ER | 1.90903 × 10−12 | RPL8/RPL36/RPS6/RPL32/RPL30/RPL15/ RPL6/RPL13/RPS20/RPS12/RPL7/RPS14/ RPLP2/RPL28/SEC63/HSPA5/RPL37/RPL29 |
| GO:0006614 | SRP-dependent cotranslational protein targeting to membrane | 2.21935 × 10−12 | RPL8/RPL36/RPS6/RPL32/RPL30/RPL15/ RPL6/RPL13/RPS20/RPS12/RPL7/RPS14/ RPLP2/RPL28/SEC63/RPL37/RPL29 |
| GO:0072599 | establishment of protein localization to endoplasmic reticulum | 3.39106 × 10−12 | RPL8/RPL36/RPS6/RPL32/RPL30/RPL15/ RPL6/RPL13/RPS20/RPS12/RPL7/RPS14/ RPLP2/RPL28/SEC63/HSPA5/RPL37/RPL29 |
| GO:0006613 | cotranslational protein targeting to membrane | 4.15652 × 10−12 | RPL8/RPL36/RPS6/RPL32/RPL30/ RPL15/RPL6/RPL13/RPS20/RPS12/RPL7/ RPS14/RPLP2/RPL28/SEC63/RPL37/RPL29 |
| GO:0000184 | nuclear-transcribed mRNA catabolic process, nonsense-mediated decay | 2.04422 × 10−11 | RPL8/RPL36/RPS6/RPL32/RPL30/ SMG1/RPL15/RPL6/RPL13/RPS20/ RPS12/RPL7/RPS14/RPLP2/RPL28/RPL37/RPL29 |
| GO:0070972 | protein localization to endoplasmic reticulum | 1.10638 × 10−10 | RPL8/RPL36/RPS6/RPL32/RPL30/ RPL15/RPL6/RPL13/RPS20/RPS12/RPL7/ RPS14/RPLP2/RPL28/SEC63/HSPA5/RPL37/RPL29 |
| GO:0019083 | viral transcription | 2.04724 × 10−10 | NELFB/RPL8/RPL36/RPS6/RPL32/ RPL30/RPL15/RPL6/RPL13/RPS20/RPS12 RPL7/RPS14/RPLP2/POM121C/RPL28/ TARDBP/RPL37/RPL29 |
| GO:0006612 | protein targeting to membrane | 5.26086 × 10−10 | RPL8/RPL36/RPS6/RPL32/RPL30/KCNB1/ RPL15/RPL6/RPL13/RPS20/RPS12/RPL7/ RPS14/RPLP2/RPL28/SEC63/HSPA5/ RPL37/RPL29/ITGB2 |
| GO:0006119 | oxidative phosphorylation | 6.61515 × 10−10 | COX1/ND4/ND6/COX3/COX2/ND1/ PINK1/ATP5PD/ND5/ND4L/CYTB/ SDHC/ATP6/ND2/VCP/PDE12/FXN |
| GO:0046034 | ATP metabolic process | 7.36436 × 10−10 | COX1/ND4/ND6/COX3/COX2/ND1/ SLC4A1/PINK1/ATP5PD/PFKFB4/ ND5/ND4L/HSPA8/CYTB/PKM/SDHC/ POM121C/ATP6/ND2/ENO1/ALDOA/ VCP/PDE12/FXN |
| List of the Top 20 Up-Regulated Molecules in the Infectious Pleural Effussion | |||
| Gene Symbol | Gene Name | logFC | p-Value |
| UBE2S | Ubiquitin Conjugating Enzyme E2S | 7.08513297 | 0.03788703 |
| ST6GALNAC5 | ST6 N-Acetylgalactosaminide Alpha-2,6-Sialyltransferase 5 | 6.26329843 | 0.0257876 |
| SMAD4 | Mothers against decapentaplegic homolog 4 | 6.26329843 | 0.0257876 |
| RPL7A | Ribosomal Protein L7a | 6.26329843 | 0.0257876 |
| RGSL1 | (Regulator Of G Protein Signaling Like 1 | 6.54938616 | 0.03166481 |
| PGBD1 | PiggyBac Transposable Element-Derived 1 | 6.54938616 | 0.03166481 |
| PARP15 | Poly(ADP-Ribose) Polymerase Family Member 15 | 5.90608458 | 0.01969065 |
| NLGN4Y | Neuroligin 4 Y-linked | 5.90608458 | 0.01969065 |
| LPAL2 | Lipoprotein(a) like 2 (pseudogene) | 6.78802257 | 0.03396501 |
| LOC100419170 | Toll like receptor 2 pseudogene | 6.26329843 | 0.0257876 |
| L3MBTL3 | Lethal(3)malignant brain tumor-like protein 3 | 5.68769633 | 0.01856111 |
| FRA10AC1 | FRA10A Associated CGG Repeat 1 | 6.54938616 | 0.03166481 |
| DNAJC10 | DnaJ Heat Shock Protein Family (Hsp40) Member C10 | 6.78802257 | 0.03396501 |
| DGKE | Diacylglycerol Kinase Epsilon, | 5.68769633 | 0.01856111 |
| DDB1 | Damage Specific DNA Binding Protein 1 | 5.68769633 | 0.01856111 |
| CYP4A22 | Cytochrome P450 Family 4 Subfamily A Member 22 | 6.54938616 | 0.03166481 |
| CMC1 | COX assembly mitochondrial protein 1 homolog | 5.90608458 | 0.01969065 |
| CCNC | Cyclin-C | 5.90608458 | 0.01969065 |
| ATP10B | ATPase Phospholipid Transporting 10B | 6.41342214 | 0.02907238 |
| ARF4 | ADP Ribosylation Factor 4 | 5.68769633 | 0.01856111 |
| List of the Top 20 Down-Regulated Molecules in the Infectious Pleural Effussion | |||
| Gene Symbol | Gene Name | logFC | p-Value |
| NCL | Lipofuscionosis neuronal ceroidea | −5.84529934 | 0.01856111 |
| C15orf40 | Chromosome 15 opne reding frame 40 | −3.88554066 | 0.027804565 |
| DCTN3 | Dynactin subunit 3 protein | −3.70467213 | 0.01856111 |
| RPL10 | Ribosomal Protein L10 | −3.55664134 | 0.023162056 |
| HADHA | Hydroxyacil CoA dehydrogenase trifunctional mutienzyme complex subunit alpha | −3.40781869 | 0.01856111 |
| PPFIA1 | Protein Tyrosine Phosphatase receptor type F polypeptide | −3.16943063 | 0.01856111 |
| CRIP1 | Cysteine-Rich protein 1 human recombinant | −3.00284513 | 0.02430739 |
| CTSB | Cathepsin B | −2.95194295 | 0.019054027 |
| CD74 | Cluster of differentiation 74 | −2.88370022 | 0.029072383 |
| LEPROTL1 | Leptin receptor overlapping transcript like 1 | −2.88370022 | 0.029072383 |
| PPIF | Peptidylprolyl Isomerase F | −2.88370022 | 0.029072383 |
| MX2 | MX dynamin like GTPase 2 | −2.87009164 | 0.027066857 |
| ALOX5AP | Arachidonate 5-lipoxygenase activating protein | −2.71636567 | 0.049560165 |
| POM121L15P | POM121 transmembrane nucleoporin like 15, pseudogen | −2.71636567 | 0.049560165 |
| RNF213 | Ring fing protein 213 | −2.64251631 | 0.017582598 |
| RPL4 | Large ribosomal subunit protein L4 | −2.56186959 | 0.048122461 |
| KPNB1 | Karyopherin subunit beta 1 | −2.48646613 | 0.043130785 |
| GAPDH | Glyceraldeyde-3-phosphatase deshydrogenase | −2.47163268 | 0.021680223 |
| PCSK5 | Proprotein convertase subtilisin/kexin type 5 | −2.45241044 | 0.015410669 |
| CORO1C | Coronin 1c | −2.24808151 | 0.033965007 |
| List of the Top 20 Up-Regulated Molecules in the Tumoral Pleural Effussion | |||
| Gene Symbol | Gene Name | logFC | p-Value |
| CYP4A22 | Cytochrome P450 family 4 subfamily A member 22 | 6.410457063 | 0.005199019 |
| NELFB | Negative elongation factor complex member B | 6.410457063 | 0.005199019 |
| ESCO1 | Establishment of sister chromatid cohesion N-acetyltransferase 1 | 6.052724677 | 0.004716469 |
| ZCCHC24 | Zinc finger CCHC-type containing 24 | 5.833949909 | 0.004716469 |
| CYB5R3 | Cytochrome b5 reductase 3 | 5.575965026 | 0.006286137 |
| PFKFB4 | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4 | 5.575965026 | 0.006286137 |
| ZNF43 | Zinc finger protein 43 | 5.575965026 | 0.006286137 |
| RIN2 | Ras and Rab interactor 2 | 5.261578232 | 0.012908359 |
| GRM2 | Glutamate metabotropic receptor 2 | 5.261578232 | 0.012908359 |
| WDR72 | WD repeat domain 72 | 5.261578232 | 0.012908359 |
| SLC25A30 | Solute carrier family 25 member 30 | 5.261578232 | 0.012908359 |
| PTPN20 | Protein tyrosine phosphatase non-receptor type 20 | 5.261578232 | 0.012908359 |
| HLA-C | Major histocompatibility complex, class I, C | 5.261578232 | 0.012908359 |
| SULT1C2P1 | Sulfotransferase Family 1C | 4.859022623 | 0.041574119 |
| HP | Haptoglobine | 4.859022623 | 0.041574119 |
| AGAP12P | ArfGAP GTPase, ankyrin repeat PH domain 12, pseudogene | 4.859022623 | 0.041574119 |
| SSX4B | SSX family member 4B | 4.859022623 | 0.041574119 |
| LOC642929 | General Transcription Factor II, I Pseudogen | 4.859022623 | 0.041574119 |
| List of the Top 20 Down-Regulated Molecules in the Tumoral Pleural Effussion | |||
| Gene Symbol | Gene Name | logFC | p-Value |
| SAMD12 | Sterile Alpha Motif Domain Containing 12 | −10.40752199 | 0.027309577 |
| C1orf116 | Chormosome 1 Open Reading Frame 116 | −8.692680487 | 0.042610742 |
| FANCI | Fanconi Anemia Complementary group 1 | −8.538352536 | 0.041574119 |
| NASP | Nuclear Autoantigenic Sperm Protein | −8.203765305 | 0.030086669 |
| GLIPR1L2 | GLI pathogenesis-related 1 like 2 | −7.8571759 | 0.027309577 |
| COL28A1 | Collagen Type XXVIII Alpha 1 Chain | −7.8571759 | 0.027309577 |
| CLU | Clustering | −7.514895677 | 0.021227484 |
| FOXN3 | Forkhead Box N3 | −7.400075641 | 0.018726604 |
| AGBL5 | Cytosolic carboxypeptidase-like protein 5 | −7.400075641 | 0.018726604 |
| PABIR3 | PABIR family number 3 | −7.400075641 | 0.018726604 |
| ANKS6 | Ankyrin Repeat And Sterile Alpha Motif Domain Containing 6 | −7.208650249 | 0.013964234 |
| CARS1 | Cysteinyl-TRNA Syntethase 1 | −7.138748507 | 0.012908359 |
| TRIM73 | Tripartite Motif Containing 73 | −7.065286645 | 0.012908359 |
| TECPR2 | Tectonin Beta-Propeller Repeat Containing 2 | −6.819377539 | 0.010672183 |
| PSMD11 | Proteasome 26S Subunit, Not-ATPase 11 | −6.727119503 | 0.008987272 |
| SMAD6 | Supressor of Mothers against Decaapentaplegic 6 | −6.727119503 | 0.008987272 |
| SCML4 | Scm Polycomb GroupnProtein Like 4 | −6.727119503 | 0.008987272 |
| MFSD14CP | Major Facilitator Superfamjily Domain Containing 14C | −6.727119503 | 0.008987272 |
| GBP4 | Guanilate Binding Protein 4 | −6.628556208 | 0.007560163 |
| XKR9 | XK erlated protein 9 | −6.628556208 | 0.007560163 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamora-Molina, L.; García-Pachón, E.; Amorós, M.; Gijón-Martínez, J.; Sánchez-Almendro, J.; Baeza-Martínez, C.; Hernández-Blasco, L.; Galiana, A. Transcriptomic Profiling of Pleural Effusions: Differences in Malignant and Infectious Fluids. Medicina 2024, 60, 424. https://doi.org/10.3390/medicina60030424
Zamora-Molina L, García-Pachón E, Amorós M, Gijón-Martínez J, Sánchez-Almendro J, Baeza-Martínez C, Hernández-Blasco L, Galiana A. Transcriptomic Profiling of Pleural Effusions: Differences in Malignant and Infectious Fluids. Medicina. 2024; 60(3):424. https://doi.org/10.3390/medicina60030424
Chicago/Turabian StyleZamora-Molina, Lucía, Eduardo García-Pachón, Marta Amorós, Julia Gijón-Martínez, Judith Sánchez-Almendro, Carlos Baeza-Martínez, Luis Hernández-Blasco, and Antonio Galiana. 2024. "Transcriptomic Profiling of Pleural Effusions: Differences in Malignant and Infectious Fluids" Medicina 60, no. 3: 424. https://doi.org/10.3390/medicina60030424