

In Silico Identification and Molecular Mechanism of Novel Tyrosinase Inhibitory Peptides Derived from Nacre of Pinctada martensii


 , ,
, ,
Abstract

1. Introduction
2. Results and Discussion
2.1. Inhibition of Tyrosinase and Antioxidant Activity of the Enzymatic Hydrolysis Product
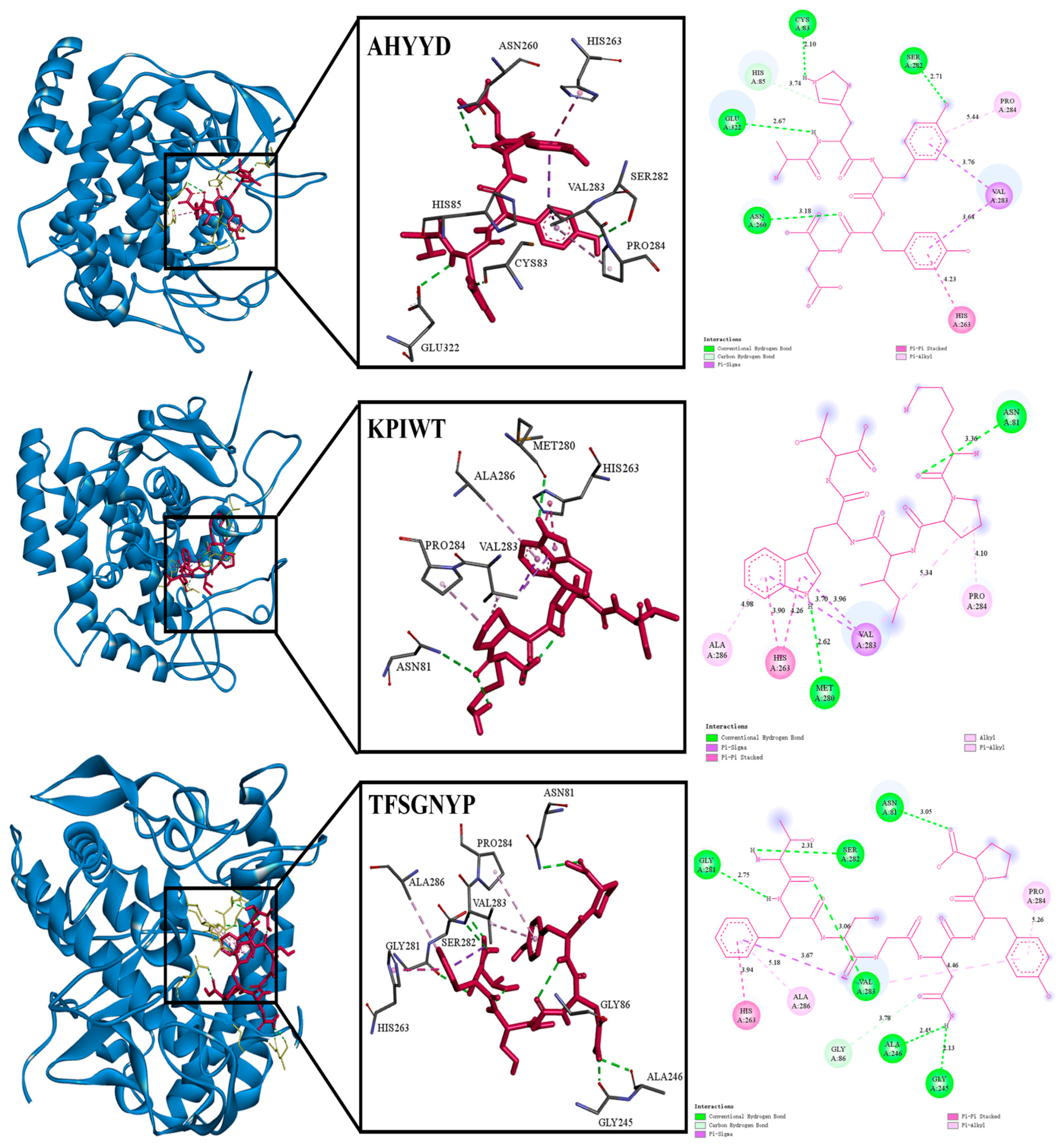
2.2. Identification and Molecular Docking
2.3. Molecular Dynamics Simulation
2.4. Tyrosinase-Inhibitory Activity and Antioxidant Capacity of Synthetic Peptides
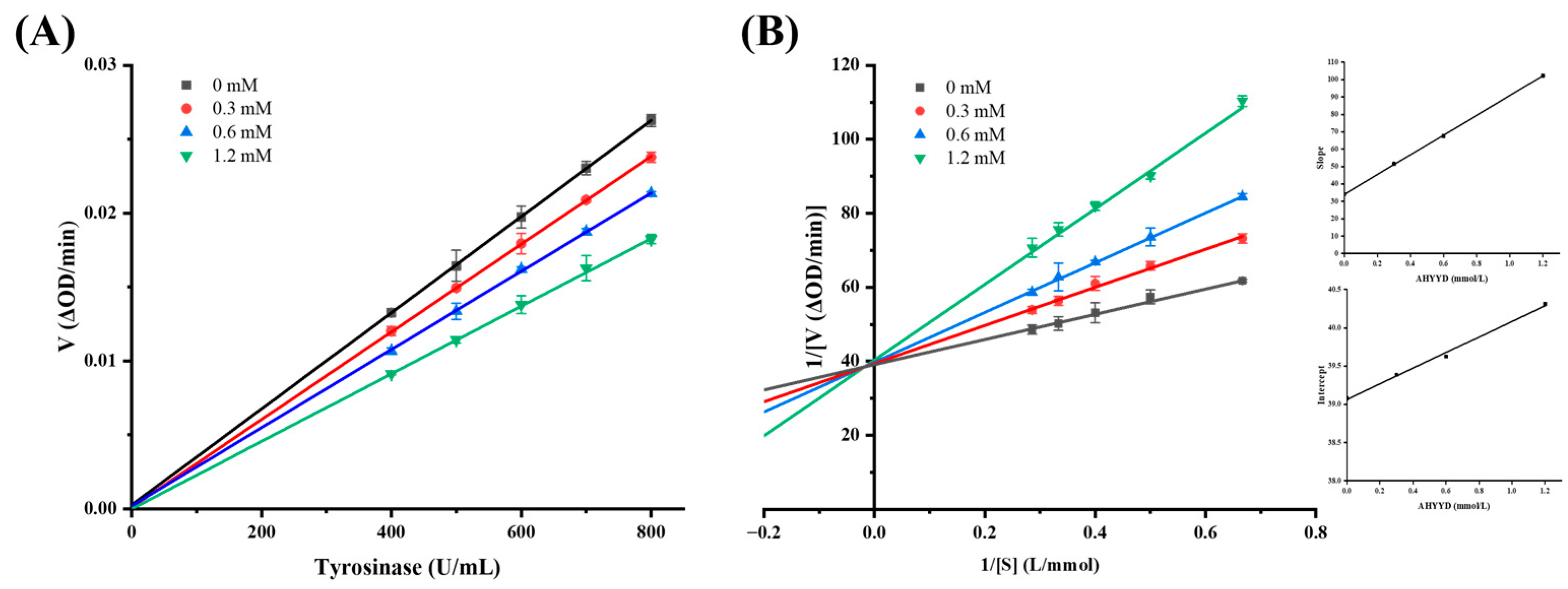
2.5. Inhibition Dynamics Analysis
2.6. CD Spectra Analysis
2.7. Isothermal Titration Calorimetry Analysis
2.8. Study on the Whitening Effect of Peptides Based on B16F10 Cells
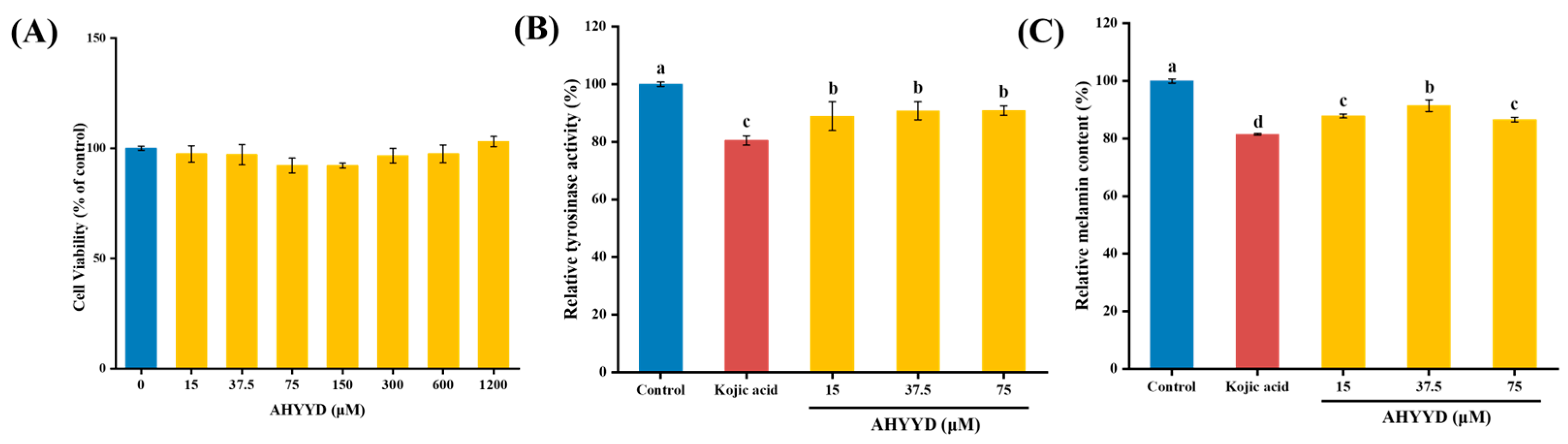
2.8.1. Cell Viability
2.8.2. Effect of AHYYD on the Tyrosinase Activity and Melanin Content of B16F10 Cells
2.8.3. Effect of AHYYD on the Antioxidant Enzyme Activity and ROS Content of B16F10 Cells
3. Materials and Methods
3.1. Materials
3.2. Preparation of Enzymatic Digests of Nacre Peptides
3.3. Tyrosinase-Inhibitory Activity and Antioxidant Activity Assay of Enzyme Digests
3.4. Protein Sequence Identification
3.5. Molecular Docking Studies
3.6. Molecular Dynamics Simulations
3.7. Peptides Synthesis
3.8. Determination of Inhibition Type
3.9. CD Measurements
3.10. Isothermal Titration Calorimetry Analysis
3.11. Cytotoxicity of AHYYD to B16F10 Cells
3.12. Effect of AHYYD on Melanin and Tyrosinase Synthesis in B16F10 Melanoma Cells
3.13. Effect of AHYYD on Antioxidant Enzyme Activity and ROS in B16F10 Cells
3.14. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Deng, Z.; Sun, J.; Wei, H.; Zhao, W.; Chen, M.; Li, Y.; Yu, G.; Wang, Y. Shell colors and microstructures of four pearl oyster species in the South China Sea. Aquac. Rep. 2022, 25, 101214. [Google Scholar] [CrossRef]
- Marie, B.; Joubert, C.; Tayale, A.; Zanella-Cleon, I.; Belliard, C.; Piquemal, D.; Cochennec-Laureau, N.; Marin, F.; Gueguen, Y.; Montagnani, C. Different secretory repertoires control the biomineralization processes of prism and nacre deposition of the pearl oyster shell. Proc. Natl. Acad. Sci. USA 2012, 109, 20986–20991. [Google Scholar] [CrossRef] [PubMed]
- Mariom; Take, S.; Igarashi, Y.; Yoshitake, K.; Asakawa, S.; Maeyama, K.; Nagai, K.; Watabe, S.; Kinoshita, S. Gene expression profiles at different stages for formation of pearl sac and pearl in the pearl oyster Pinctada fucata. BMC Genom. 2019, 20, 240. [Google Scholar] [CrossRef] [PubMed]
- Committee, C.P. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2020. [Google Scholar]
- Zhang, J.-X.; Li, S.-R.; Yao, S.; Bi, Q.-R.; Hou, J.-J.; Cai, L.-Y.; Han, S.-M.; Wu, W.-Y.; Guo, D.-A. Anticonvulsant and sedative-hypnotic activity screening of pearl and nacre (mother of pearl). J. Ethnopharmacol. 2016, 181, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Chen, S.; Li, L.; Zeng, Y.; Hu, X. The Hypopigmentation Mechanism of Tyrosinase Inhibitory Peptides Derived from Food Proteins: An Overview. Molecules 2022, 27, 2710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, Y.; Lai, R.; Deng, H.; Zhou, F.; Wang, P.; Pang, X.; Huang, G.; Chen, X.; Lin, H.; et al. Protective Effect of the Pearl Extract from Pinctada fucata martensii Dunker on UV-Induced Photoaging in Mice. Chem. Biodivers. 2022, 19, e202100876. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.-F.; Hsiao, S.-C.; Lu, Y.-Y.; Han, Y.-C.; Shen, Y.-C.; Venkatakrishnan, K.; Wang, C.-K. Efficacy of protein rich pearl powder on antioxidant status in a randomized placebo-controlled trial. J. Food Drug Anal. 2018, 26, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Loh, X.J.; Young, D.J.; Guo, H.; Tang, L.; Wu, Y.; Zhang, G.; Tang, C.; Ruan, H. Pearl Powder-An Emerging Material for Biomedical Applications: A Review. Materials 2021, 14, 2797. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.B.; Casadevall, A. Melanin. Curr. Biol. 2020, 30, R142–R143. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Sarna, T.; Plonka, P.M.; Raman, C.; Brozyna, A.A.; Slominski, A.T. Melanoma, Melanin, and Melanogenesis: The Yin and Yang Relationship. Front. Oncol. 2022, 12, 842496. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiong, B.; Xing, S.; Chen, Y.; Liao, Q.; Mo, J.; Chen, Y.; Li, Q.; Sun, H. Medicinal Prospects of Targeting Tyrosinase: A Feature Review. Curr. Med. Chem. 2023, 30, 2638–2671. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Wu, Z.; Zheng, R.; Yin, N.; Han, F.; Zhao, Z.; Dai, M.; Han, D.; Wang, W.; Niu, L. Inhibition mechanism of melanin formation based on antioxidant scavenging of reactive oxygen species. Analyst 2022, 147, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Qu, L.; Li, H.; He, J.; Wang, L.; Fang, Y.; Yan, X.; Yang, Q.; Peng, B.; Wu, W.; et al. Advances in Biomedical Functions of Natural Whitening Substances in the Treatment of Skin Pigmentation Diseases. Pharmaceutics 2022, 14, 3154. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, M.; Eslamifar, M.; Khezri, K. Kojic acid applications in cosmetic and pharmaceutical preparations. Biomed. Pharmacother. 2019, 110, 582–593. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Fan, M.; Yang, W.; Peng, Z.; Wang, G. Novel kojic acid-1,2,4-triazine hybrids as anti-tyrosinase agents: Synthesis, biological evaluation, mode of action, and anti-browning studies. Food Chem. 2023, 419, 136047. [Google Scholar] [CrossRef] [PubMed]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef] [PubMed]
- Zilles, J.C.; dos Santos, F.L.; Kulkamp-Guerreiro, I.C.; Contri, R.V. Biological activities and safety data of kojic acid and its derivatives: A review. Exp. Dermatol. 2022, 31, 1500–1521. [Google Scholar] [CrossRef] [PubMed]
- Masub, N.; Khachemoune, A. Cosmetic skin lightening use and side effects. J. Dermatol. Treat. 2022, 33, 1287–1292. [Google Scholar] [CrossRef]
- Hu, S.; Laughter, M.R.; Anderson, J.B.; Sadeghpour, M. Emerging topical therapies to treat pigmentary disorders: An evidence-based approach. J. Dermatol. Treat. 2022, 33, 1931–1937. [Google Scholar] [CrossRef] [PubMed]
- Reboucas, J.S.A.; Oliveira, F.P.S.; Araujo, A.C.D.S.; Gouveia, H.L.; Latorres, J.M.; Martins, V.G.; Prentice Hernandez, C.; Tesser, M.B. Shellfish industrial waste reuse. Crit. Rev. Biotechnol. 2023, 43, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wei, M.; You, L. Protective Effect of Peptides from Pinctada Martensii Meat on the H2O2-Induced Oxidative Injured HepG2 Cells. Antioxidants 2023, 12, 535. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Qiu, H.; Zhou, J.; Yang, C.; Chen, Y.; You, L. The Anti-Photoaging Activity of Peptides from Pinctada martensii Meat. Mar. Drugs 2022, 20, 770. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, C.; Tamura, S.; Tohse, R.; Fujita, S.; Kikuchi, M.; Asada, C.; Nakamura, Y. Isolation and identification of an angiotensin I-converting enzyme inhibitory peptide from pearl oyster (Pinctada fucata) shell protein hydrolysate. Process Biochem. 2019, 77, 137–142. [Google Scholar] [CrossRef]
- Kongsompong, S.; E-Kobon, T.; Taengphan, W.; Sangkhawasi, M.; Khongkow, M.; Chumnanpuen, P. Computer-Aided Virtual Screening and In Vitro Validation of Biomimetic Tyrosinase Inhibitory Peptides from Abalone Peptidome. Int. J. Mol. Sci. 2023, 24, 3154. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Miao, J.; Liao, W.; Huang, C.; Chen, B.; Li, Y.; Wang, X.; Yu, Y.; Liang, X.; Zhao, H.; et al. Rapid screening of novel tyrosinase inhibitory peptides from a pearl shell meat hydrolysate by molecular docking and the anti-melanin mechanism. Food Funct. 2023, 14, 1446–1458. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Wei, Z. Molecular simulation for food protein-ligand interactions: A comprehensive review on principles, current applications, and emerging trends. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13280. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guo, J.; Lian, J.; Gao, F.; Khan, A.J.; Wang, T.; Zhang, F. Molecular Simulation Study on the Interaction between Tyrosinase and Flavonoids from Sea Buckthorn. Acs Omega 2021, 6, 21579–21585. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. Software News and Update AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Skolnick, J.; Zhou, H. Implications of the Essential Role of Small Molecule Ligand Binding Pockets in Protein-Protein Interactions. J. Phys. Chem. B 2022, 126, 6853–6867. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lin, H.; Shen, W.; Qin, X.; Gao, J.; Cao, W.; Zheng, H.; Chen, Z.; Zhang, Z. Optimization of a Novel Tyrosinase Inhibitory Peptide from Atrina pectinata Mantle and Its Molecular Inhibitory Mechanism. Foods 2023, 12, 3884. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.-L.; Lim, G.T.; Yin, S.-J.; Lee, J.; Si, Y.-X.; Yang, J.-M.; Park, Y.-D.; Qian, G.-Y. The inhibitory effect of pyrogallol on tyrosinase activity and structure: Integration study of inhibition kinetics with molecular dynamics simulation. Int. J. Biol. Macromol. 2019, 121, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Tan, L.; Zhang, Q.; Chen, F.; Yu, Z. In silico identification and mechanistic evaluation of novel tyrosinase inhibitory peptides derived from coconut proteins. Food Biosci. 2024, 61, 104595. [Google Scholar] [CrossRef]
- Yu, Z.; Fu, L.; Zhang, Q.; Zhao, W. In silico identification and molecular mechanism of novel egg white-derived tyrosinase inhibitory peptides. Food Biosci. 2024, 57, 103567. [Google Scholar] [CrossRef]
- Najafi, Z.; Haramabadi, M.Z.; Chehardoli, G.; Ebadi, A.; Iraji, A. Design, synthesis, and molecular dynamics simulation studies of some novel kojic acid fused 2-amino-3-cyano-4H-pyran derivatives as tyrosinase inhibitors. BMC Chem. 2024, 18, 41. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-C.; Hsiao, N.-W.; Tseng, T.-S.; Chen, W.-C.; Lin, H.-H.; Leu, S.-J.; Yang, E.-W.; Tsai, K.-C. Phage Display-Mediated Discovery of Novel Tyrosinase-Targeting Tetrapeptide Inhibitors Reveals the Significance of N-Terminal Preference of Cysteine Residues and Their Functional Sulfur Atom. Mol. Pharmacol. 2015, 87, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; He, M.; Zhai, Y.; Xie, Z.; Xu, S.; Yu, S.; Xiao, H.; Song, Y. Inhibitory activity and mechanism of trilobatin on tyrosinase: Kinetics, interaction mechanism and molecular docking. Food Funct. 2021, 12, 2569–2579. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, M.; Murakami, K. A graphical method for determining inhibition constants. J. Enzym. Inhib. Med. Chem. 2009, 24, 1288–1290. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, L.; Mavridis, L.; Wallace, B.A.; Janes, R.W. DichroMatch at the protein circular dichroism data bank (DM@PCDDB): A web-based tool for identifying protein nearest neighbors using circular dichroism spectroscopy. Protein Sci. 2018, 27, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Paudyal, S.; Sharma, S.K.; da Silva, R.L.C.G.; Mintz, K.J.; Liyanage, P.Y.; Al-Youbi, A.O.; Bashammakh, A.S.; El-Shahawi, M.S.; Leblanc, R.M. Tyrosinase enzyme Langmuir monolayer: Surface chemistry and spectroscopic study. J. Colloid Interface Sci. 2020, 564, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Ott, F.; Rabe, K.S.; Niemeyer, C.M.; Gygli, G. Toward Reproducible Enzyme Modeling with Isothermal Titration Calorimetry. Acs Catal. 2021, 11, 10695–10704. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; He, X.; Teng, B.; McRae, J.M. Valonea Tannin: Tyrosinase Inhibition Activity, Structural Elucidation and Insights into the Inhibition Mechanism. Molecules 2021, 26, 2747. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Sha, X.; Zhang, L.; Huang, S.; Tu, Z. Effect of Grass Carp Scale Collagen Peptide FTGML on cAMP-PI3K/Akt and MAPK Signaling Pathways in B16F10 Melanoma Cells and Correlation between Anti-Melanin and Antioxidant Properties. Foods 2022, 11, 391. [Google Scholar] [CrossRef] [PubMed]
- Soares de Castro, R.J.; Sato, H.H. Biologically active peptides: Processes for their generation, purification and identification and applications as natural additives in the food and pharmaceutical industries. Food Res. Int. 2015, 74, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Van der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Yu, Q.; Fan, L.; Ding, Z. The inhibition mechanisms between asparagus polyphenols after hydrothermal treatment and tyrosinase: A circular dichroism spectrum, fluorescence, and molecular docking study. Food Biosci. 2022, 48, 101790. [Google Scholar] [CrossRef]
- Wang, W.; Gao, Y.; Wang, W.; Zhang, J.; Yin, J.; Le, T.; Xue, J.; Engelhardt, U.H.; Jiang, H. Kojic Acid Showed Consistent Inhibitory Activity on Tyrosinase from Mushroom and in Cultured B16F10 Cells Compared with Arbutins. Antioxidants 2022, 11, 502. [Google Scholar] [CrossRef]
- Zhang, X.; Li, J.; Li, Y.; Liu, Z.; Lin, Y.; Huang, J.-A. Anti-melanogenic effects of epigallocatechin-3-gallate (EGCG), epicatechin-3-gallate (ECG) and gallocatechin-3-gallate (GCG) via down-regulation of cAMP/CREB/MITF signaling pathway in B16F10 melanoma cells. Fitoterapia 2020, 145, 104634. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymatic Hydrolysis Product | Tyrosinase Inhibitory Activity (IC50, mg/mL) | DPPH Free Radical Scavenging (1 mg/mL, %) | ABTS Free Radical Scavenging (1 mg/mL, %) |
|---|---|---|---|
| NP-PMH | 6.743 ± 0.067 | 1.139 ± 0.232 | 85.050 ± 2.770 |
| NO. | Sequences | Length | Score | Toxicity | Affinity | Gravy |
|---|---|---|---|---|---|---|
| 1 | AHYYD | 5 | 27.12 | Non | −8.0 | −1.5 |
| 2 | GGFGNW | 6 | 30.94 | Non | −7.8 | −0.47 |
| 3 | KPIWT | 5 | 20.49 | Non | −7.4 | −0.52 |
| 4 | TFSGNYP | 7 | 21.74 | Non | −7.3 | −0.79 |
| 5 | ATFDAI | 6 | 27.86 | Non | −7.2 | 1.12 |
| 6 | NRIPN | 5 | 25.39 | Non | −7.2 | −1.72 |
| 7 | KRSLE | 5 | 21.85 | Non | −7.0 | −1.78 |
| 8 | HKDGY | 5 | 26.84 | Non | −6.9 | −2.46 |
| 9 | ERHLGY | 6 | 24.12 | Non | −6.9 | −1.52 |
| 10 | SIIDEVVA | 8 | 23.32 | Non | −6.9 | 1.43 |
| 11 | KDLFF | 5 | 20.31 | Non | −6.9 | 0.4 |
| 12 | GHSLTQF | 7 | 35.06 | Non | −6.8 | −0.29 |
| 13 | FGSLSF | 6 | 24.8 | Non | −6.8 | 1.23 |
| 14 | GGSFSVR | 7 | 26 | Non | −6.8 | 0.01 |
| 15 | TNNFT | 5 | 20.4 | Non | −6.7 | −1.12 |
| 16 | LPEEV | 5 | 23 | Non | −6.7 | −0.12 |
| 17 | SASTTLEE | 8 | 22.84 | Non | −6.6 | −0.55 |
| 18 | VTANPANT | 8 | 21 | Non | −6.6 | −0.28 |
| 19 | HSSAHS | 6 | 20.36 | Non | −6.5 | −1.17 |
| 20 | TNTSNP | 6 | 23.65 | Non | −6.5 | −1.8 |
| 21 | SDLGGI | 6 | 21.62 | Non | −6.5 | 0.53 |
| 22 | MVSLEG | 6 | 20.69 | Non | −6.5 | 0.87 |
| 23 | LKGHEDL | 7 | 20.01 | Non | −6.4 | −0.99 |
| 24 | SIDLYK | 6 | 20.7 | Non | −6.4 | −0.2 |
| 25 | MDLSHA | 6 | 21.73 | Non | −6.3 | 0 |
| 26 | DYQLP | 5 | 21.75 | Non | −6.2 | −1.22 |
| 27 | KEMQGG | 6 | 20.62 | Non | −6.2 | −1.63 |
| 28 | HTLESKPNPD | 10 | 22.99 | Non | −5.9 | −1.85 |
| 29 | KEPNK | 5 | 20.77 | Non | −5.9 | −3.28 |
| 30 | TDIIDG | 6 | 24.25 | Non | −5.8 | 0.15 |
| 31 | KKQLM | 5 | 20.39 | Non | −5.8 | −1.12 |
| 32 | MQVTPASA | 8 | 22.86 | Non | −5.8 | 0.39 |
| Peptides | Hydrogen Bonds | Hydrophobic Interaction | Electrostatic Interaction |
|---|---|---|---|
| AHYYD | Ser282,Cys83 Glu322,Asn260 | Val283,Pro284,His263 | |
| KPIWT | Met280,Asn81 | Ala286,His263Val283,Pro284 | |
| TFSGNYP | Asn81,Ser282,Gly281,Val283,Ala246,Gly245 | His263,Ala286,Pro284 |
| Samples | IC50 (mM) |
|---|---|
| AHYYD | 2.012 ± 0.088 |
| TFSGNYP | >5 |
| KPIWT | >10 |
| Kojic acid | 0.01 ± 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Lin, H.; Qin, X.; Gao, J.; Chen, Z.; Cao, W.; Zheng, H.; Xie, S. In Silico Identification and Molecular Mechanism of Novel Tyrosinase Inhibitory Peptides Derived from Nacre of Pinctada martensii. Mar. Drugs 2024, 22, 359. https://doi.org/10.3390/md22080359
Li F, Lin H, Qin X, Gao J, Chen Z, Cao W, Zheng H, Xie S. In Silico Identification and Molecular Mechanism of Novel Tyrosinase Inhibitory Peptides Derived from Nacre of Pinctada martensii. Marine Drugs. 2024; 22(8):359. https://doi.org/10.3390/md22080359
Chicago/Turabian StyleLi, Fei, Haisheng Lin, Xiaoming Qin, Jialong Gao, Zhongqin Chen, Wenhong Cao, Huina Zheng, and Shaohe Xie. 2024. "In Silico Identification and Molecular Mechanism of Novel Tyrosinase Inhibitory Peptides Derived from Nacre of Pinctada martensii" Marine Drugs 22, no. 8: 359. https://doi.org/10.3390/md22080359
APA StyleLi, F., Lin, H., Qin, X., Gao, J., Chen, Z., Cao, W., Zheng, H., & Xie, S. (2024). In Silico Identification and Molecular Mechanism of Novel Tyrosinase Inhibitory Peptides Derived from Nacre of Pinctada martensii. Marine Drugs, 22(8), 359. https://doi.org/10.3390/md22080359