Abstract
Consumers’ preference towards healthy and novel foods dictates the production of organic unfiltered bottled vinegar that still contains acetic acid bacteria. After ingesting vinegar, the bacteria come into close contact with the human microbiota, creating the possibility of horizontal gene transfer, including genetic determinants for antibiotic resistance. Due to the global spread of antimicrobial resistance (AMR), we analyzed the AMR of Acetobacter and Komagataeibacter species originating mainly from vinegars. Six antibiotics from different structural groups and mechanisms of action were selected for testing. The AMR was assessed with the disk diffusion method using various growth media. Although the number of resistant strains differed among the growth media, 97.4%, 74.4%, 56.4%, and 33.3% of strains were resistant to trimethoprim, erythromycin, ciprofloxacin, and chloramphenicol, respectively, on all three media. Moreover, 17.9% and 53.8% of all strains were resistant to four and three antibiotics of different antimicrobial classes, respectively. We then looked for antimicrobial resistance genes in the genome sequences of the reference strains. The most common genetic determinant potentially involved in AMR encodes an efflux pump. Since these genes pass through the gastrointestinal tract and may be transferred to human microbiota, further experiments are needed to analyze the probability of this scenario in more detail.
1. Introduction
Many genetic determinants for antimicrobial resistance (AMR) are widely spread among clinically relevant bacteria, mainly because of humans’ excessive or improper use of antibiotics and the very efficient gene exchange and adaptation mechanisms in bacteria [1]. The AMR genetic determinants are also circulating among bacteria in the natural environment and in starter cultures used for food production, through which they enter the food chain and humans. In this way, they come into close contact with human microbiota [2,3].
A group of bacteria used for the production of vinegar, kefir, certain types of beer, nanocellulose, kombucha tea, nata de coco, and others, are acetic acid bacteria in the family Acetobacteraceae [4,5]. Two of the predominant genera in these processes are Komagataeibacter and Acetobacter. They are used for production at the industrial level and household needs. The natural environment of these bacteria are fruits and flowers, where different sugars and acids are available as carbon sources for their growth and multiplication. Circulation in the environment may enable the acetic acid bacteria to gain AMR via different gene exchange mechanisms from environmental microbiota. From the natural environment, such as vineyards, orchards, and meadows, the acetic acid bacteria are brought to the industrial and home environments. In this way, the acetic acid bacteria that originate from the surface of apples are reaching bioprocesses for the initiation of ethanol oxidation during apple cider vinegar production [6]. So far, researchers have mainly analyzed the biotechnologically relevant characteristics of acetic acid bacteria, such as the acetic acid production rate, tolerance to acetic acid and ethanol, and tolerance to high temperatures [7], but very rarely their AMR.
Data on the AMR of acetic acid bacteria have been mentioned in papers describing certain species of the Acetobacter genus in association with human diseases. In 2007, an isolate identified as Acetobacter cibinongensis was obtained from a blood sample of a 40 year old man in France suffering from bronchitis [8]. The antimicrobial drug susceptibility pattern could not be validated owing to the lack of assay reproducibility. However, the patient responded to the broad-spectrum antimicrobial therapy. In 2016, an isolate from a catheter of a child in Germany suffering from bacteremia was described as Acetobacter indonesiensis and was multidrug-resistant, with inhibition zones in an agar diffusion test only for imipenem, meropenem, fosfomycin, and tigecycline [9]. The isolate from the tracheostomy suction fluid of a 51 year old woman after lung transplantation was highly drug-resistant. This was A. indonesiensis as well, but showed in vitro activity against aminoglycosides, tetracyclines, imipenem, and ceftriaxone [10]. Recently, Wu et al. [11] identified, in a metagenomic approach, a genus Acetobacter in human fecal samples as one of the main genera carrying the top 20 antibiotic resistance gene types. So far, there are no data on the AMR of Komagataeibacter species.
To get more information on the AMR among two well-known acetic acid bacteria genera, we analyzed the resistance to selected antibiotics on a model group of different Acetobacter and Komagataeibacter species originating mainly from vinegars. To deepen this information to the DNA level, we searched in the genome sequences of type species for potential genetic information of AMR.
2. Materials and Methods
2.1. Strains of Acetic Acid Bacteria
The list of acetic acid bacteria used in this study is presented in Table 1. The model group presents 39 taxonomically well-characterized strains from the genera Acetobacter and Komagataeibacter. Although Gluconacetobacter entanii taxonomically belongs to the genus Komagataeibacter, this species remained in the genus Gluconacetobacter due to the absence of a cultivable form of the type strain of Gluconacetobacter entanii from any of the international culture collections of microorganisms.

Table 1.
List of strains used in this study.
Most of the strains originated from vinegars and were produced in different countries. The novel isolates of acetic acid bacteria were identified at the species level by the well-established method of 16S-23S rRNA ITS gene sequence analysis and the comparison to reference strains, as described before [12,13]. The sequences were deposited into the GenBank database under the accession numbers MZ747098, MZ747100, MZ747099, MZ725321, OL703587, MZ725320, OL703586, MZ725322, OL703585, OL703589, MZ725319, OL703588, MZ735454, and MT423517 for the strains Acetobacter estunensis AV380, Acetobacter estunensis AV390, Acetobacter pasteurianus AV366, Acetobacter pasteurianus SI3123, Acetobacter pomorum AV440, Komagataeibacter melomenusus SI3083, Komagateibacter oboediens AV371, Komagataeibacter oboediens SI3053, Komagataeibacter pomaceti AV445, Komagataeibacter pomaceti AV446, Komagataeibacter pomaceti SI3133, Komagataeibacter saccharivorans AV378, Gluconacetobacter entanii SI2035, and Gluconacetobacter entanii AV429, respectively.
The strains were revitalized from −80 °C on RAE medium composed of glucose (40 g/L), peptone (10 g/L), yeast extract (10 g/L), citric acid (1.37 g/L), Na2HPO4 × 2H2O (3.38 g/L), 1 vol% of ethanol, 1 vol% of acetic acid, and agar (10 g/L) [14]. The plates were incubated at 92–96% relative air humidity for 3 days at 30 °C. Additionally, strains were grown on media with mannitol (MA) and glucose (GY) at 30 °C for 3 days. The MA medium contained D-mannitol (25 g/L), peptone (3 g/L), yeast extract (5 g/L), and agar (15 g/L) (medium number 13 from the Belgian Coordinated Collections of Microorganisms), while the GY medium contained D-glucose (50 g/L), yeast extract (5 g/L), and agar (15 g/L) (modified medium 404 from the Belgian Coordinated Collections of Microorganisms).
2.2. Analysis of Antimicrobial Resistance (AMR)
The disk diffusion method adapted from EUCAST guidelines was used [15]. After successful revitalization, the strains were precultured onto RAE, GY, and MA media and incubated for three days at 30 °C; plates with the RAE medium were incubated at high relative air humidity. Then, the biomass from each plate was harvested into saline (0.85% NaCl) and the turbidity was adjusted to the value of the McFarland standard of 0.5. The bacterial suspension thus prepared was spread evenly with a sterile cotton swab over the entire surface of each medium. The antibiotic disks were applied onto plates. The following commercial antibiotic disks (BioRad, Hercules, USA) were applied onto the media: GMN10 for 10 μg of gentamicin, AMP10 for 10 μg of ampicillin, CHL30 for 30 μg of chloramphenicol, CIP5 for 5 μg of ciprofloxacin, ERY15 for 15 μg for erythromycin, and TMP5 for 5 μg of trimethoprim. Antibiotic resistance was compared among the strains by measuring the diameter of the growth inhibition zone around disks after two days of incubation at 30 °C of the inoculated medium, and in the case of the RAE medium, at a high relative air humidity.
2.3. Bioinformatics
The presence of homologues associated with AMR in the genome sequences of the type strains of acetic acid bacteria listed in Table 1 was analyzed using the online tool Resistance Gene Identifier (RGI) from the Comprehensive Antibiotic Resistance Database (CARD) [16]. The genome sequences of the type strains were downloaded from the NCBI database, annotated by RGI, and compared to the AMR reference database in CARD. Since none of the genome sequences had a homologue with a perfect matching, the selection criteria were extended to loose hits. The AMR homologues matching in length to the reference hit from 95% to 105% were selected as putative homologues. Additionally, a minimum of 25% amino acid identity matching the reference hit was set as a second criterion.
3. Results and Discussion
Acetic acid bacteria are widespread microorganisms in nature. They have been used to produce different foods and beverages [13] and pharmaceutical and medical products [17]. They are generally recognized as safe bacteria, but antibiotic resistance has not been systematically investigated in acetic acid bacteria.
In this study we analyzed the resistance of taxonomically well-defined representatives of Acetobacter and Komagataeibacter species, originating from different geographic areas, against antibiotics of different chemical structures and mechanisms of action. We selected ampicillin as a representative of bacterial cell wall synthesis inhibitor, chloramphenicol, erythromycin, and gentamicin as representative of bacterial protein synthesis inhibitors, and ciprofloxacin and trimethoprim as representative of bacterial DNA synthesis inhibitors. All these antibiotics belong to different antimicrobial classes: ampicillin to penicillins, chloramphenicol to phenicols, erythromycin to macrolides, gentamicin to aminoglycosides, ciprofloxacin to fluoroquinolones, and trimethoprim to diaminopyrimidines.
Since there is no standard method and medium defined yet for testing the antibiotic resistance of acetic acid bacteria, we used three different media which enable the growth of acetic acid bacteria. The main differences among media were in major carbon sources and pH values. The RAE growth medium has glucose and ethanol as carbon sources and a pH value of 3.5, the MA growth medium has mannitol and a pH of 6.7, and the GY medium has glucose and a pH of 6.9. The area of inhibition for each antibiotic differed among the media used (Tables S1–S3). Different factors can explain differences in the growth inhibition zones on different media against the same antibiotic. For example, in the case of trimethoprim, lower pH in the medium causes the ionization of trimethoprim, affecting its ability to cross the cell membrane in E. coli [18]. It is also possible that the medium components, such as sugars and acids, may direct the cells to different metabolic pathways, resulting in metabolites that may interact with antibiotics [19]. Then, the cell components, of which the synthesis has been inhibited by antibiotics, may be replaced by a compound present in the medium. Therefore, the formulation of an optimal medium that effectively supports bacterial growth but does not adversely affect its mechanism of action is difficult, especially for bacteria with specific growth requirements, including acetic acid bacteria.
Many acetic acid bacteria strains did not produce growth inhibition zones around the antibiotic disks (Tables S1–S3). These data were combined in one table (Table 2) according to the criterion that the absence of an inhibition zone on all of the applied growth media represents a resistant strain to a particular antibiotic. In this way, 97.4%, 74.4%, 56.4%, and 33.3% were resistant to trimethoprim, erythromycin, ciprofloxacin, and chloramphenicol, respectively (Figure 1). Moreover, 53.8% of all strains were resistant to three or more antimicrobial classes [20], and thus represented multidrug-resistant strains. The most common profile of multiple resistance was trimethoprim/ciprofloxacin/erythromycin. Only a few strains were resistant to ampicillin or gentamicin; on all three media, only one strain was resistant towards ampicillin, and none against gentamicin (Figure 1). Separate figures for Acetobacter and Komagataeibacter species show that a substantial difference in resistance against chloramphenicol, ciprofloxacin, and erythromycin exists (Figure S1); however, this has to be proved with a higher number of tested strains from each genus in the future.
Table 2.
Resistance of Acetobacter and Komagataeibacter strains against antibiotics: gentamicin (GMN), ampicillin (AMP), chloramphenicol (CHL), ciprofloxacin (CIP), erythromycin (ERY) and trimethoprim (TMP). The X-label represents the growth arrest of a particular strain around the antibiotic disk on all three tested growth media.
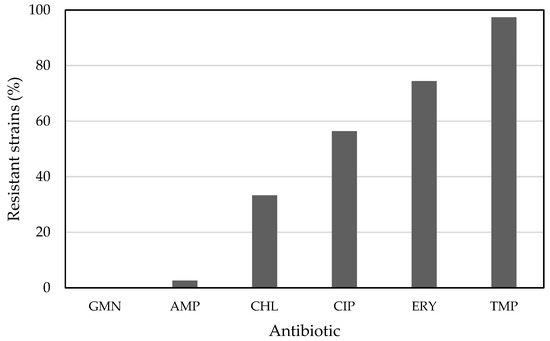
Figure 1.
Acetic acid bacteria-resistant strains to gentamicin (GMN), ampicillin (AMP), chloramphenicol (CHL), ciprofloxacin (CIP), erythromycin (ERY), and trimethoprim (TMP).
Of all tested strains and growth media, only the strain Komagataeibacter melaceti AV382T cultured on the RAE medium was sensitive to trimethoprim. Its growth inhibition zone (8–9 mm) was obvious and repeatedly detected (Table S1). Resistance to trimethoprim is caused by decreased cell permeability and modifications in the bacterial target for trimethoprim, i.e., dihydrofolate reductase [21]. Trimethoprim resistance can also originate from cross-resistance to other antibiotics [22]. Since most strains resistant to trimethoprim in this work also possess resistance to some other antibiotics (i.e., erythromycin, ciprofloxacin, and chloramphenicol), a general nonspecific mechanism, such as transport out of the cell, may be responsible for trimethoprim resistance. Although trimethoprim resistance has been detected in almost all strains, a trimethoprim-sensitive strain (Table 2) suggests that trimethoprim resistance is not an intrinsic characteristic for acetic acid bacteria.
Erythromycin resistance has been well-studied in another group of food-grade bacteria, lactic acid bacteria (LAB). A total of 97 out of 155 isolates of LAB from human feces were resistant to erythromycin, and 19 of them carried ermB, coding for erythromycin esterase. The gene was also successfully transferable to Enterococcus faecalis [23]. Bacteria have also developed other mechanisms to circumvent the action of macrolides, such as decreasing intracellular concentration via the use of efflux pumps, ribosome modification, ribosome protection, and macrolide phosphotransferase-mediated modification [23]. In the presented study, none of the strains of Acetobacter and Komagataeibacter resisted erythromycin on the RAE medium (Table S1), in contrast to the successful growth of some strains in the presence of erythromycin on the GY (Table S2) and MA media (Table S3). This suggests the instability of erythromycin at a low pH in the RAE medium; an observation also supported by the study of Brisaert, who showed that the stability of erythromycin was negatively affected by a low pH [24].
Ciprofloxacin is an ampholytic compound with a pKa value of 6.09 for the carboxylic group and 8.74 for the nitrogen on the piperazinyl ring [25]. Zwitterion structure formed at a pH of 7.4 and was susceptible to photodegradation. Maximal stability of ciprofloxacin was observed at a pH of 3.0 and 4.0 [26]. This may also explain why most of the analyzed strains of Acetobacter and Komagataeibacter were resistant against this drug in the RAE medium, of which the pH value was 3.5. Interestingly, the only exception to this was the strain Gluconacetobacter entanii AV429. A total of 66.7% and 64.1% of strains on the GY and MA media were ciprofloxacin-resistant. Ciprofloxacin targets bacterial topoisomerase II (DNA gyrase), which are involved in DNA replication and transcription [27]. Generally, the mutation in genes coding for gyrase (gyrA) is responsible for the resistant phenotype; however, other mechanisms, especially efflux pumps, may also prevent the intracellular action of ciprofloxacin [28].
Chloramphenicol sensitivity among strains of acetic acid bacteria has been observed on all three media, and also on the RAE medium with an acidic pH (Table S1), which is probably the result of chloramphenicol pH stability over a wide pH range [29]. However, only 13 strains, distributed among the Acetobacter and Komagataeibacter genera, resisted chloramphenicol on all three media (Table 2), suggesting that other factors may be involved in the susceptibility to this drug. Bacteria have developed different chloramphenicol resistance mechanisms, a very specific enzymatic inactivation of the drug by chloramphenicol acetyltransferase, but also efflux drug removal, ribosome protection, and the nitro reduction of chloramphenicol [30].
Ampicillin resistance was detected on each of the three media; however, only one strain exhibited resistance on all three media. The number of strains resistant to ampicillin on the RAE and GY media was the same, but not the same strains exhibited resistance on both media. The number of strains resistant to ampicillin was the lowest on the MA medium. The previous study showed the instability of ampicillin at a higher pH [31]; however, a high number of ampicillin resistance strains on the GY medium (pH 6.9) (Table S2) in contrast to the RAE medium (pH 3.5) (Table S1) suggests that other factors are involved in the ampicillin resistance of acetic acid bacteria on these media.
We detected a high difference in the number of strains resistant to gentamicin among the culture media. The number of strains resisting gentamicin on the GY and MA media was one and three, respectively, but on the RAE medium it was 22. Since gentamicin is stable at a pH of 2–14, the reason for these differences has to be in other factors. Gentamicin resistance is often associated with aminoglycoside-modifying enzymes such as acetyltransferases, phosphotransferases, and methyltransferases. Besides, target site modification and drug efflux may also be responsible for gentamicin resistance [32].
The antibiotic resistome prediction in the genome sequences of type strains revealed matching regions to known molecular determinants of antibiotic resistance (Table S4). These genetic determinants corresponded to five resistance mechanisms: antibiotic efflux, antibiotic target alteration, antibiotic inactivation, reduced permeability to the antibiotic, and antibiotic target replacement (Table 3). The proportion of individual mechanisms was similar in each genus (Table S5). The highest number of homologues, among them AcrA, MdtA, MexB, MuxB, OpmB, OprM, AmvA, BaeR, RanA, EmrE, QacL, TriC, MdsC, MexK, RosA, EmrB, FarA, MexL, RosB, TxR, SdiA, KdpE, MsbA, QacE, MexS, MtrA, and NmcR, were in the category antibiotic efflux (Table S4). These putative pumps may transport different antibiotics out of the cell. Further on, at least one homologue putatively encoding β-lactamase was detected for each of the analyzed reference strains. Since this is the most well-known mechanism for ampicillin deactivation [33], ampicillin resistance was expected for these strains and confirmed for each reference strain on at least one of the three growth media. In two reference strains, a genetic determinant putatively coding for chloramphenicol acetyltransferase was detected; both strains also exhibited resistance to chloramphenicol on at least one of the growth media. The correlation between the resistance of acetic acid bacteria to the specific antibiotic and genetic determinants responsible for it needs to be further elucidated by genetic studies in acetic acid bacteria.
Table 3.
Resistance mechanisms of putative antimicrobial resistance gene families identified in type strains of Acetobacter and Komagataeibacter species listed in Table 1.
4. Conclusions
This work is the first pilot systematic study on AMR in the genera Acetobacter and Komagataeibacter. Although the assessment of AMR among acetic acid bacteria is confounded due to the lack of standards for testing, these results have undoubtedly revealed widespread AMR among Acetobacter and Komagataeibacter species. This is just a first step in this research field that needs further studies to identify the exact genetic determinants and molecular mechanisms for AMR in acetic acid bacteria, but also to find out if this genetic information is transferable to human pathogens and to environmental bacteria, from where it can further spread to clinically relevant bacteria. It is also crucial that, in the future, a standardized method is established for the analysis of antibiotic resistance in this group of bacteria, or separately for each genus of acetic acid bacteria.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ijerph19010463/s1, Table S1: Antibiogram of Acetobacter and Komagataeibacter species on RAE medium. No inhibition zone is presented as /; Table S2: Antibiogram of Acetobacter and Komagataeibacter species on GY medium. No inhibition zone is presented as /; Table S3: Antibiogram of Acetobacter and Komagataeibacter species on MA medium. No inhibition zone is presented as /; Table S4: List of homologues to antibiotic resistance genes in the genome sequences of the type strains analyzed in this study; Table S5: Resistance mechanisms of putative resistance gene families identified in type strains of Acetobacter species (A) and Komagataeibacter species (B). Figure S1: Strains of Acetobacter species (A) and Komagataeibacter species (B) resistant to gentamicin (GMN), ampicillin (AMP), chloramphenicol (CHL), ciprofloxacin (CIP), erythromycin (ERY), and trimethoprim (TMP).
Author Contributions
Conceptualization, J.T.; methodology, J.T. and E.C.; analysis, E.C. and J.T.; writing—original draft preparation, J.T.; writing—review and editing, J.T. and E.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Slovenian Research Agency through the programme P2-0006 and the project RI-SI-LifeWatch, co-financed by the Republic of Slovenia, the Ministry of Education, Science and Sport and the European Union from the European Regional Development Fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge Tadeja Vajdič from the University of Maribor for isolation and AMR testing of acetic acid bacteria strains from the vinegar company Simonič in Slovenia, and Jure Škraban and Lora Klanfar from the University of Maribor for the initial experiments of this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Aarts, H.; Margolles, A. Antibiotic resistance genes in food and gut (non pathogenic) bacteria. Bad genes in good bugs. Front. Microbiol. 2015, 5, 754. [Google Scholar] [CrossRef] [PubMed]
- Grevskott, D.H.; Ghavidel, F.Z.; Svanevik, C.S.; Marathe, N.P. Resistance profiles and diversity of β-lactamases in Escherichia coli strains isolated from city-scale sewage surveillance in Bergen, Norway mimic clinical prevalence. Ecotoxicol. Environ. Saf. 2021, 226, 112788. [Google Scholar] [CrossRef] [PubMed]
- Trček, J.; Mahnič, A.; Rupnik, M. Diversity of the microbiota involved in wine and organic apple cider submerged vinegar production as revealed by DHPLC analysis and next-generation sequencing. Int. J. Food Microbiol. 2016, 223, 57–62. [Google Scholar] [CrossRef]
- Škraban, J.; Cleenwerck, I.; Vandamme, P.; Fanedl, L.; Trček, J. Genome sequences and description of novel exopolysaccharides producing species Komagataeibacter pomaceti sp. nov. and reclassification of Komagataeibacter kombuchae (Dutta and Gachhui 2007) Yamada et al., 2013 as a later heterotypic synonym of Komagataeibacter hansenii (Gosselé et al. 1983) Yamada et al., 2013. Syst. Appl. Microbiol. 2018, 41, 581–592. [Google Scholar] [CrossRef]
- Štornik, A.; Skok, B.; Trček, J. Comparison of cultivable acetic acid bacterial microbiota in organic and conventional apple cider vinegar. Food Technol. Biotechnol. 2016, 54, 113–119. [Google Scholar] [CrossRef]
- Trcek, J.; Toyama, H.; Czuba, J.; Misiewicz, A.; Matsushita, K. Correlation between acetic acid resistance and characteristics of PQQ-dependent ADH in acetic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 70, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Gouby, A.; Teyssier, C.; Vecina, F.; Marchandin, H.; Granolleras, C.; Zorgniotti, I.; Jumas-Bilak, E. Acetobacter cibinongensis bacteremia in human. Emerg. Infect. Dis. 2007, 13, 784–785. [Google Scholar] [CrossRef] [PubMed]
- Kohlmann, K.; Barenberg, K.; Anders, A.; Gatermann, S.G. Acetobacter indonesiensis Bacteremia in Child with Metachromatic Leukodystrophy. Emerg. Infect. Dis. 2016, 22, 1681–1683. [Google Scholar] [CrossRef]
- Basu, S.S.; Delaney, M.L.; Li, N.; Onderdonk, A.B.; Bry, L. Acetobacter indonesiensis Pneumonia after Lung Transplantation. Emerg. Infect. Dis. 2018, 24, 598–599. [Google Scholar] [CrossRef]
- Wu, L.; Xie, X.; Li, Y.; Liang, T.; Zhong, H.; Ma, J.; Yang, L.; Yang, J.; Li, L.; Xi, Y.; et al. Metagenomics-based analysis of the age-related cumulative effect of antibiotic resistance genes in gut microbiota. Antibiotics 2021, 10, 1006. [Google Scholar] [CrossRef] [PubMed]
- Trček, J.; Teuber, M. Genetic and restriction analysis of the 16S-23S rDNA internal transcribed spacer regions of the acetic acid bacteria. FEMS Microbiol. Lett. 2002, 208, 69–75. [Google Scholar] [CrossRef]
- Trček, J.; Barja, F. Updates on quick identification of acetic acid bacteria with a focus on the 16S-23S rRNA gene internal transcribed spacer and the analysis of cell proteins by MALDI-TOF mass spectrometry. Int. J. Food Microbiol. 2015, 196, 137–144. [Google Scholar] [CrossRef]
- Sokollek, S.J.; Hammes, W.P. Description of a Starter Culture Preparation for Vinegar Fermentation. Syst. Appl. Microbiol. 1997, 20, 481–491. [Google Scholar] [CrossRef]
- Antimicrobial Susceptibility Testing EUCAST Disk Diffusion Method Version 8.0. 2020, pp. 1–21. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Disk_test_documents/2020_manuals/Manual_v_8.0_EUCAST_Disk_Test_2020.pdf (accessed on 16 June 2020).
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Gorgieva, S.; Trček, J. Bacterial cellulose: Production, modification and perspectives in biomedical applications. Nanomaterials 2019, 9, 1352. [Google Scholar] [CrossRef]
- AlRabiah, H.; Allwood, J.W.; Correa, E.; Xu, Y.; Goodacre, R. pH plays a role in the mode of action of trimethoprim on Escherichia coli. PLoS ONE 2018, 13, e0200272. [Google Scholar] [CrossRef]
- Jiang, M.; Kuang, S.; Lai, S.; Zhang, S.; Yang, J.; Peng, B.; Peng, X.; Chen, Z.G.; Li, H. Na+-NQR Confers Aminoglycoside Resistance via the Regulation of L-Alanine Metabolism. mBIO 2020, 11, e02086-20. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Amyes, S.G.B.; Towner, K.J. Trimethoprim resistance; Epidemiology and molecular aspects. J. Med. Microbiol. 1990, 31, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kahlmeter, G.; Menday, P. Cross-resistance and associated resistance in 2478 Escherichia coli isolates from the Pan-European ECO·SENS project surveying the antimicrobial susceptibility of pathogens from uncomplicated urinary tract infections. J. Antimicrob. Chemother. 2003, 52, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Milanović, V.; Osimani, A.; Cardinali, F.; Litta-Mulondo, A.; Vignaroli, C.; Citterio, B.; Mangiaterra, G.; Aquilanti, L.; Garofalo, C.; Biavasco, F.; et al. Erythromycin-resistant lactic acid bacteria in the healthy gut of vegans, ovo-lacto vegetarians and omnivores. PLoS ONE 2019, 14, e0220549. [Google Scholar] [CrossRef] [PubMed]
- Brisaert, M.; Gabriëls, M.; Plaizier-Vercammen, J. Investigation of the chemical stability of an erythromycin-tretinoin lotion by the use of an optimization system. Int. J. Pharm. 2000, 197, 153–160. [Google Scholar] [CrossRef]
- Torniainen, K.; Tammilehto, S.; Ulvi, V. The effect of pH, buffer type and drug concentration on the photodegradation of ciprofloxacin. Int. J. Pharm. 1996, 131, 53–61. [Google Scholar] [CrossRef]
- Gavrilin, M.V.; Ushakova, L.S.; Gonyan, S.A. Stability of an infusion form of ciprofloxacin hydrochloride. Pharm. Chem. J. 2003, 37, 106–109. [Google Scholar] [CrossRef]
- D’Atanasio, N.; de Joannon, A.C.; Di Sante, L.; Mangano, G.; Ombrato, R.; Vitiello, M.; Bartella, C.; Magarò, G.; Prati, F.; Milanese, C.; et al. Antibacterial activity of novel dual bacterial DNA type II topoisomerase inhibitors. PLoS ONE 2020, 15, e0228509. [Google Scholar] [CrossRef]
- Papkou, A.; Hedge, J.; Kapel, N.; Young, B.; MacLean, R.C. Efflux pump activity potentiates the evolution of antibiotic resistance across S. aureus isolates. Nat. Commun. 2020, 11, 3970. [Google Scholar] [CrossRef]
- Trivedi, M.K.; Patil, S.; Shettigar, H.; Bairwa, K.; Jana, S. Spectroscopic Characterization of Chloramphenicol and Tetracycline: An Impact of Biofield Treatment. Pharm. Anal. Acta 2015, 6, 395. [Google Scholar] [CrossRef]
- Crofts, T.S.; Sontha, P.; King, A.O.; Wang, B.; Biddy, B.A.; Zanolli, N.; Gaumnitz, J.; Dantas, G. Discovery and Characterization of a Nitroreductase Capable of Conferring Bacterial Resistance to Chloramphenicol. Cell Chem. Biol. 2019, 26, 559–570.e6. [Google Scholar] [CrossRef]
- Gilbert, D.N.; Eubanks, N. Effect of pH and human serum on the susceptibility of group D streptococci (Enterococci) to ampicillin in vitro. Antimicrob. Agents Chemother. 1975, 7, 387–395. [Google Scholar] [CrossRef][Green Version]
- Garneau-Tsodikova, S.; Labby, K.J. Mechanisms of Resistance to Aminoglycoside Antibiotics: Overview and Perspectives. Med. Chem. Commun. 2016, 7, 11–27. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 417–433. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).