
Actinoflavosides B–D, Flavonoid Type Glycosides from Tidal Mudflat-Derived Actinomyces
Abstract
:1. Introduction
2. Results and Discussion
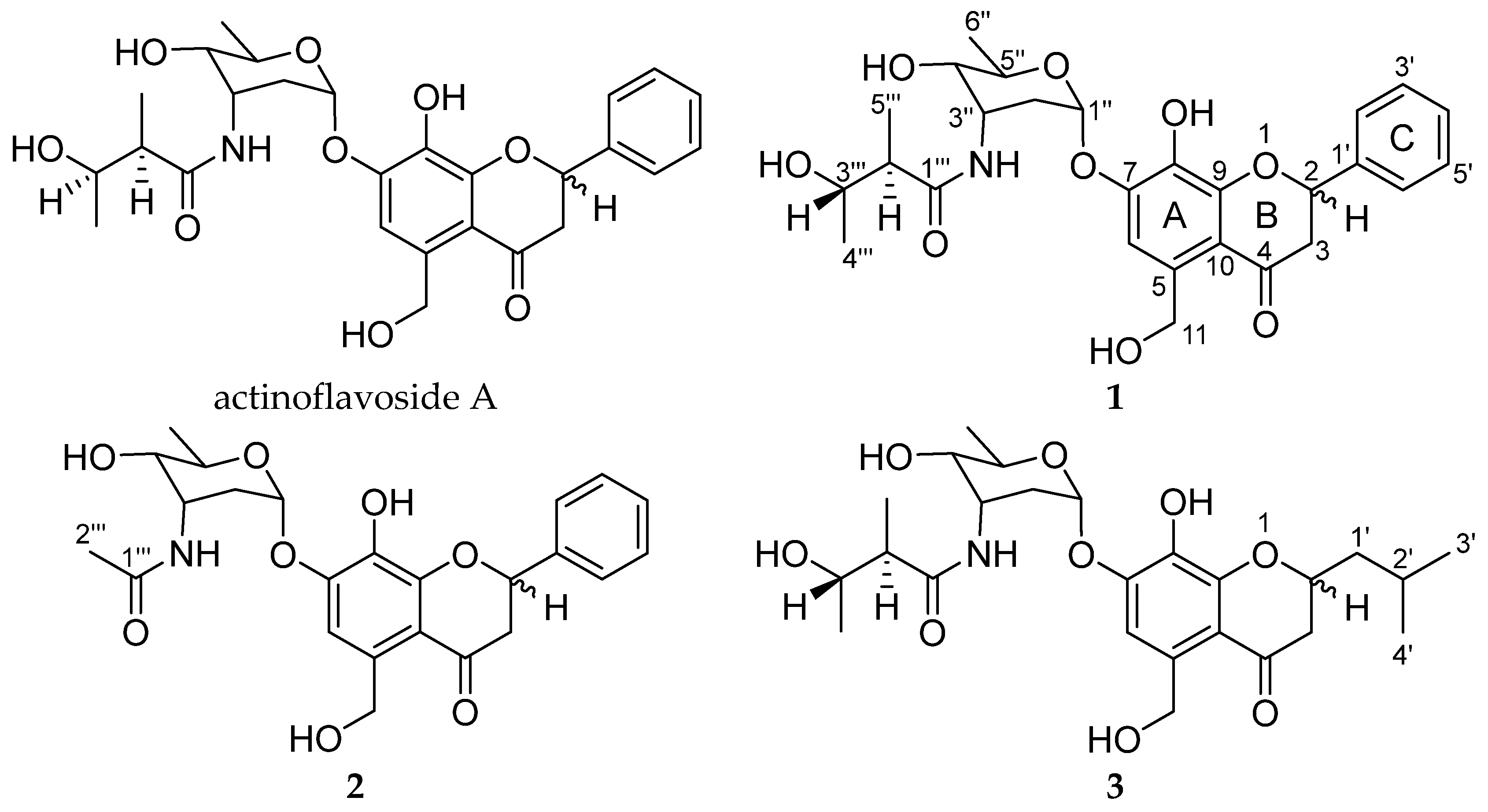
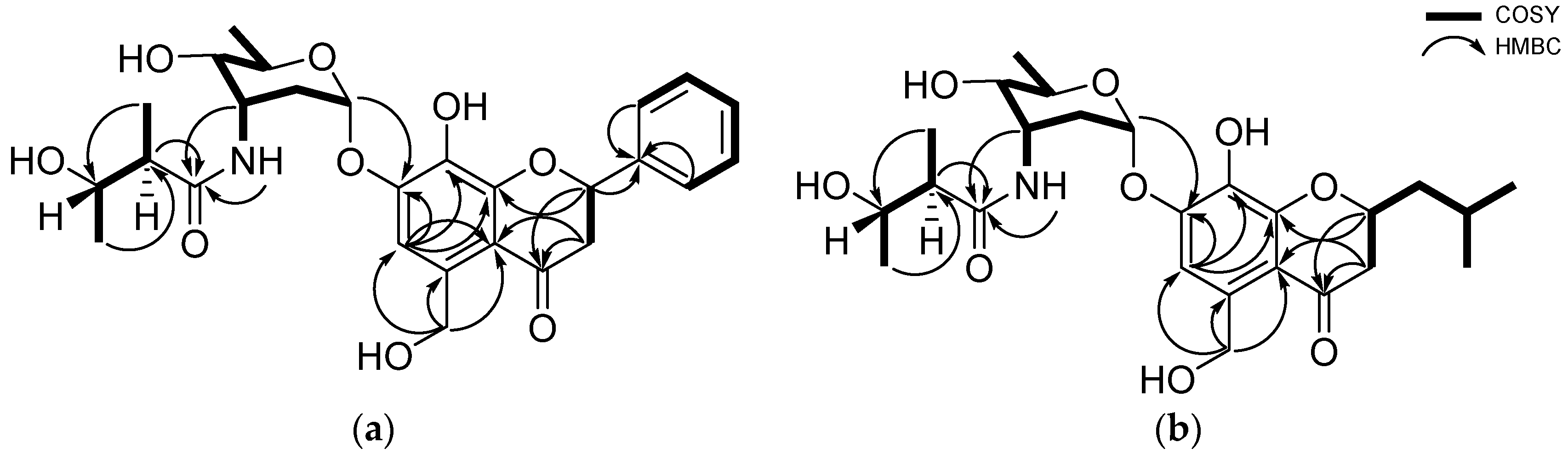
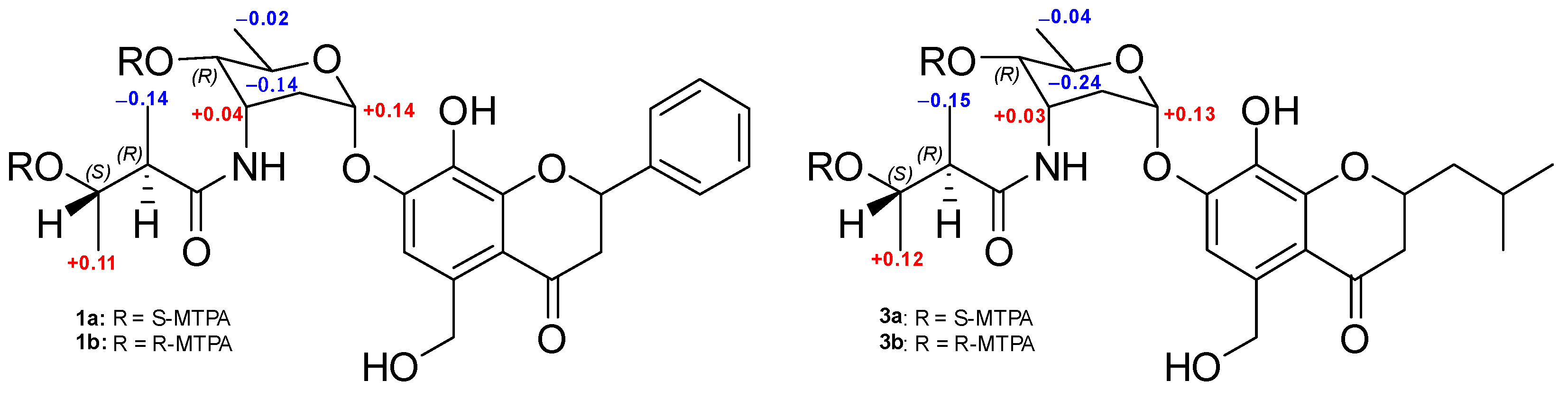
2.1. Structural Elucidation
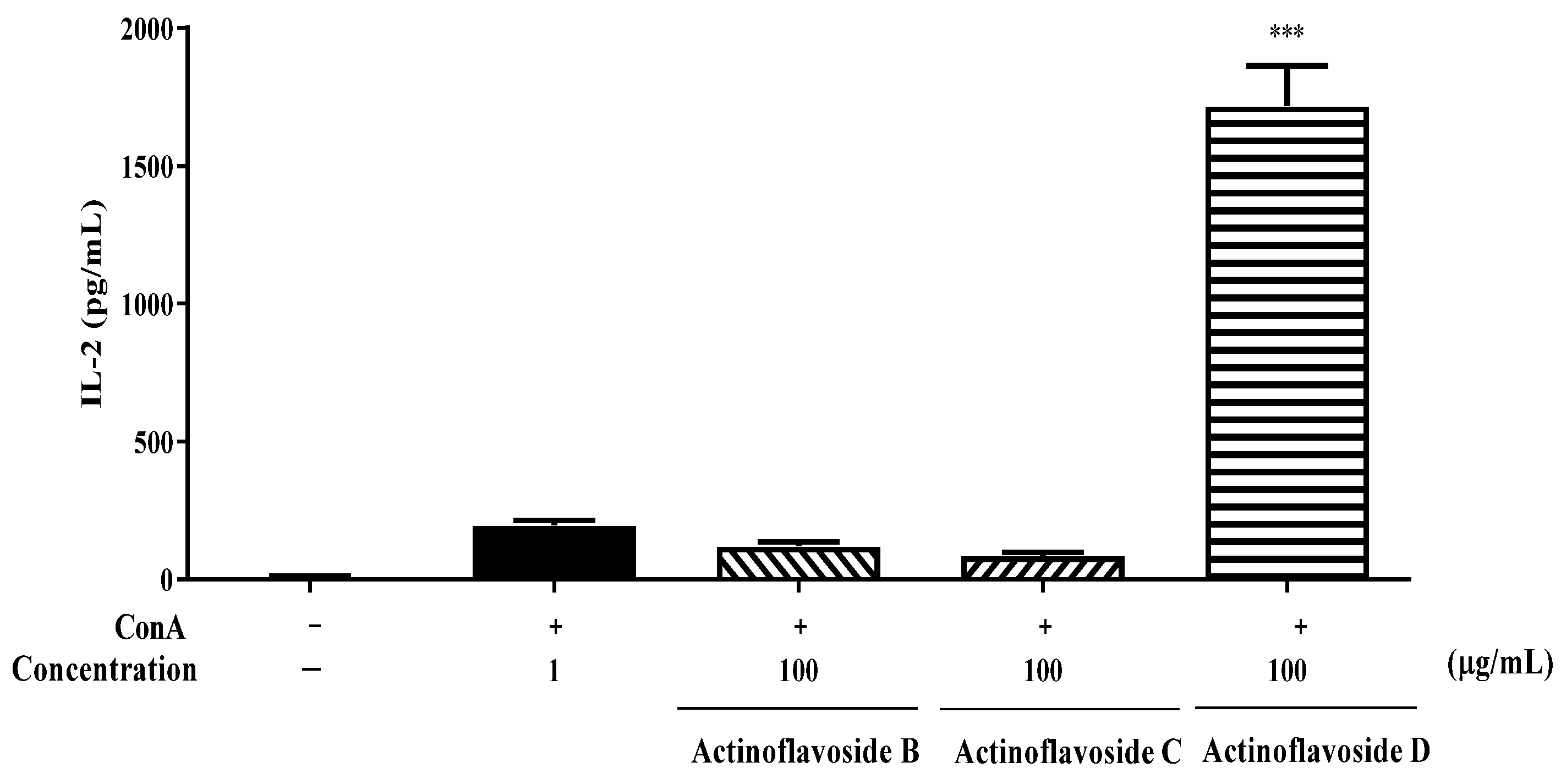
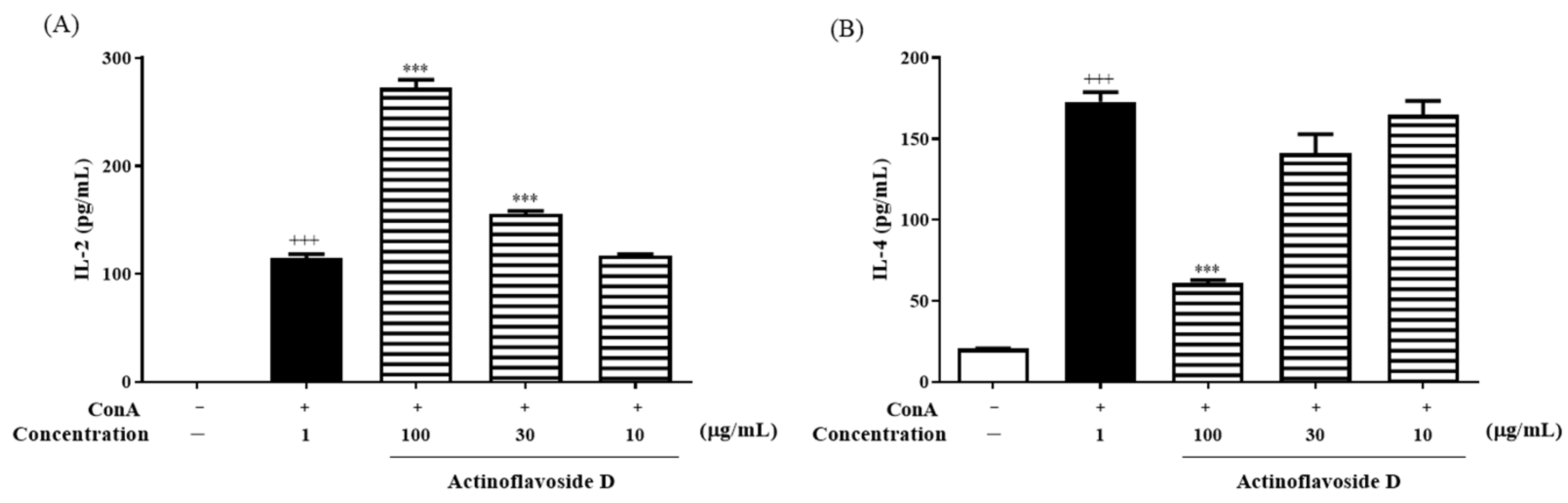
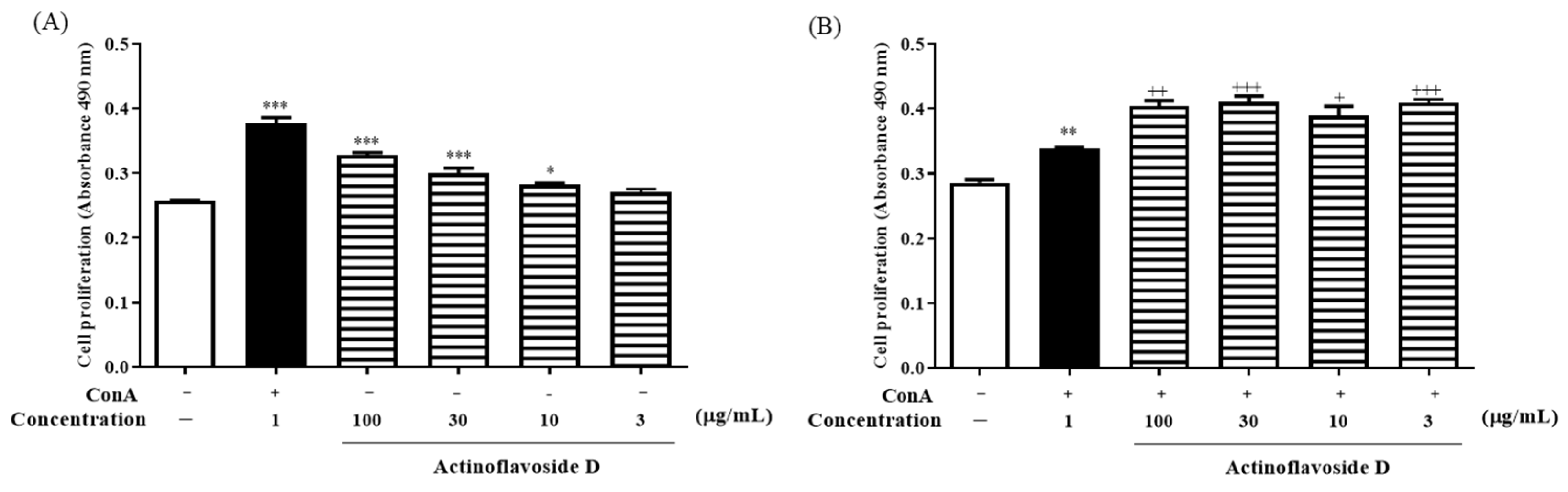
2.2. Biological Evaluation
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Bacterial Isolation
3.3. Cultivation and Extraction
3.4. Isolation of Actinoflavosides
3.5. MTPA Esterification of Actinoflavosides B and D
3.6. Antibacterial Activity Assay
3.7. Immunomodulatory Activity Assay
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L.; Sanchez, S. Microbial drug discovery: 80 years of progress. J. Antibiot. 2009, 62, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Gade, I.S.; Chadeneau, C.; Richard, T.S.; Atchade, A.D.; Talla, E.; Seite, P.; Vannier, B.; Guillard, J.M.; Laurent, S.; Henoumont, C.; et al. A new flavonoid glycoside from Tapinanthus sp. (Loranthaceae) and evaluation of anticancer activity of extract and some isolated compounds. Nat. Prod. Res. 2021, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Oomah, B.D.; Mazza, G. Flavonoids and antioxidative activities in buckwheat. J. Agric. Food Chem. 1996, 44, 1746–1750. [Google Scholar] [CrossRef]
- Lee, S.R.; Schalk, F.; Schwitalla, J.W.; Benndorf, R.; Vollmers, J.; Kaster, A.K.; de Beer, Z.W.; Park, M.; Ahn, M.J.; Jung, W.H.; et al. Polyhalogenation of Isoflavonoids by the Termite-Associated Actinomadura sp. RB99. J. Nat. Prod. 2020, 83, 3102–3110. [Google Scholar]
- Middleton, E. Effect of plant flavonoids on immune and inflammatory cell function. Adv. Exp. Med. Biol. 1998, 439, 175–182. [Google Scholar] [PubMed]
- Martins, B.T.; da Silva, M.C.; Pinto, M.; Cidade, H.; Kijjoa, A. Marine natural flavonoids: Chemistry and biological activities. Nat. Prod. Res. 2019, 33, 3260–3272. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Tsuruga, T.; Hiranuma, K.; Yamaura, M. Synthesis of 7,8-dihydroxy-5-hydroxymethyl-2-phenyl-chroman-4-one; the aglycon of actinoflavoside. Synlett 2004, 1, 116–118. [Google Scholar]
- Jiang, Z.D.; Jensen, P.R.; Fenical, W. Actinoflavoside, a novel flavonoid-like glycoside produced by a marine bacterium of the genus Streptomyces. Tetrahedron Lett. 1997, 38, 5065–5068. [Google Scholar] [CrossRef]
- Moon, K.; Chung, B.; Shin, Y.; Rheingold, A.L.; Moore, C.E.; Park, S.J.; Park, S.; Lee, S.K.; Oh, K.B.; Shin, J.; et al. Pentacyclic Antibiotics from a Tidal Mud Flat-Derived Actinomycete. J. Nat. Prod. 2015, 78, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Moon, K. Structure and Bioactivity of Boholamide A from a Tidal Mudflat Actinomycete. Korean J. Pharmacogn. 2021, 52, 203–207. [Google Scholar]
- Seo, J.; Shin, Y.-H.; Jo, S.J.; Du, Y.E.; Um, S.; Kim, Y.R.; Moon, K. Cystargamides C and D, New Cyclic Lipopeptides From a Tidal Mudflat-Derived Streptomyces sp. JMS132. Front. Microbiol. 2022, 13, 904954. [Google Scholar] [PubMed]
- Abbas, A.K.; Trotta, E.; Simeonov, D.R.; Marson, A.; Bluestone, J.A. Revisiting IL-2: Biology and therapeutic prospects. Sci. Immunol. 2018, 3, eaat1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly-Welch, A.; Hanson, E.M.; Keegan, A.D. Interleukin-4 (IL-4) Pathway. Sci. Stke 2005, 293, cm9. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.H.; Park, J.U.; Jo, S.J.; Ahn, J.H.; Park, J.H.; Yang, J.Y.; Lee, S.S.; Park, M.J.; Kim, Y.R. Anti-allergic Inflammatory Effects of the Essential Oil From Fruits of Zanthoxylum coreanum Nakai. Front. Pharmacol. 2018, 9, 1441. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Actinoflavoside B * | Actinoflavoside C * | Actinoflavoside D * | ||||
|---|---|---|---|---|---|---|
| Position | δC, Type | δH, Mult (J in Hz) | δC, Type | δH, Mult (J in Hz) | δC, Type | δH, Mult (J in Hz) |
| 2 | 78.6, CH | 5.61, m | 78.5, CH | 5.6, m | 75.8, CH | 4.51, m |
| 3 | 44.7, CH2 | 3.16, 2.79, m | 44.6, CH2 | 3.15, 2.81, m | 43.8 CH2 | 2.62, m |
| 4 | 192.3, C | 192.2, C | 191.0, C | |||
| 5 | 136.8, C | 136.9, C | 136.6, C | |||
| 6 | 106.5, CH | 7.17, s | 106.3, CH | 7.15, s | 106.6, CH | 7.10, s |
| 7 | 149.1, C | 149.1, C | 148.8, C | |||
| 8 | 133.6, C | 133.5, C | 133.6, C | |||
| 9 | 151.5, C | 151.6, C | 151.0, C | |||
| 10 | 112.9, C | 112.8, C | 113.0, C | |||
| 11 | 61.7, CH2 | 4.78, s | 61.6, CH2 | 4.77, s | 61.6, CH2 | 4.74, s |
| 1’ | 139.1, C | 139.2, C | 43.1, CH2 | 1.79, 1.47, m | ||
| 2’, 6’ | 126.7, CH | 7.59, m | 126.7, CH | 7.58, m | 23.5, CH | 1.94, m |
| 3’, 5’ | 128.4, CH | 7.38, m | 128.3, CH | 7.38, m | (2’ in 3)22.3, CH3(3’ in 3) | 0.92, m |
| 4’ | 128.5, CH | 7.43, m | 128.4, CH | 7.44, m | 22.8, CH3 | 0.92, m |
| 1’’ | 94.5, CH | 5.80, t (3.0) | 94.3, CH | 5.80, m | 94.8, CH | 5.77, t (3.0) |
| 2’’ | 32.9, CH2 | 2.08, 1.99, m | 32.7, CH2 | 2.03, 1.90, m | 32.9, CH2 | 2.07, 1.98, m |
| 3 | 44.8, CH | 4.34, dt (8.5, 4.0) | 45.4, CH | 4.29, dt (8.5, 4.0) | 44.8, CH | 4.33, m |
| 4’’ | 70.4, CH | 3.32, dd (9.0, 4.0) | 70.6, CH | 3.30, dd (9.0, 4.0) | 70.3, CH | 3.31, m |
| 5 | 66.4, CH | 3.79, m | 66.4, CH | 3.76, m | 66.5, CH | 3.81, m |
| 6’’ | 17.8, CH3 | 1.09, d (6.0) | 17.9 CH3 | 1.07, d (6.0) | 17.8, CH3 | 1.07, overlapped |
| NH | 7.61, m | 7.85, m | 7.6, m | |||
| 1’’’ | 174.8, C | 169.6, C | 171.0, C | |||
| 2’’’ | 47.9, CH | 2.28, m | 23.2, CH3 | 1.91, s | 47.3, CH | 2.30, m |
| 3’’’ | 68.3, CH | 3.61, dq (12.5, 6.0) | 68.3, CH | 3.63, dq (12.5, 6.0) | ||
| 4’’’ | 21.2, CH3 | 1.03, d (6.0) | 21.0, CH3 | 1.02, d (6.0) | ||
| 5’’’ | 14.2, CH3 | 1.06, d (7.0) | 14.2, CH3 | 1.08, overlapped | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.; Jo, S.J.; Bae, M.; Kim, Y.R.; Moon, K. Actinoflavosides B–D, Flavonoid Type Glycosides from Tidal Mudflat-Derived Actinomyces. Mar. Drugs 2022, 20, 565. https://doi.org/10.3390/md20090565
Jeong H, Jo SJ, Bae M, Kim YR, Moon K. Actinoflavosides B–D, Flavonoid Type Glycosides from Tidal Mudflat-Derived Actinomyces. Marine Drugs. 2022; 20(9):565. https://doi.org/10.3390/md20090565
Chicago/Turabian StyleJeong, Hyeongju, Se Jin Jo, Munhyung Bae, Young Ran Kim, and Kyuho Moon. 2022. "Actinoflavosides B–D, Flavonoid Type Glycosides from Tidal Mudflat-Derived Actinomyces" Marine Drugs 20, no. 9: 565. https://doi.org/10.3390/md20090565
APA StyleJeong, H., Jo, S. J., Bae, M., Kim, Y. R., & Moon, K. (2022). Actinoflavosides B–D, Flavonoid Type Glycosides from Tidal Mudflat-Derived Actinomyces. Marine Drugs, 20(9), 565. https://doi.org/10.3390/md20090565