
Prenatal Bisphenol a Exposure, DNA Methylation, and Low Birth Weight: A Pilot Study in Taiwan

 , ,
, ,
Abstract
:1. Introduction
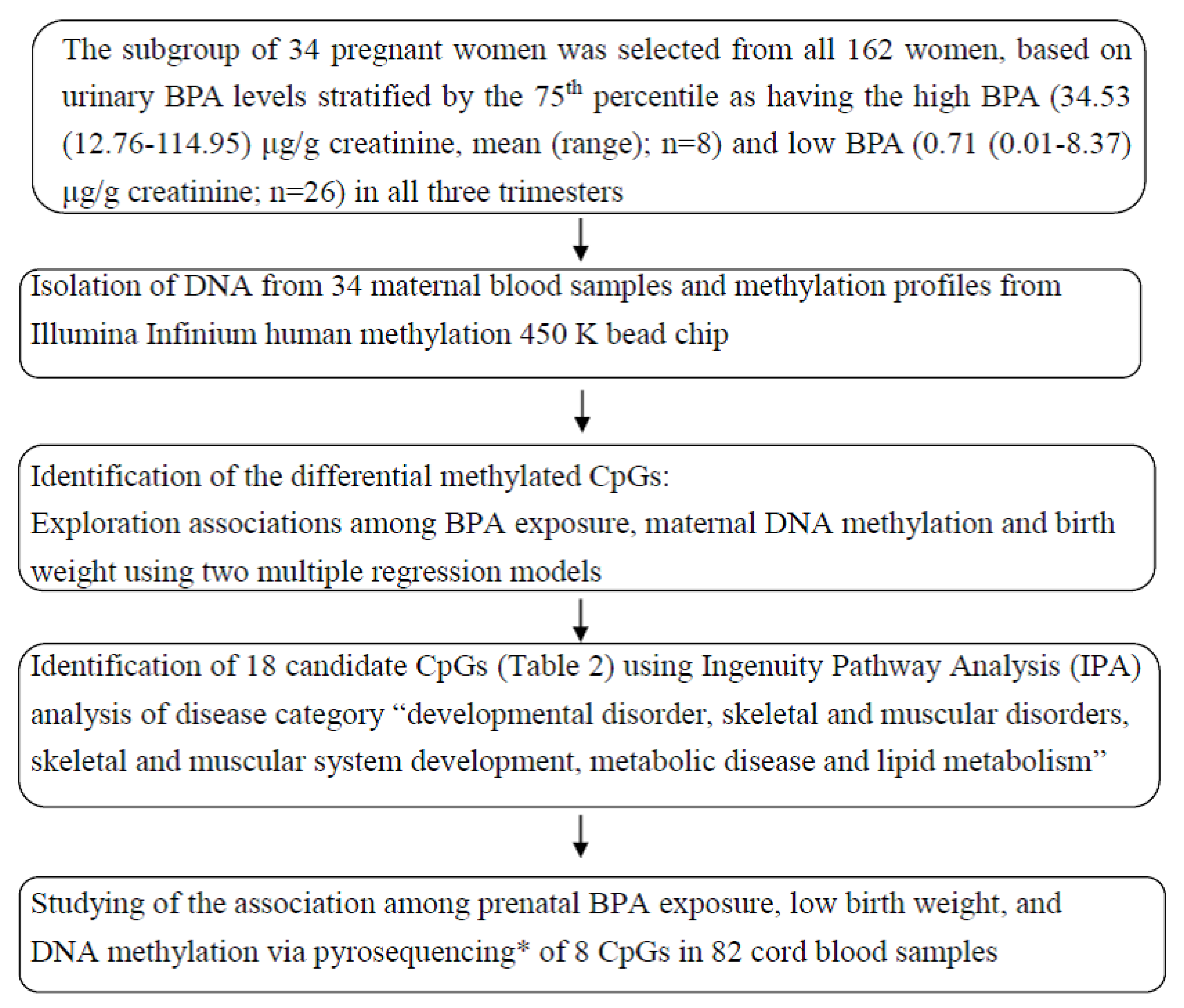
2. Material and Methods
2.1. Study Participants and Sample Collection
2.2. Measurement of BPA and Creatinine
2.3. Quantification of DNA Methylation and Normalization
2.4. Data Analysis
2.4.1. Screen and Selection of Candidate CpGs
Screen Candidate CpGs by Multivariate Linear Regression
Selection of Candidate Genes by Ingenuity Pathway Analysis (IPA)
2.5. Analysis of Methylation by Pyrosequencing and Validation in Cord Blood Samples
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of In Utero and Early-Life Conditions on Adult Health and Disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J.P.; Osmond, C.; Winter, P.D.; Margetts, B.; Simmonds, S.J. Weight in infancy and death from ischaemic heart disease. Lancet 1989, 334, 577–580. [Google Scholar] [CrossRef]
- Al-Gubory, K.H. Multiple exposures to environmental pollutants and oxidative stress: Is there a sex specific risk of developmental complications for fetuses? Birth Defects Res. Part C Embryo Today Rev. 2016, 108, 351–364. [Google Scholar]
- CDC, Centers for Disease Control and Prevention. Fourth National Reporton Human Exposure to Environmental Chemicals, 2017th ed.; CDC: Atlanta, GA, USA, 2014. [Google Scholar]
- Susiarjo, M.; Xin, F.; Bansal, A.; Stefaniak, M.; Li, C.; Simmons, R.A.; Bartolomei, M.S. Bisphenol A Exposure Disrupts Metabolic Health Across Multiple Generations in the Mouse. Endocrinology 2015, 156, 2049–2058. [Google Scholar] [CrossRef]
- Cheong, A.; Zhang, X.; Cheung, Y.Y.; Tang, W.Y.; Chen, J.; Ye, S.H.; Medvedovic, M.; Leung, Y.-K.; Prins, G.S.; Ho, S.-M. DNA methylome changes by estradiol benzoate and bisphenol A links early-life environmental exposures to prostate cancer risk. Epigenetics 2016, 11, 674–689. [Google Scholar] [CrossRef]
- Huang, Y.F.; Pan, W.C.; Tsai, Y.A.; Chang, C.H.; Chen, P.J.; Shao, Y.S.; Tsai, M.S.; Hou, J.-W.; Lu, C.; Chen, M.L. Concurrent exposures to nonylphenol, bisphenol A, phthalates, and organophosphate pesticides on birth outcomes: A cohort study in Taipei, Taiwan. Sci. Total Environ. 2017, 607–608, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Chou, W.C.; Chen, J.L.; Lin, C.F.; Chen, Y.C.; Shih, F.C.; Chuang, C.Y. Biomonitoring of bisphenol A concentrations in maternal and umbilical cord blood in regard to birth outcomes and adipokine expression: A birth cohort study in Taiwan. Environ. Health 2011, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Huo, W.; Xia, W.; Wan, Y.; Zhang, B.; Zhou, A.; Zhang, Y.; Huang, K.; Zhu, Y.; Wu, C.; Peng, Y. Maternal urinary bisphenol A levels and infant low birth weight: A nested case–control study of the Health Baby Cohort in China. Environ. Int. 2015, 85, 96–103. [Google Scholar] [CrossRef]
- Huang, Y.F.; Wang, P.W.; Huang, L.W.; Lai, C.H.; Yang, W.; Wu, K.Y.; Lu, C.; Chen, H.C.; Chen, M.L. Prenatal nonylphenol and bisphenol A exposures and inflammation are determinants of oxidative/nitrative stress: A Taiwanese cohort study. Environ. Sci. Technol. 2017, 51, 6422–6429. [Google Scholar] [CrossRef]
- Singh, S.; Li, S.S.L. Epigenetic Effects of Environmental Chemicals Bisphenol A and Phthalates. Int. J. Mol. Sci. 2012, 13, 10143–10153. [Google Scholar] [CrossRef]
- Hou, L.; Zhang, X.; Wang, D.; Baccarelli, A. Environmental chemical exposures and human epigenetics. Int. J. Epidemiol. 2012, 41, 79–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junge, K.M.; Leppert, B.; Jahreis, S.; Wissenbach, D.K.; Feltens, R.; Grützmann, K.; Thürmann, L.; Bauer, T.; Ishaque, N.; Schick, M. MEST mediates the impact of prenatal bisphenol A exposure on long-term body weight development. Clin. Epigenetics 2018, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Lee, Y.A.; Hong, Y.C.; Cho, J.; Lee, K.S.; Shin, C.H.; Kim, B.N.; Kim, J.I.; Park, S.J.; Bisgaard, H. Effect of prenatal bisphenol A exposure on early childhood body mass index through epigenetic influence on the insulin-like growth factor 2 receptor (IGF2R) gene. Environ. Int. 2020, 143, 105929. [Google Scholar] [CrossRef]
- Hanna, C.W.; Bloom, M.S.; Robinson, W.P.; Kim, D.; Parsons, P.J.; vom Saal, F.S.; Taylor, J.A.; Steuerwald, A.J.; Fujimoto, V.Y. DNA methylation changes in whole blood is associated with exposure to the environmental contaminants, mercury, lead, cadmium and bisphenol A, in women undergoing ovarian stimulation for IVF. Hum. Reprod. 2012, 27, 1401–1410. [Google Scholar] [CrossRef]
- Nahar, M.S.; Liao, C.; Kannan, K.; Harris, C.; Dolinoy, D.C. In utero bisphenol A concentration, metabolism, and global DNA methylation across matched placenta, kidney, and liver in the human fetus. Chemosphere 2015, 124, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Miura, R.; Araki, A.; Minatoya, M.; Miyake, K.; Chen, M.L.; Kobayashi, S.; Miyashita, C.; Yamamoto, J.; Matsumura, T.; Ishizuka, M.; et al. An epigenome-wide analysis of cord blood DNA methylation reveals sex-specific effect of exposure to bisphenol A. Sci. Rep. 2019, 9, 12369. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, M. Uber den niederschlag, welchen pikriksaure in normalen harn erzeugt und uber eine neue reaction des kreatinins. Z. Physiol. Chem. 1886, 10, 391. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Braun, J.M.; Smith, K.W.; Williams, P.L.; Calafat, A.M.; Berry, K.; Ehrlich, S.; Hauser, R. Variability of Urinary Phthalate Metabolite and Bisphenol A Concentrations before and during Pregnancy. Environ. Health Perspect. 2012, 120, 739–745. [Google Scholar] [CrossRef]
- Ye, X.; Pierik, F.H.; Hauser, R.; Duty, S.; Angerer, J.; Park, M.M.; Burdorf, A.; Hofman, A.; Jaddoe, V.W.V.; Mackenbach, J.P.; et al. Urinary metabolite concentrations of organophosphorous pesticides, bisphenol A, and phthalates among pregnant women in Rotterdam, the Netherlands: The Generation R study. Environ. Res. 2008, 108, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Arbuckle, T.E.; Weiss, L.; Fisher, M.; Hauser, R.; Dumas, P.; Berube, R.; Neisa, A.; LeBlanc, A.; Lang, C.; Ayotte, P.; et al. Maternal and infant exposure to environmental phenols as measured in multiple biological matrices. Sci. Total Environ. 2015, 508, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Casas, L.; Fernandez, M.F.; Llop, S.; Guxens, M.; Ballester, F.; Olea, N.; Irurzun, M.B.; Rodriguez, L.S.; Riano, I.; Tardon, A.; et al. Urinary concentrations of phthalates and phenols in a population of Spanish pregnant women and children. Environ. Int. 2011, 37, 858–866. [Google Scholar] [CrossRef]
- Callan, A.C.; Hinwood, A.L.; Heffernan, A.; Eaglesham, G.; Mueller, J.; Odland, J.Ø. Urinary bisphenol A concentrations in pregnant women. Int. J. Hyg. Environ. Health 2013, 216, 641–644. [Google Scholar] [CrossRef]
- Philippat, C.; Mortamais, M.; Chevrier, C.; Petit, C.; Calafat, A.M.; Ye, X.; Silva, M.J.; Brambilla, C.; Pin, I.; Charles, M.-A. Exposure to phthalates and phenols during pregnancy and offspring size at birth. Environ. Health Perspect. 2012, 120, 464–470. [Google Scholar] [CrossRef]
- Lee, J.; Choi, K.; Park, J.; Moon, H.-B.; Choi, G.; Lee, J.J.; Suh, E.; Kim, H.-J.; Eun, S.-H.; Kim, G.-H. Bisphenol A distribution in serum, urine, placenta, breast milk, and umbilical cord serum in a birth panel of mother–neonate pairs. Sci. Total Environ. 2018, 626, 1494–1501. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; He, Y.; Zhu, H.; Huang, X.; Bai, X.; Kannan, K.; Zhang, T. Concentrations of bisphenol A and its alternatives in paired maternal–fetal urine, serum and amniotic fluid from an e-waste dismantling area in China. Environ. Int. 2020, 136, 105407. [Google Scholar] [CrossRef] [PubMed]
- Philippat, C.; Wolff, M.S.; Calafat, A.M.; Ye, X.; Bausell, R.; Meadows, M.; Stone, J.; Slama, R.; Engel, S.M. Prenatal exposure to environmental phenols: Concentrations in amniotic fluid and variability in urinary concentrations during pregnancy. Environ. Health Perspect. 2013, 121, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Snijder, C.A.; Heederik, D.; Pierik, F.H.; Hofman, A.; Jaddoe, V.W.; Koch, H.M.; Longnecker, M.P.; Burdorf, A. Fetal growth and prenatal exposure to bisphenol A: The generation R study. Environ. Health Perspect. 2013, 121, 393. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.E.; Park, H.; Hong, Y.C.; Ha, M.; Kim, Y.; Chang, N.; Kim, B.N.; Kim, Y.J.; Yu, S.D.; Ha, E.H. Prenatal bisphenol A and birth outcomes: MOCEH (Mothers and Children’s Environmental Health) study. Int. J. Hyg. Environ. Health 2014, 217, 328–334. [Google Scholar] [CrossRef]
- Wolff, M.S.; Engel, S.M.; Berkowitz, G.S.; Ye, X.; Silva, M.J.; Zhu, C.; Wetmur, J.; Calafat, A.M. Prenatal phenol and phthalate exposures and birth outcomes. Environ. Health Perspect. 2008, 116, 1092. [Google Scholar] [CrossRef]
- Xu, X.; Chiung, Y.M.; Lu, F.; Qiu, S.; Ji, M.; Huo, X. Associations of cadmium, bisphenol A and polychlorinated biphenyl co-exposure in utero with placental gene expression and neonatal outcomes. Reprod. Toxicol. 2015, 52, 62–70. [Google Scholar] [CrossRef]
- Casas, M.; Valvi, D.; Ballesteros-Gomez, A.; Gascon, M.; Fernández, M.F.; Garcia-Esteban, R.; Iñiguez, C.; Martínez, D.; Murcia, M.; Monfort, N.; et al. Exposure to Bisphenol A and Phthalates during Pregnancy and Ultrasound Measures of Fetal Growth in the INMA-Sabadell Cohort. Environ. Health Perspect. 2016, 124, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.; Susiarjo, M.; Bartolomei, M.S. Multigenerational and transgenerational effects of endocrine disrupting chemicals: A role for altered epigenetic regulation? Semin. Cell Dev. Biology 2015, 43, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schübeler, D. Function and information content of DNA methylation. Nature 2015, 517, 321. [Google Scholar] [CrossRef]
- Reik, W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nature 2007, 447, 425. [Google Scholar] [CrossRef] [PubMed]
- Pilsner, J.R.; Hu, H.; Ettinger, A.; Sánchez, B.N.; Wright, R.O.; Cantonwine, D.; Lazarus, A.; Lamadrid-Figueroa, H.; Mercado-García, A.; Téllez-Rojo, M.M. Influence of prenatal lead exposure on genomic methylation of cord blood DNA. Environ. Health Perspect. 2009, 117, 1466. [Google Scholar] [CrossRef] [PubMed]
- LaRocca, J.; Binder, A.M.; McElrath, T.F.; Michels, K.B. The impact of first trimester phthalate and phenol exposure on IGF2/H19 genomic imprinting and birth outcomes. Environ. Res. 2014, 133, 396–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, E.J.; You, Y.A.; Park, B.; Ha, E.H.; Kim, H.S.; Park, H.; Kim, Y.J. Association between the DNA methylations of POMC, MC4R, and HNF4A and metabolic profiles in the blood of children aged 7–9 years. BMC Pediatr. 2018, 18, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, E.J.; Lee, H.A.; You, Y.A.; Park, H.; Cho, S.J.; Ha, E.H.; Kim, Y.J. DNA methylations of MC4R and HNF4α are associated with increased triglyceride levels in cord blood of preterm infants. Medicine 2016, 95, e4590. [Google Scholar] [CrossRef]
- Dolinoy, D.C.; Huang, D.; Jirtle, R.L. Maternal nutrient supplementation counteracts bisphenol A-induced DNA hypomethylation in early development. Proc. Natl. Acad. Sci. USA 2007, 104, 13056–13061. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.F.; Karmaus, W.J.J.; Yang, C.C.; Chen, M.L.; Wang, I.J. Bisphenol a Exposure, DNA Methylation, and Asthma in Children. Int. J. Environ. Res. Public Health 2020, 17, 298. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chen, J.; Wang, X.; Song, Q.; Xu, H.H.; Zhang, Y.H. Third trimester phthalate exposure is associated with DNA methylation of growth-related genes in human placenta. Sci. Rep. 2016, 6, 33449. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.E.; Meyer, N.; Santamaria, C.G.; Schumacher, A.; Luque, E.H.; Zenclussen, M.L.; Zenclussen, A.C. Bisphenol A exposure during early pregnancy impairs uterine spiral artery remodeling and provokes intrauterine growth restriction in mice. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Ye, Y.; Tang, Y.; Xiong, Y.; Feng, L.; Li, X. Bisphenol A exposure alters placentation and causes preeclampsia-like features in pregnant mice involved in reprogramming of DNA methylation of WNT2. FASEB J. 2019, 33, 2732–2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Dai, Y.; Jiang, X.; Liu, Y.; Chen, H.; Han, F.; Liu, J. Role of DNA methylation in bisphenol A exposed mouse spermatocyte. Environ. Toxicol. Pharmacol. 2016, 48, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Susiarjo, M.; Sasson, I.; Mesaros, C.; Bartolomei, M.S. Bisphenol a exposure disrupts genomic imprinting in the mouse. PLoS Genet. 2013, 9, e1003401. [Google Scholar] [CrossRef] [Green Version]
- Guida, M.; Troisi, J.; Ciccone, C.; Granozio, G.; Cosimato, C.; Sardo, A.D.S.; Di Carlo, C. Bisphenol A and congenital developmental defects in humans. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2015, 774, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Gervin, K.; Page, C.M.; Aass, H.C.D.; Jansen, M.A.; Fjeldstad, H.E.; Andreassen, B.K.; Duijts, L.; van Meurs, J.B.; van Zelm, M.C.; Jaddoe, V.W. Cell type specific DNA methylation in cord blood: A 450K-reference data set and cell count-based validation of estimated cell type composition. Epigenetics 2016, 11, 690–698. [Google Scholar] [CrossRef]

{kind=link}
| Variables | Total Subjects (n = 162) | Subjects with Illumina DNA Methylation Levels (n = 34) | p-Value |
|---|---|---|---|
| Maternal age (year) | 32.2 (20–43) | 32.6 (22.7–38.1) | 0.90 |
| Pre-pregnancy BMI (Kg/m2) | 21.1 (15.4–34.1) | 20.9 (17.3–28.2) | 0.57 |
| Maternal weight gain (Kg) | 12.4 (8.6–16.2) | 12.6 (8.0–19.0) | 0.93 |
| Birth weight (g) | 3121 (2400–3880) | 2928 (2520–3880) | 0.15 |
| BPA levels (μg/g creatinine) | |||
| 1st trimester | 2.46 (0.01–46.5) | 2.64 (0.01–24.1) | 0.74 |
| 2nd trimester | 4.53 (0.01–85.6) | 6.14 (0.01–32.4) | 0.30 |
| 3rd trimester | 11.31 (0.02–153.0) | 17.23 (0.03–114.9) | 0.61 |
| BPA stratified by the 75th percentile * Low exposure group | |||
| 1st trimester (<1.4 μg/g creatinine) | 0.10 (0.01–1.3) | 0.71 (0.01–8.4) | 0.37 |
| 2nd trimester (<3.3 μg/g creatinine) | 0.51 (0.01–3.3) | 0.48 | |
| 3rd trimester (<5.0 μg/g creatinine | 0.31 (0.01–4.3) | 0.86 | |
| High exposure group | |||
| 1st trimester (>1.4 μg/g creatinine) | 9.72 (1.4–46.5) | 34.53 (12.8–114.9) | NA |
| 2nd trimester (>3.3 μg/g creatinine) | 18.07 (3.7–85.6) | NA | |
| 3rd trimester (>5.0 μg/g creatinine) | 41.6 (5.0–153.0) | 0.38 | |
| Probe ID | CHR | Arm | Gene | Feature | CpG Islands | No. of CpG | Gene Name |
|---|---|---|---|---|---|---|---|
| cg18854735 | 1 | p | GNB1 | TSS1500 | island | 6 | guanine nucleotide binding protein (G protein), beta polypeptide 1 |
| cg24529814 | 1 | p | PRDM16 | Body | island | 4 | PR domain containing 16 |
| cg23231974 | 1 | p | PLOD1 | 1stExon | island | 7 | procollagen-lysine, 2-oxoglutarate 5-dioxygenase 1 |
| cg10635194 | 1 | p | KCND3 | TSS200 | island | 6 | potassium channel, voltage gated Shal related subfamily D, member 3 |
| cg00251716 | 1 | q | SDCCAG8 | Body | open sea | 1 | serologically defined colon cancer antigen 8 |
| cg03706175 | 2 | p | EPCAM | Body | shore | 2 | epithelial cell adhesion molecule |
| cg27640254 | 2 | q | DHRS9 | TSS1500 | open sea | 2 | dehydrogenase/reductase (SDR family) member 9 |
| cg11543686 | 2 | q | SLC19A3 | Body | open sea | 2 | solute carrier family 19 (thiamine transporter), member 3 |
| cg05524038 | 5 | q | CSF1R | TSS1500 | open sea | 2 | colony stimulating factor 1 receptor |
| cg18031134 | 6 | p | HLA-G | Body | island | 5 | major histocompatibility complex, class I, G |
| cg19768311 | 6 | p | DST | 3’UTR | open sea | 1 | dystonin |
| cg01502353 | 6 | p | DST | Body | open sea | 1 | dystonin |
| cg23244463 | 8 | q | KCNB2 | Body | open sea | 4 | potassium channel, voltage gated Shal related subfamily D, member 2 |
| cg07349217 | 8 | q | TG | TSS1500 | open sea | 1 | thyroglobulin |
| cg19427642 | 10 | q | KCNMA1 | Body | open sea | 1 | potassium channel, calcium activated large conductance subfamily M alpha, member 1 |
| cg27420224 | 20 | q | HNF4A | TSS200 | open sea | 4 | hepatocyte nuclear factor 4, alpha |
| cg00636769 | 20 | q | GNASAS | Body | open sea | 1 | GNAS antisense RNA 1 |
| cg17527673 | 22 | q | SCARF2 | Body | island | 4 | scavenger receptor class F, member 2 |
| Probe ID | Gene | No. of CpG | Average DNA Methylation (%) |
|---|---|---|---|
| cg01502353 | DST | 1 | 88 |
| cg05524038 | CSF1R | 2 | CpG1st (90) CpG2nd (93) |
| 1st and 2nd CpG (91) | |||
| cg07349217 | TG | 1 | 96 |
| cg19427642 | KCNMA1 | 1 | 90 |
| cg19768311 | DST | 1 | 91 |
| cg23244463 | KCNB2 | 4 | CpG1st (93) CpG2nd (100) |
| CpG3rd (98) CpG4th (100) | |||
| Average of the above CG site (97) | |||
| cg27420224 | HNF4A | 4 | CpG1st (86) CpG2nd (71) |
| CpG3rd (58) CpG4th (77) | |||
| Average of the above CpG (79) | |||
| cg27640254 | DHRS9 | 2 | CpG1st (94) CpG2nd (82) |
| 1st and 2nd CpG (88) |
| Genes | DNA Methylation Levels at CpG Sites (Cutoff by Median) | Odds Ratio | 95% CI | p-Value |
|---|---|---|---|---|
| DST | cg01502353 (methylation >88% vs. <88%) | 1.06 | 0.99–1.14 | 0.09 |
| CSF1R | cg05524038 CpG2nd (methylation >93% vs. <93%) | 1.03 | 0.95–1.11 | 0.50 |
| cg05524038 1st and 2nd CpG sites (methylation >91% vs. <91%) | 0.97 | 0.89–1.04 | 0.37 | |
| TG | cg07349217 (methylation >96% vs. <96%) | 1.01 | 0.94–1.09 | 0.74 |
| DST | cg19768311 (methylation >91% vs. <91%) | 0.96 | 0.89–1.03 | 0.23 |
| KCNB2 | cg23244463 CpG3rd (methylation >98% vs. <98%) | 1.00 | 0.94–1.09 | 0.82 |
| cg23244463 1 to 4 CpGs (methylation >96.8% vs. <96.8%) | 1.00 | 0.94–1.08 | 0.86 | |
| HNF4A | cg27420224 CpG1st (methylation>86% vs. <86%) | 0.97 | 0.90–1.04 | 0.40 |
| cg27420224 1 to 4 CpGs (methylation >79% vs. <79%) | 1.01 | 0.94–1.09 | 0.74 | |
| DHRS9 | cg27640254 CpG2nd (methylation >82.5% vs. <82.5%) | 1.10 | 1.00–1.21 | 0.047 |
| cg27640254 1st and 2nd CpGs (methylation >88.5% vs. <88.5%) | 0.96 | 0.88–1.03 | 0.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.-F.; Chang, C.-H.; Chen, P.-J.; Lin, I.-H.; Tsai, Y.-A.; Chen, C.-F.; Wang, Y.-C.; Huang, W.-Y.; Tsai, M.-S.; Chen, M.-L. Prenatal Bisphenol a Exposure, DNA Methylation, and Low Birth Weight: A Pilot Study in Taiwan. Int. J. Environ. Res. Public Health 2021, 18, 6144. https://doi.org/10.3390/ijerph18116144
Huang Y-F, Chang C-H, Chen P-J, Lin I-H, Tsai Y-A, Chen C-F, Wang Y-C, Huang W-Y, Tsai M-S, Chen M-L. Prenatal Bisphenol a Exposure, DNA Methylation, and Low Birth Weight: A Pilot Study in Taiwan. International Journal of Environmental Research and Public Health. 2021; 18(11):6144. https://doi.org/10.3390/ijerph18116144
Chicago/Turabian StyleHuang, Yu-Fang, Chia-Huang Chang, Pei-Jung Chen, I-Hsuan Lin, Yen-An Tsai, Chian-Feng Chen, Yu-Chao Wang, Wei-Yun Huang, Ming-Song Tsai, and Mei-Lien Chen. 2021. "Prenatal Bisphenol a Exposure, DNA Methylation, and Low Birth Weight: A Pilot Study in Taiwan" International Journal of Environmental Research and Public Health 18, no. 11: 6144. https://doi.org/10.3390/ijerph18116144