Subjective Salience of Birdsong and Insect Song with Equal Sound Pressure Level and Loudness


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjective Salience Test
2.2. Physical Parameters
2.3. Multiple Regression Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Soeta, Y.; Kagawa, H. Subjective preferences for birdsong and insect song in equal sound pressure level. Appl. Sci. 2020, 10, 849. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, E.; Gatersleben, B.; Sowden, P.T. Bird sounds and their contributions to perceived attention restoration and stress recovery. J. Environ. Psychol. 2013, 38, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, E.; Gatersleben, B.; Sowden, P.T. Associations with bird sounds: How do they relate to perceived restorative potential? J. Environ. Psychol. 2016, 47, 136–144. [Google Scholar] [CrossRef]
- Axelsson, Ö.; Nilsson, M.E.; Hellström, B.; Lundén, P. A field experiment on the impact of sounds from a jet-and-basin fountain on soundscape quality in an urban park. Landsc. Urban Plan. 2014, 123, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Galbrun, L.; Calarco, F. Audio-visual interaction and perceptual assessment of water features used over road traffic noise. J. Acoust. Soc. Am. 2014, 136, 2609–2620. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Aletta, F.; Gjestland, T.T.; Brown, L.A.; Botteldooren, D.; Schulte-Fortkamp, B.; Lercher, P.; van Kamp, I.; Genuit, K.; Fiebig, A.; et al. Ten questions on the soundscapes of the built environment. Build. Environ. 2016, 108, 284–294. [Google Scholar] [CrossRef] [Green Version]
- De Coensel, B.; Vanwetswinkel, S.; Botteldooren, D. Effects of natural sounds on the perception of road traffic noise. J. Acoust. Soc. Am. 2011, 129, EL148–EL153. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.Y.; Lee, P.J.; You, J.; Kang, J. Perceptual assessment of quality of urban soundscapes with combined noise sources and water sounds. J. Acoust. Soc. Am. 2010, 127, 1357–1366. [Google Scholar] [CrossRef]
- Hong, J.Y.; Jeon, J.Y. Designing sound and visual components for enhancement of urban soundscapes. J. Acoust. Soc. Am. 2013, 134, 2026–2036. [Google Scholar] [CrossRef]
- Ou, D.; Mak, C.M.; Pan, S. A method for assessing soundscape in urban parks based on the service quality measurement models. Appl. Acoust. 2017, 127, 184–193. [Google Scholar] [CrossRef]
- Foundation for Promoting Personal Mobility and Ecological Transportation. Basic Case Study Report on Movement Support for Traffic Hub; Foundation for Promoting Personal Mobility and Ecological Transportation: Tokyo, Japan, 2009. (In Japanese) [Google Scholar]
- Ministry of Land, Infrastructure, Transport and Tourism. Guidelines for Improving the Facilitation of Transportation for Passenger Facilities of Public Transportation; Ministry of Land, Infrastructure, Transport and Tourism: Tokyo, Japan, 2020. (In Japanese)
- Sato, H.; Morimoto, M.; Sato, H. Effect of noise and reverberation on sound localization of acoustic guide signal for visually impaired persons in public spaces. Noise Control Eng. J. 2014, 62, 1–9. [Google Scholar] [CrossRef]
- Sato, H.; Morimoto, M.; Sato, H. Perception of azimuth angle of sound source located at high elevation angle: Effective distance of auditory guide signal. Appl. Acoust. 2020, 159, 107084. [Google Scholar] [CrossRef]
- Soeta, Y.; Nakagawa, S. Prediction of optimal auditory signals using auditory evoked magnetic responses. Build. Environ. 2015, 94, 924–929. [Google Scholar] [CrossRef]
- Huang, N.; Elhilali, M. Auditory salience using natural soundscapes. J. Acoust. Soc. Am. 2017, 141, 2163–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Lin, K.; Walther, D.B.; Hasegawa-Johnson, M.A.; Huang, T.S. Automatic detection of auditory salience with optimized linear filters derived from human annotation. Pattern Recognit. Lett. 2014, 38, 78–85. [Google Scholar] [CrossRef]
- Takeshima, H.; Suzuki, Y.; Suzuki, Y.; Sone, T. Growth of the loudness of a tone burst with a duration up to 10 seconds. J. Acoust. Soc. Jpn. 1988, 9, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Glasberg, B.R.; Moore, B.C.J. A model of loudness applicable to time-varying sounds. J. Audio Eng. Soc. 2002, 50, 331–342. [Google Scholar]
- Soeta, Y.; Kagawa, H. Three dimensional psychological evaluation of aircraft noise and prediction by physical parameters. Build. Environ. 2020, 16, 106445. [Google Scholar] [CrossRef]
- Scheffé, H. An analysis of variance for paired comparisons. J. Am. Stat. Assoc. 1952, 147, 381–400. [Google Scholar]
- Sato, S. Statistical Method of Sensory Test; JUSE Press: Tokyo, Japan, 1985. (In Japanese) [Google Scholar]
- Nagasawa, S. Improvement of the Scheffe’s method for paired comparisons. Kansei Eng. J. 2002, 3, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Ando, Y.; Cariani, P. Auditory and Visual Sensations; Springer: New York, NY, USA, 2009. [Google Scholar]
- Soeta, Y.; Ando, Y. Neurally Based Measurement and Evaluation of Environmental Noise; Springer: Tokyo, Japan, 2015. [Google Scholar]
- Ando, Y.; Kurihara, Y. Nonlinear response in evaluating the subjective diffuseness of sound field. J. Acoust. Soc. Am. 1986, 80, 833–836. [Google Scholar] [CrossRef]
- Sato, S.; Ando, Y. Apparent source width (ASW) of complex noises in relation to the interaural cross-correlation function. J. Temporal Des. Archit. Environ. 2002, 2, 29–32. [Google Scholar]
- Zwicker, E.; Fastl, H. Psychoacoustics: Facts and Models; Springer: Berlin, Germany, 1999. [Google Scholar]
- Giannakopoulos, T.; Pikrakis, A. Introduction to Audio Analysis: A MATLAB® Approach; Academic Press: Oxford, UK, 2008. [Google Scholar]
- Peeters, G. A large set of audio features for sound description (similarity and classification) in the CUIDADO. CUIDADO IST Proj. Rep. 2004, 54, 1–25. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Sato, S.; Kitamura, T.; Ando, Y. Annoyance of noise stimuli in relation to the spatial factors extracted from the interaural cross-correlation function. J. Sound Vib. 2004, 277, 511–521. [Google Scholar] [CrossRef]
- Sato, S.; You, J.; Jeon, J.Y. Sound quality characteristics of refrigerator noise in real living environments with relation to psychoacoustical and autocorrelation function parameters. J. Acoust. Soc. Am. 2007, 122, 314–325. [Google Scholar] [CrossRef]
- Jeon, J.Y.; Sato, S. Annoyance caused by heavyweight floor impact sounds in relation to the autocorrelation function and sound quality metrics. J. Sound Vib. 2008, 311, 767–785. [Google Scholar] [CrossRef]
- Soeta, Y.; Shimokura, R. Sound quality evaluation of air-conditioner noise based on factors of the autocorrelation function. Appl. Acoust. 2017, 124, 11–19. [Google Scholar] [CrossRef]
- McEvoy, L.K.; Picton, T.W.; Champagne, S.C. The timing of the processes underlying lateralization: Psychophysical and evoked potential measures. Ear Hear. 1991, 12, 389–398. [Google Scholar] [CrossRef]
- Zimmer, U.; Macaluso, E. High binaural coherence determines successful sound localization and increased activity in posterior auditory areas. Neuron 2005, 47, 893–905. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
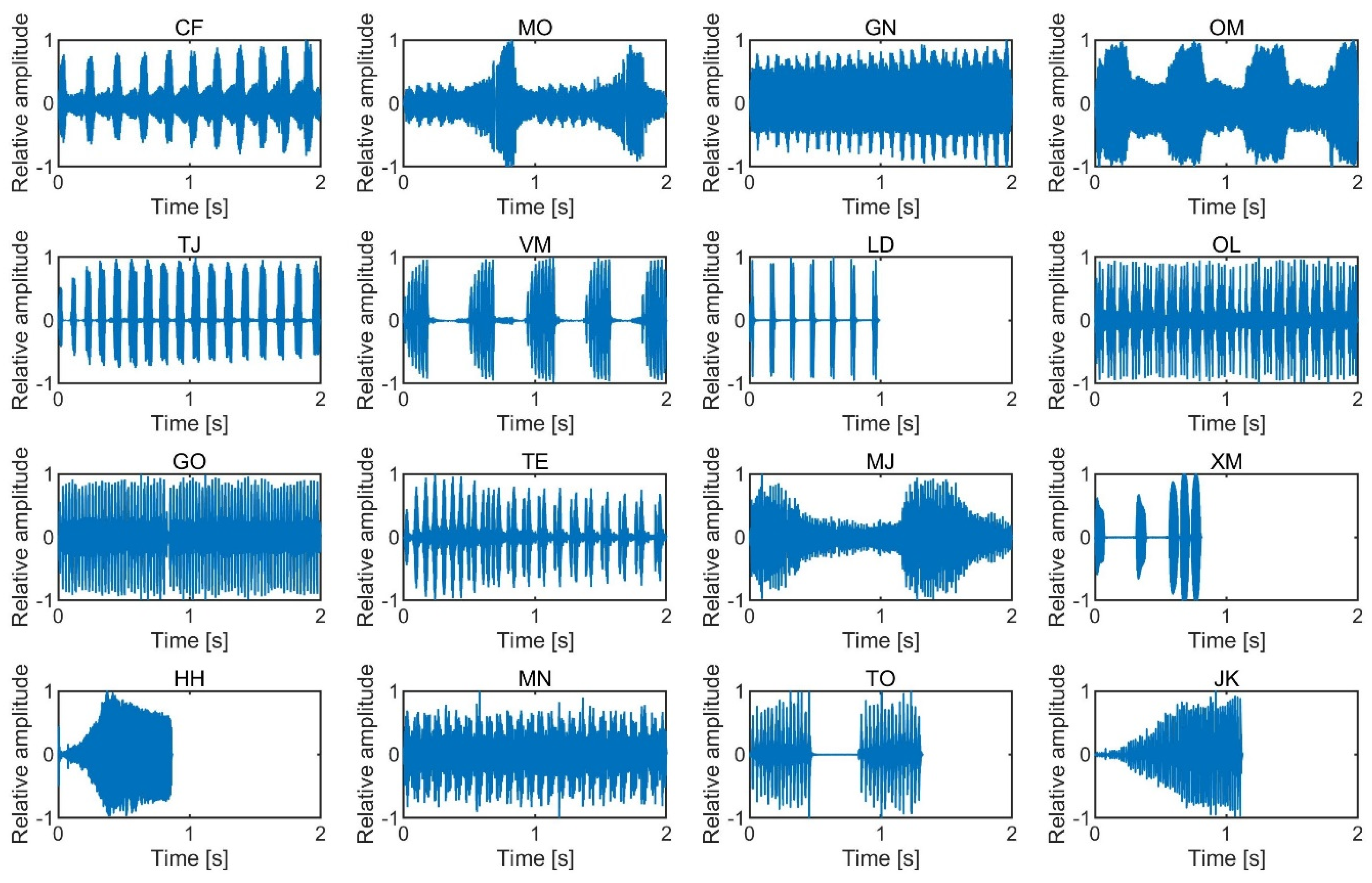{kind=link}
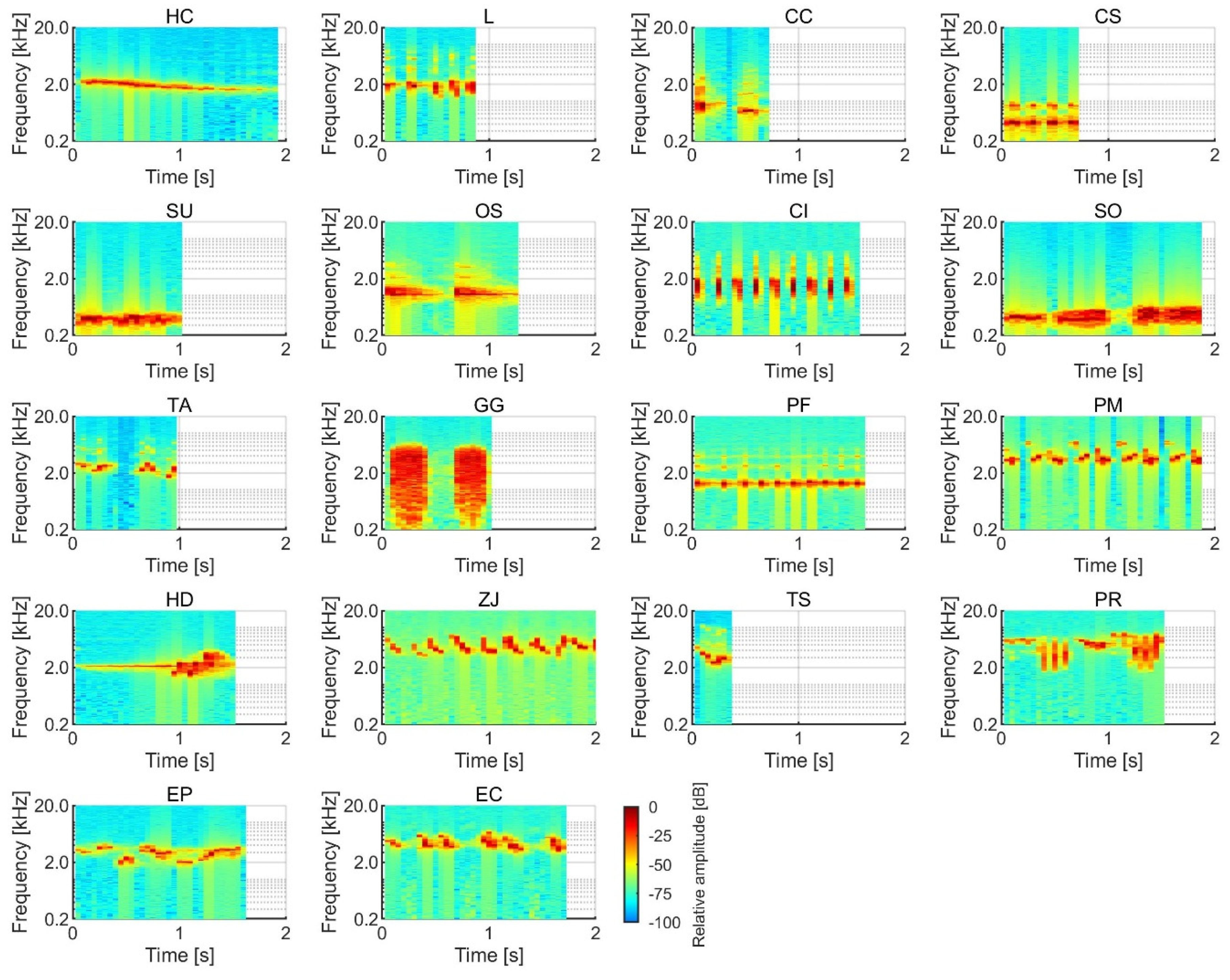{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entropy | Spectral Flux | Spectral Skewness | |
|---|---|---|---|
| IACC | 0.44 | −0.49 | 0.17 |
| Entropy | −0.31 | 0.43 | |
| Spectral flux | −0.49 |
| τε_QR | Wϕ(0)_QR | IACC | Loudness_QR | Roughness | |
|---|---|---|---|---|---|
| τ1 | 0.51 | 0.40 | 0.16 | 0.01 | −0.30 |
| τε_QR | −0.04 | 0.25 | 0.01 | −0.19 | |
| Wϕ(0)_QR | −0.61 | 0.22 | −0.54 | ||
| IACC | −0.35 | 0.43 | |||
| Loudness_QR | −0.22 |
| Predictive Parameter | Standardized Partial Regression Coefficients | |
|---|---|---|
| Equal LAeq | b1, IACC ** | 0.18 |
| b2, entropy * | −0.15 | |
| b3, spectral flux ** | 0.38 | |
| b4, spectral skewness ** | 0.79 | |
| Equal loudness | c1, τ1 ** | −0.57 |
| c2, τe_QR ** | 0.25 | |
| c3, Wϕ(0)_QR ** | 0.37 | |
| c4, IACC ** | 0.36 | |
| c5, loudness_QR ** | 0.49 | |
| c6, roughness ** | 0.17 |
| Loudness_QR | Fluctuation Strength | Spectral Entropy | Spectral Skewness | |
|---|---|---|---|---|
| Wϕ(0)_QR | 0.31 | 0.38 | 0.18 | 0.08 |
| Loudness_QR | −0.05 | 0.01 | −0.44 | |
| Fluctuation strength | 0.23 | 0.27 | ||
| Spectral entropy | 0.32 |
| τe | Roughness | Fluctuation Strength | Spectral Entropy_QR | |
|---|---|---|---|---|
| ϕ1 | 0.55 | 0.18 | 0.59 | −0.19 |
| τe | 0.10 | 0.46 | −0.20 | |
| Roughness | 0.03 | −0.11 | ||
| Fluctuation strength | 0.01 |
| Predictive Parameter | Standardized Partial Regression Coefficients | |
|---|---|---|
| Equal LAeq | i1, Wϕ(0)_QR * | 0.15 |
| i2, loudness_QR ** | −0.25 | |
| i3, fluctuation strength ** | 0.36 | |
| i4, spectral entropy ** | 0.44 | |
| i5, spectral skewness ** | 0.18 | |
| Equal loudness | j1, ϕ1 ** | −0.35 |
| j2, τe ** | 0.24 | |
| j3, roughness ** | −0.32 | |
| j4, fluctuation strength ** | 0.20 | |
| j5, spectral entropy_QR ** | −0.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soeta, Y.; Ariki, A. Subjective Salience of Birdsong and Insect Song with Equal Sound Pressure Level and Loudness. Int. J. Environ. Res. Public Health 2020, 17, 8858. https://doi.org/10.3390/ijerph17238858
Soeta Y, Ariki A. Subjective Salience of Birdsong and Insect Song with Equal Sound Pressure Level and Loudness. International Journal of Environmental Research and Public Health. 2020; 17(23):8858. https://doi.org/10.3390/ijerph17238858
Chicago/Turabian StyleSoeta, Yoshiharu, and Ayaka Ariki. 2020. "Subjective Salience of Birdsong and Insect Song with Equal Sound Pressure Level and Loudness" International Journal of Environmental Research and Public Health 17, no. 23: 8858. https://doi.org/10.3390/ijerph17238858
APA StyleSoeta, Y., & Ariki, A. (2020). Subjective Salience of Birdsong and Insect Song with Equal Sound Pressure Level and Loudness. International Journal of Environmental Research and Public Health, 17(23), 8858. https://doi.org/10.3390/ijerph17238858