
Effects of a 6-Month Aerobic Exercise Intervention on Mood and Amygdala Functional Plasticity in Young Untrained Subjects

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
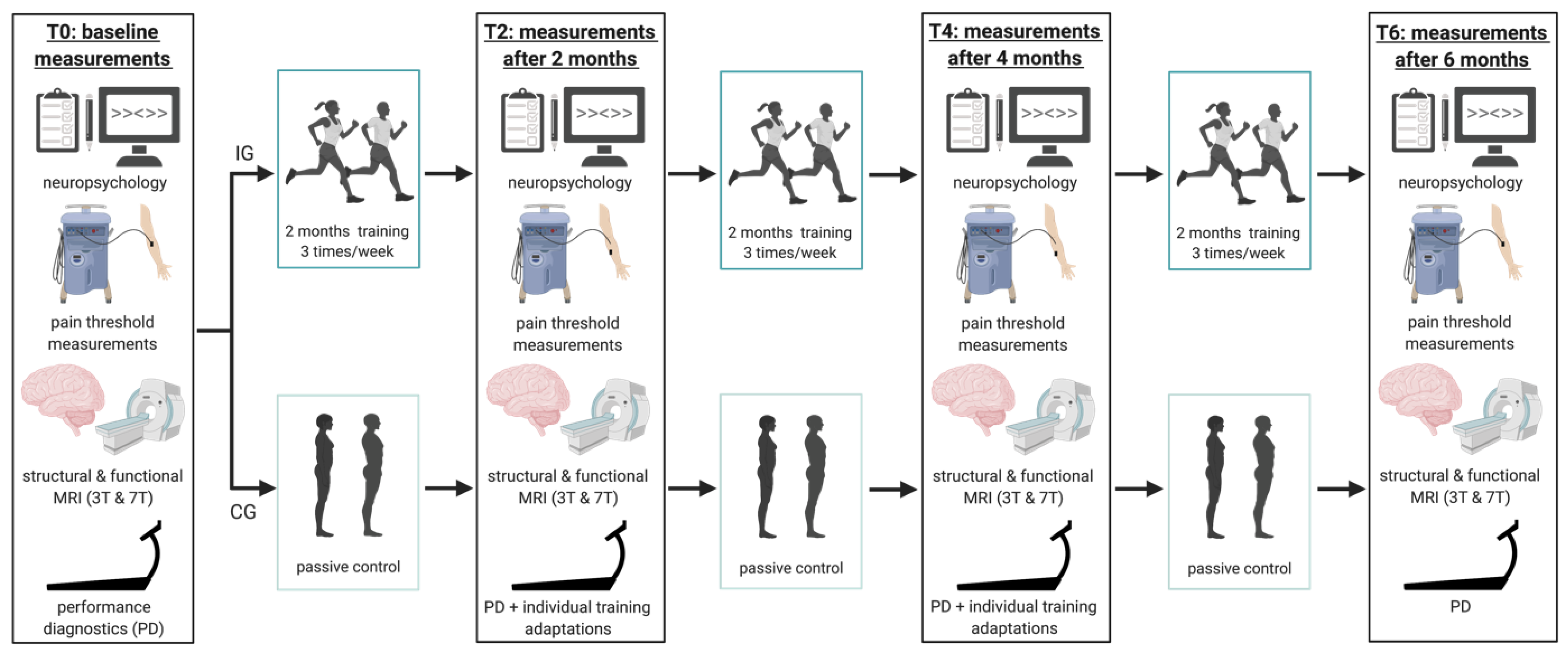
2.2. Experimental Procedure
2.3. Performance Diagnostics
2.4. Intervention
2.5. Questionnaires
2.6. MRI Acquisition
2.7. Amygdala Segmentation
2.8. fMRI Data Preprocessing
2.8.1. Anatomical Data Preprocessing
2.8.2. Functional Data Preprocessing
2.8.3. Seed-to-Whole-Brain Analysis
2.9. Statistical Analysis
2.9.1. Physiological, Behavioral and Structural Data
2.9.2. Resting-State Functional Connectivity
2.9.3. Correlation Analyses
3. Results
3.1. Participants
3.2. Physiological Data—relVO2max
3.3. Questionnaires
3.3.1. STAI State
3.3.2. MoodMeter®
3.4. Structural MRI
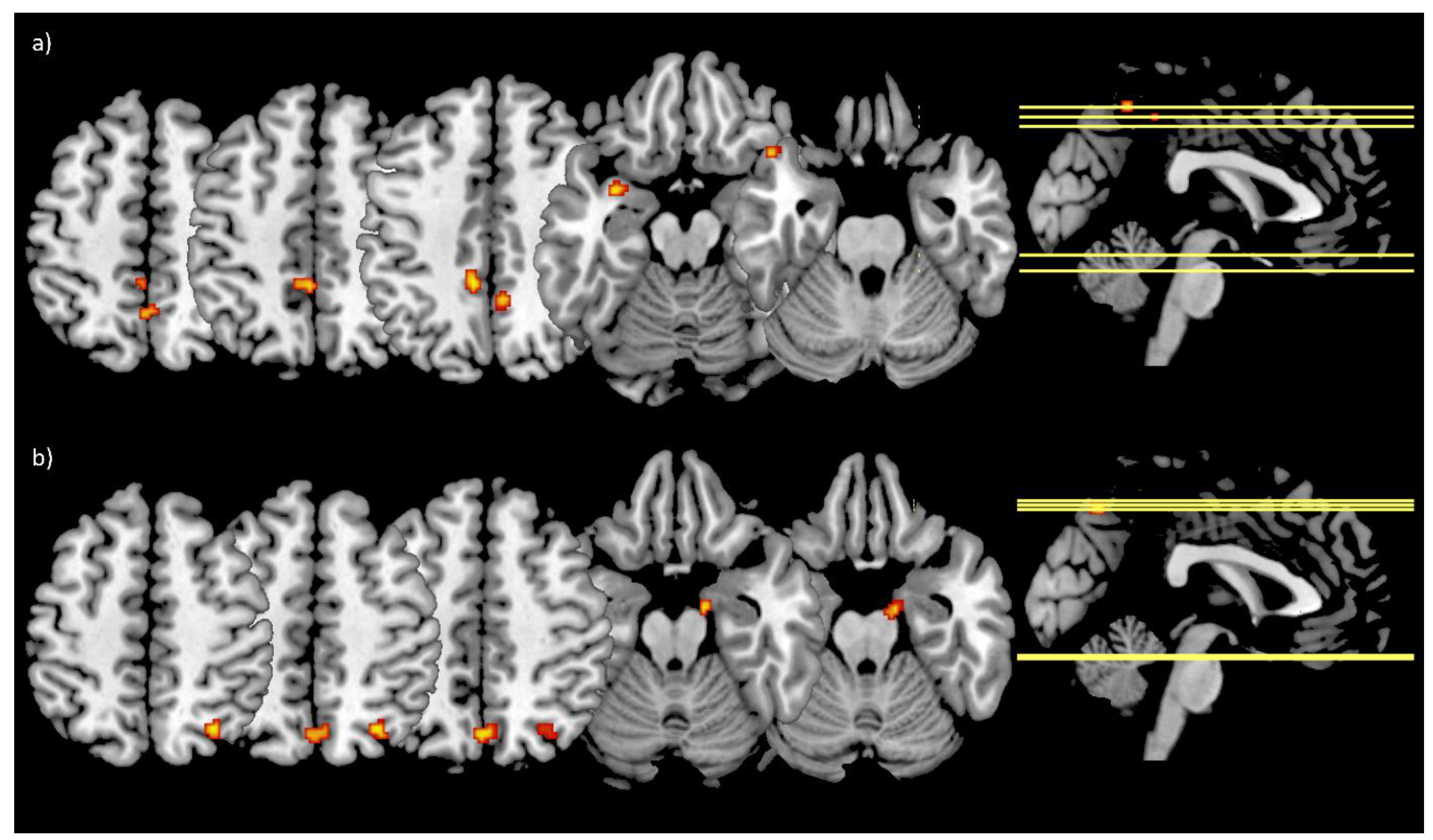
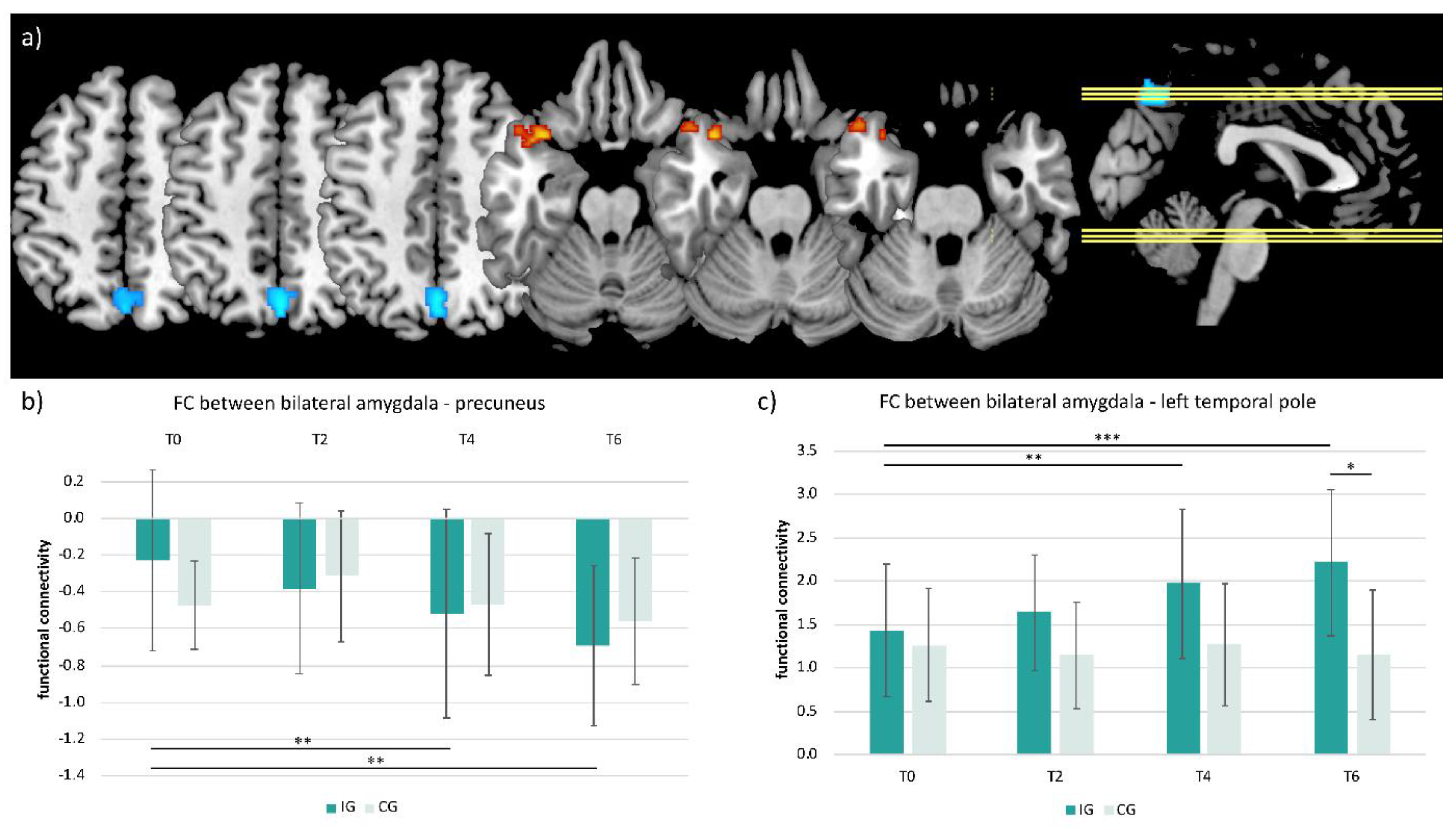
3.5. Functional Connectivity
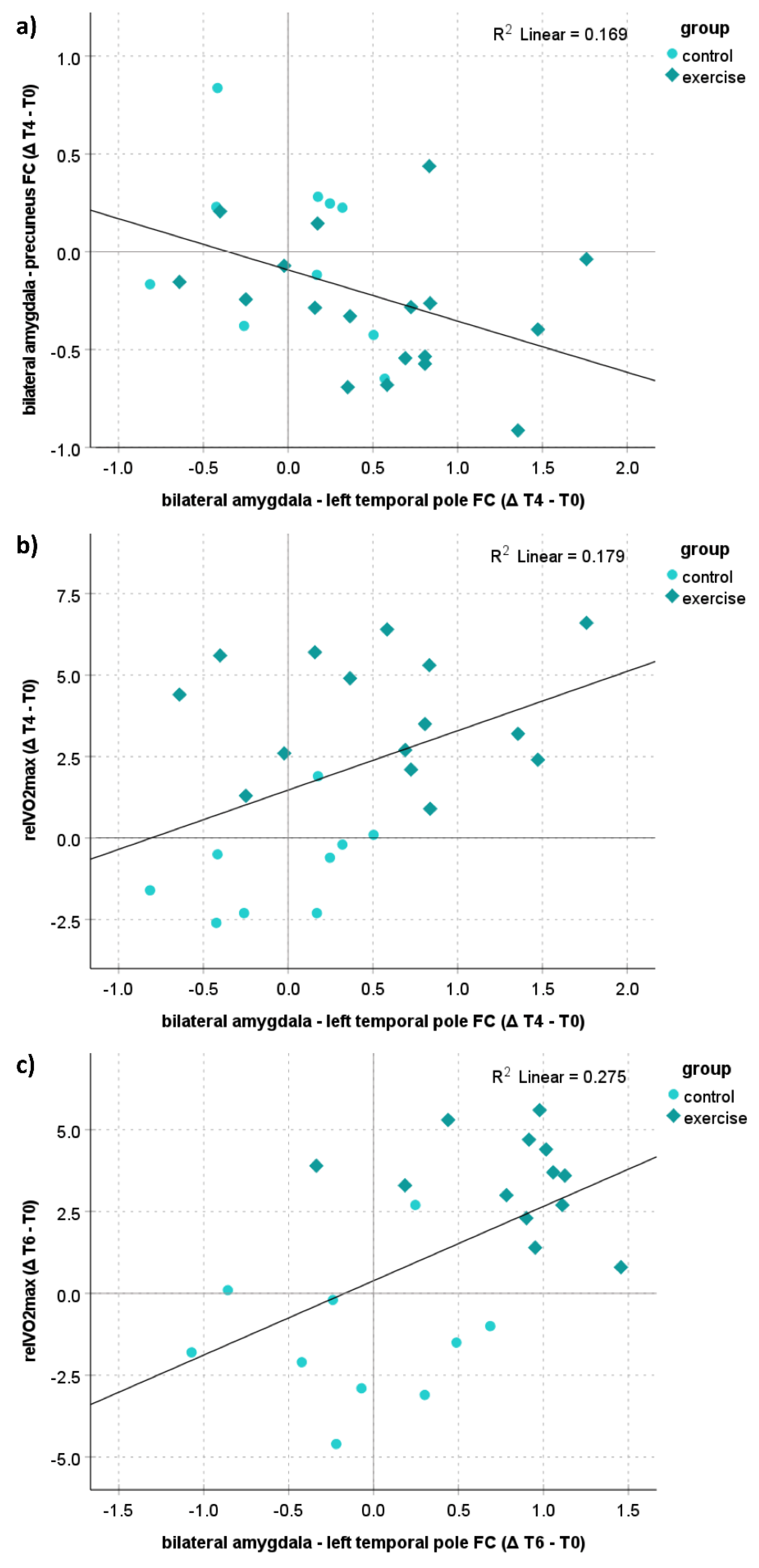
3.6. Correlation Analyses
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirby, L.A.J.; Robinson, J.L. Affective mapping: An activation likelihood estimation (ALE) meta-analysis. Brain Cogn. 2017, 118, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.M. Defining biotypes for depression and anxiety based on large-scale circuit dysfunction: A theoretical review of the evidence and future directions for clinical translation. Depress. Anxiety 2017, 34, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Zhang, Y.; Yang, Z.; Han, S.; Cheng, J. Reduced Brain Gray Matter Volume in Patients with First-Episode Major Depressive Disorder: A Quantitative Meta-Analysis. Front. Psychiatry 2021, 12, 671348. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.P.; Siemer, M.; Gotlib, I.H. Amygdala volume in major depressive disorder: A meta-analysis of magnetic resonance imaging studies. Mol. Psychiatry 2008, 13, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, J. Abnormal neural activities in adults and youths with major depressive disorder during emotional processing: A meta-analysis. Brain Imaging Behav. 2021, 15, 1134–1154. [Google Scholar] [CrossRef]
- Tang, S.; Lu, L.; Zhang, L.; Hu, X.; Bu, X.; Li, H.; Hu, X.; Gao, Y.; Zeng, Z.; Gong, Q.; et al. Abnormal amygdala resting-state functional connectivity in adults and adolescents with major depressive disorder: A comparative meta-analysis. EBioMedicine 2018, 36, 436–445. [Google Scholar] [CrossRef]
- Bourbeau, K.; Moriarty, T.; Ayanniyi, A.; Zuhl, M. The Combined Effect of Exercise and Behavioral Therapy for Depression and Anxiety: Systematic Review and Meta-Analysis. Behav. Sci. 2020, 10, 116. [Google Scholar] [CrossRef]
- Pascoe, M.C.; Parker, A.G. Physical activity and exercise as a universal depression prevention in young people: A narrative review. Early Interv. Psychiatry 2019, 13, 733–739. [Google Scholar] [CrossRef]
- Schuch, F.B.; Stubbs, B. The Role of Exercise in Preventing and Treating Depression. Curr. Sports Med. Rep. 2019, 18, 299–304. [Google Scholar] [CrossRef]
- Schmitt, A.; Upadhyay, N.; Martin, J.A.; Rojas Vega, S.; Struder, H.K.; Boecker, H. Affective Modulation after High-Intensity Exercise Is Associated with Prolonged Amygdalar-Insular Functional Connectivity Increase. Neural Plast. 2020, 2020, 7905387. [Google Scholar] [CrossRef] [Green Version]
- Weng, T.B.; Pierce, G.L.; Darling, W.G.; Falk, D.; Magnotta, V.A.; Voss, M.W. The Acute Effects of Aerobic Exercise on the Functional Connectivity of Human Brain Networks. Brain Plast. 2017, 2, 171–190. [Google Scholar] [CrossRef] [Green Version]
- Ge, L.K.; Hu, Z.; Wang, W.; Siu, P.M.; Wei, G.X. Aerobic Exercise Decreases Negative Affect by Modulating Orbitofrontal-Amygdala Connectivity in Adolescents. Life 2021, 11, 577. [Google Scholar] [CrossRef]
- Tozzi, L.; Carballedo, A.; Lavelle, G.; Doolin, K.; Doyle, M.; Amico, F.; McCarthy, H.; Gormley, J.; Lord, A.; O’Keane, V.; et al. Longitudinal functional connectivity changes correlate with mood improvement after regular exercise in a dose-dependent fashion. Eur. J. Neurosci. 2016, 43, 1089–1096. [Google Scholar] [CrossRef]
- Schmidt, K.; Metzler, P. WST-Wortschatztest; Gött Beltz Test: Göttingen, Germany, 1992. [Google Scholar]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Baker, R.; Dunbar, G.C. The Mini-International Neuropsychiatric Interview (M.I.N.I.): The development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiatry 1998, 59 Suppl. 20, 22–33. [Google Scholar] [PubMed]
- Spielberger, C.; Goruch, R.; Lushene, R.; Vagg, P.; Jacobs, G. Manual for the State-Trait Inventory STAI (form Y); Mind Garden: Palo Alto, CA, USA, 1983. [Google Scholar]
- Hautzinger, M.; Bailer, M.; Worall, H.; Keller, F. Beck-Depressions-Inventar (BDI); Huber: Bern, Switzerland, 1994. [Google Scholar]
- Heatherton, T.F.; Kozlowski, L.T.; Frecker, R.C.; Fagerstrom, K.O. The Fagerstrom Test for Nicotine Dependence: A revision of the Fagerstrom Tolerance Questionnaire. Br. J. Addict. 1991, 86, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Krohne, H.W.; Egloff, B.; Kohlmann, C.W.; Tausch, A. Untersuchungen mit einer deutschen Version der “Positive and Negative Affect Schedule” (PANAS). Diagnostica 1996, 42, 139–156. [Google Scholar]
- Wollseiffen, P.; Ghadiri, A.; Scholz, A.; Struder, H.K.; Herpers, R.; Peters, T.; Schneider, S. Short Bouts of Intensive Exercise During the Workday Have a Positive Effect on Neuro-cognitive Performance. Stress Health 2016, 32, 514–523. [Google Scholar] [CrossRef]
- Bentley, D.J.; Newell, J.; Bishop, D. Incremental exercise test design and analysis: Implications for performance diagnostics in endurance athletes. Sports Med. 2007, 37, 575–586. [Google Scholar] [CrossRef]
- Borg, G. Ratings of Perceived Exertion and Heart-Rates during Short-Term Cycle Exercise and Their Use in a New Cycling Strength Test. Int. J. Sports Med. 1982, 3, 153–158. [Google Scholar] [CrossRef]
- Midgley, A.W.; McNaughton, L.R.; Polman, R.; Marchant, D. Criteria for determination of maximal oxygen uptake: A brief critique and recommendations for future research. Sports Med. 2007, 37, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Knight, R.G.; Waal-Manning, H.J.; Spears, G.F. Some norms and reliability data for the State--Trait Anxiety Inventory and the Zung Self-Rating Depression scale. Br. J. Clin. Psychol. 1983, 22 Pt 4, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Stirnberg, R.; Stöcker, T. Segmented K-space blipped-controlled aliasing in parallel imaging for high spatiotemporal resolution EPI. Magn. Reson. Med. 2020, 85, 1540–1551. [Google Scholar] [CrossRef]
- Stirnberg, R.; Huijbers, W.; Brenner, D.; Poser, B.A.; Breteler, M.; Stöcker, T. Rapid whole-brain resting-state fMRI at 3 T: Efficiency-optimized three-dimensional EPI versus repetition time-matched simultaneous-multi-slice EPI. NeuroImage 2017, 163, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.; Stirnberg, R.; Pracht, E.D.; Stöcker, T. Two-dimensional accelerated MP-RAGE imaging with flexible linear reordering. Magn. Reson. Mater. Phys. Biol. Med. 2014, 27, 455–462. [Google Scholar] [CrossRef]
- Reuter, M.; Schmansky, N.J.; Rosas, H.D.; Fischl, B. Within-subject template estimation for unbiased longitudinal image analysis. NeuroImage 2012, 61, 1402–1418. [Google Scholar] [CrossRef] [Green Version]
- Reuter, M.; Rosas, H.D.; Fischl, B. Highly accurate inverse consistent registration: A robust approach. Neuroimage 2010, 53, 1181–1196. [Google Scholar] [CrossRef] [Green Version]
- Buckner, R.L.; Head, D.; Parker, J.; Fotenos, A.F.; Marcus, D.; Morris, J.C.; Snyder, A.Z. A unified approach for morphometric and functional data analysis in young, old, and demented adults using automated atlas-based head size normalization: Reliability and validation against manual measurement of total intracranial volume. NeuroImage 2004, 23, 724–738. [Google Scholar] [CrossRef]
- Esteban, O.; Birman, D.; Schaer, M.; Koyejo, O.O.; Poldrack, R.A.; Gorgolewski, K.J. MRIQC: Advancing the automatic prediction of image quality in MRI from unseen sites. PLoS ONE 2017, 12, e0184661. [Google Scholar] [CrossRef] [Green Version]
- Esteban, O.; Markiewicz, C.J.; Blair, R.W.; Moodie, C.A.; Isik, A.I.; Erramuzpe, A.; Kent, J.D.; Goncalves, M.; DuPre, E.; Snyder, M.; et al. fMRIPrep: A robust preprocessing pipeline for functional MRI. Nat. Methods 2019, 16, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Gorgolewski, K.; Burns, C.D.; Madison, C.; Clark, D.; Halchenko, Y.O.; Waskom, M.L.; Ghosh, S.S. Nipype: A flexible, lightweight and extensible neuroimaging data processing framework in python. Front. Neuroinform. 2011, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tustison, N.J.; Avants, B.B.; Cook, P.A.; Yuanjie, Z.; Egan, A.; Yushkevich, P.A.; Gee, J.C. N4ITK: Improved N3 Bias Correction. IEEE Trans. Med. Imaging 2010, 29, 1310–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avants, B.B.; Epstein, C.L.; Grossman, M.; Gee, J.C. Symmetric diffeomorphic image registration with cross-correlation: Evaluating automated labeling of elderly and neurodegenerative brain. Med. Image Anal. 2008, 12, 26–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Brady, M.; Smith, S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans. Med. Imaging 2001, 20, 45–57. [Google Scholar] [CrossRef]
- Dale, A.M.; Fischl, B.; Sereno, M.I. Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage 1999, 9, 179–194. [Google Scholar] [CrossRef]
- Klein, A.; Ghosh, S.S.; Bao, F.S.; Giard, J.; Hame, Y.; Stavsky, E.; Lee, N.; Rossa, B.; Reuter, M.; Chaibub Neto, E.; et al. Mindboggling morphometry of human brains. PLoS Comput. Biol. 2017, 13, e1005350. [Google Scholar] [CrossRef]
- Fonov, V.S.; Evans, A.C.; McKinstry, R.C.; Almli, C.R.; Collins, D.L. Unbiased nonlinear average age-appropriate brain templates from birth to adulthood. NeuroImage 2009, 47, S102. [Google Scholar] [CrossRef]
- Glasser, M.F.; Sotiropoulos, S.N.; Wilson, J.A.; Coalson, T.S.; Fischl, B.; Andersson, J.L.; Xu, J.; Jbabdi, S.; Webster, M.; Polimeni, J.R.; et al. The minimal preprocessing pipelines for the Human Connectome Project. Neuroimage 2013, 80, 105–124. [Google Scholar] [CrossRef] [Green Version]
- Greve, D.N.; Fischl, B. Accurate and robust brain image alignment using boundary-based registration. NeuroImage 2009, 48, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, M.; Bannister, P.; Brady, M.; Smith, S. Improved optimization for the robust and accurate linear registration and motion correction of brain images. NeuroImage 2002, 17, 825–841. [Google Scholar] [CrossRef]
- Power, J.D.; Mitra, A.; Laumann, T.O.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Methods to detect, characterize, and remove motion artifact in resting state fMRI. NeuroImage 2014, 84, 320–341. [Google Scholar] [CrossRef]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. NeuroImage 2007, 37, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satterthwaite, T.D.; Elliott, M.A.; Gerraty, R.T.; Ruparel, K.; Loughead, J.; Calkins, M.E.; Eickhoff, S.B.; Hakonarson, H.; Gur, R.C.; Gur, R.E.; et al. An improved framework for confound regression and filtering for control of motion artifact in the preprocessing of resting-state functional connectivity data. NeuroImage 2013, 64, 240–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero-Gaudes, C.; Reynolds, R.C. Methods for cleaning the BOLD fMRI signal. NeuroImage 2017, 154, 128–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciric, R.; Wolf, D.H.; Power, J.D.; Roalf, D.R.; Baum, G.L.; Ruparel, K.; Shinohara, R.T.; Elliott, M.A.; Eickhoff, S.B.; Davatzikos, C.; et al. Benchmarking of participant-level confound regression strategies for the control of motion artifact in studies of functional connectivity. NeuroImage 2017, 154, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.W. AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Comput. Biomed. Res. 1996, 29, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.W.; Hyde, J.S. Software tools for analysis and visualization of fMRI data. NMR Biomed. 1997, 10, 171–178. [Google Scholar] [CrossRef]
- Desikan, R.S.; Segonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- Nickerson, L.D.; Smith, S.M.; Ongur, D.; Beckmann, C.F. Using Dual Regression to Investigate Network Shape and Amplitude in Functional Connectivity Analyses. Front. Neurosci. 2017, 11, 115. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.6.0. 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 20 May 2021).
- Kenward, M.G.; Roger, J.H. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J. A power primer. Psychol. Bull. 1992, 112, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, T.J.; Bhat, J.; Horwege, A.M.; Mathalon, D.H.; Glover, G.H.; Roach, B.J.; Badran, B.W.; Forman, S.D.; George, M.S.; Scott, J.C.; et al. Ruminative reflection is associated with anticorrelations between the orbitofrontal cortex and the default mode network in depression: Implications for repetitive transcranial magnetic stimulation. Brain Imaging Behav. 2021, 16, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Feurer, C.; Jimmy, J.; Chang, F.; Langenecker, S.A.; Phan, K.L.; Ajilore, O.; Klumpp, H. Resting state functional connectivity correlates of rumination and worry in internalizing psychopathologies. Depress. Anxiety 2021, 38, 488–497. [Google Scholar] [CrossRef]
- Herlin, B.; Navarro, V.; Dupont, S. The temporal pole: From anatomy to function-A literature appraisal. J. Chem. Neuroanat. 2021, 113, 101925. [Google Scholar] [CrossRef]
- Olson, I.R.; Plotzker, A.; Ezzyat, Y. The Enigmatic temporal pole: A review of findings on social and emotional processing. Brain 2007, 130, 1718–1731. [Google Scholar] [CrossRef] [Green Version]
- Bistricky, S.L.; Ingram, R.E.; Atchley, R.A. Facial affect processing and depression susceptibility: Cognitive biases and cognitive neuroscience. Psychol. Bull. 2011, 137, 998–1028. [Google Scholar] [CrossRef]
- Zobel, I.; Werden, D.; Linster, H.; Dykierek, P.; Drieling, T.; Berger, M.; Schramm, E. Theory of mind deficits in chronically depressed patients. Depress. Anxiety 2010, 27, 821–828. [Google Scholar] [CrossRef]
- Fan, L.; Wang, J.; Zhang, Y.; Han, W.; Yu, C.; Jiang, T. Connectivity-Based Parcellation of the Human Temporal Pole Using Diffusion Tensor Imaging. Cereb. Cortex 2014, 24, 3365–3378. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Liu, J.; Nie, B.; Li, Y.; Shan, B.; Wang, G.; Li, K. Cerebral and cerebellar gray matter reduction in first-episode patients with major depressive disorder: A voxel-based morphometry study. Eur. J. Radiol. 2011, 80, 395–399. [Google Scholar] [CrossRef]
- Colcombe, S.; Kramer, A.F. Fitness effects on the cognitive function of older adults: A meta-analytic study. Psychol. Sci. 2003, 14, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Saraulli, D.; Costanzi, M.; Mastrorilli, V.; Farioli-Vecchioli, S. The Long Run: Neuroprotective Effects of Physical Exercise on Adult Neurogenesis from Youth to Old Age. Curr. Neuropharmacol. 2017, 15, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.; Steffens, D.C.; Wang, L. Effects of physical exercise on the aging brain across imaging modalities: A meta-analysis of neuroimaging studies in randomized controlled trials. Int. J. Geriatr. Psychiatry 2021, 36, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Salvan, P.; Wassenaar, T.; Wheatley, C.; Beale, N.; Cottaar, M.; Papp, D.; Bastiani, M.; Fitzgibbon, S.; Duff, E.; Andersson, J.; et al. Multimodal Imaging Brain Markers in Early Adolescence Are Linked with a Physically Active Lifestyle. J. Neurosci. 2021, 41, 1092–1104. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Intervention N = 18 (m/f: 7/11) | Control N = 10 (m/f: 6/4) | p-Values Independent t-Test * |
|---|---|---|---|
| Age [years] | 23.9 ± 3.9 | 23.7 ± 4.2 | 0.879 |
| Height [cm] | 173.6 ± 12.1 | 176.9 ± 7.9 | 0.447 |
| Weight [kg] | 69.9 ± 15.1 | 71.2 ± 14.1 | 0.816 |
| BMI [kg/m2] | 23.1 ± 3.7 | 22.7 ± 3.6 | 0.771 |
| HRmax [bpm] | 198.5 ± 7.6 | 200.8 ± 8.5 | 0.467 |
| relVO2max [mL/min/kg] | 38.5 ± 3.4 | 41.7 ± 7.5 | 0.232 |
| Education [years] | 16.3 ± 3.1 | 15.8 ± 3.1 | 0.730 |
| EHI [L.Q.] | 74.2 ± 16.2 | 79.5 ± 13.3 | 0.390 |
| BDI | 2.6 ± 3.4 | 1.4 ± 1.5 | 0.224 |
| STAI trait | 33.9 ± 9.3 | 31.1 ± 5.8 | 0.390 |
| WST IQ | 107.0 ± 9.9 | 107.3 ± 8.8 | 0.937 |
| Questionnaire | Dimension | Group | T0 | T2 | T4 | T6 |
|---|---|---|---|---|---|---|
| STAI | State anxiety | Intervention | 33.3 ± 6.7 | 31.3 ± 5.9 | 32.1 ± 8.5 | 29.5 ± 5.4 |
| Control | 35.9 ± 7.3 | 31.6 ± 3.3 | 34.7 ± 5.0 | 32.2 ± 5.2 | ||
| PANAS | Positive affect scale | Intervention | 28.4 ± 6.4 | 27.1 ± 7.1 | 26.5 ± 7.4 | 30.1 ± 8.5 |
| Control | 25.4 ± 6.9 | 27.6 ± 7.4 | 25.9 ± 5.7 | 26.9 ± 7.0 | ||
| Negative affect scale | Intervention | 11.6 ± 1.6 | 10.9 ± 1.1 | 11.7 ± 2.5 | 10.8 ± 1.3 | |
| Control | 12.7 ± 3.3 | 11.3 ± 1.5 | 11.2 ± 1.7 | 11.2 ± 2.1 | ||
| MoodMeter® | PEPS | Intervention | 3.4 ± 0.8 | 3.6 ± 0.5 | 3.6 ± 0.5 | 3.7 ± 0.6 |
| Control | 3.1 ± 0.6 | 2.9 ± 0.4 | 3.1 ± 0.7 | 3.3 ± 0.7 | ||
| Physical energy | Intervention | 3.9 ± 1.1 | 4.0 ± 0.9 | 3.9 ± 1.1 | 3.8 ± 1.1 | |
| Control | 4.0 ± 0.6 | 4.3 ± 0.7 | 4.0 ± 0.9 | 4.2 ± 0.7 | ||
| Physical fitness | Intervention | 2.5 ± 1.1 | 2.8 ± 0.7 | 2.9 ± 0.9 | 3.0 ± 0.9 | |
| Control | 2.0 ± 1.0 | 1.8 ± 0.9 | 2.3 ± 0.8 | 2.3 ± 1.2 | ||
| Physical health | Intervention | 4.4 ± 0.6 | 4.4 ± 0.7 | 4.3 ± 0.8 | 4.5 ± 0.6 | |
| Control | 4.0 ± 0.9 | 3.6 ± 1.0 | 3.8 ± 1.0 | 4.2 ± 1.0 | ||
| Physical flexibility | Intervention | 2.9 ± 1.1 | 3.2 ± 0.7 | 3.2 ± 0.9 | 3.5 ± 0.7 | |
| Control | 2.3 ± 0.8 | 2.1 ± 1.0 | 2.4 ± 0.9 | 2.7 ± 0.9 | ||
| PSYCHO | Intervention | 3.6 ± 1.1 | 3.8 ± 0.7 | 3.6 ± 1.0 | 3.7 ± 0.9 | |
| Control | 3.6 ± 0.5 | 3.7 ± 1.0 | 3.5 ± 0.7 | 3.6 ± 0.8 | ||
| Positive mood | Intervention | 3.6 ± 1.2 | 3.8 ± 1.1 | 3.6 ± 1.3 | 3.8 ± 1.1 | |
| Control | 3.2 ± 1.2 | 3.5 ± 0.9 | 3.2 ± 1.1 | 3.4 ± 0.9 | ||
| Calmness | Intervention | 4.0 ± 0.9 | 4.3 ± 0.6 | 4.2 ± 0.8 | 4.2 ± 0.9 | |
| Control | 3.7 ± 1.3 | 3.9 ± 0.9 | 3.6 ± 1.3 | 4.0 ± 1.0 | ||
| Recovery | Intervention | 3.2 ± 1.4 | 3.3 ± 1.0 | 3.4 ± 1.2 | 3.4 ± 1.1 | |
| Control | 3.4 ± 0.8 | 3.3 ± 1.3 | 3.2 ± 0.9 | 3.1 ± 1.1 | ||
| Relaxation | Intervention | 3.6 ± 1.2 | 3.6 ± 1.1 | 3.4 ± 1.3 | 3.4 ± 1.4 | |
| Control | 4.1 ± 0.8 | 4.1 ± 1.2 | 3.8 ± 1.0 | 4.1 ± 0.9 | ||
| MOT | Intervention | 3.3 ± 0.9 | 3.4 ± 0.7 | 3.3 ± 0.8 | 3.5 ± 0.7 | |
| Control | 2.7 ± 0.9 | 2.7 ± 0.9 | 2.8 ± 0.6 | 2.9 ± 1.1 | ||
| Willingness to seek contact | Intervention | 3.6 ± 0.8 | 3.4 ± 1.2 | 3.2 ± 0.9 | 3.4 ± 1.0 | |
| Control | 2.6 ± 1.3 | 2.7 ± 0.6 | 2.6 ± 0.6 | 2.7 ± 1.4 | ||
| Social acceptance | Intervention | 3.5 ± 1.1 | 3.7 ± 0.8 | 3.5 ± 0.9 | 3.8 ± 0.9 | |
| Control | 3.1 ± 1.1 | 3.3 ± 1.3 | 3.1 ± 0.9 | 3.2 ± 1.5 | ||
| Readiness to strain | Intervention | 2.9 ± 1.3 | 3.1 ± 0.9 | 3.0 ± 1.0 | 2.8 ± 1.1 | |
| Control | 2.4 ± 0.9 | 2.1 ± 1.2 | 2.6 ± 0.6 | 2.3 ± 1.0 | ||
| Self-confidence | Intervention | 3.1 ± 0.9 | 3.4 ± 0.8 | 3.5 ± 0.7 | 3.9 ± 0.7 | |
| Control | 2.8 ± 0.9 | 2.6 ± 1.1 | 3.1 ± 1.1 | 3.6 ± 1.1 |
| Dimension | Effect of Time | Effect of Group | Time × Group Interaction | Effect of Sex | Effect of Age | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p-Value | df | F | p-Value | df | F | p-Value | df | F | p-Value | df | F | p-Value | |
| Perceived physical state (PEPS) | 1, 73.18 | 3.66 | 0.060 | 1, 40.87 | 1.81 | 0.187 | 1, 73.17 | 0.03 | 0.868 | 1, 23.87 | 0.37 | 0.551 | 1, 23.53 | 1.57 | 0.223 |
| Physical energy | 1, 72.84 | 0.00 | 0.950 | 1, 46.15 | 0.36 | 0.550 | 1, 72.83 | 0.58 | 0.447 | 1, 23.43 | 1.34 | 0.260 | 1, 22.98 | 0.18 | 0.673 |
| Physical fitness | 1, 73.54 | 7.58 | 0.007 ** | 1, 37.81 | 2.20 | 0.146 | 1, 73.53 | 0.35 | 0.558 | 1, 24.30 | 0.06 | 0.808 | 1, 24.02 | 4.46 | 0.045 * |
| Physical health | 1, 73.34 | 0.77 | 0.384 | 1, 36.13 | 2.59 | 0.116 | 1, 73.34 | 0.49 | 0.488 | 1, 24.13 | 0.02 | 0.896 | 1, 23.88 | 0.08 | 0.780 |
| Physical flexibility | 1, 73.56 | 6.40 | 0.014 * | 1, 48.43 | 3.43 | 0.070 | 1, 73.55 | 0.17 | 0.685 | 1, 24.12 | 0.39 | 0.540 | 1, 23.64 | 2.73 | 0.112 |
| Psychological strain (PSYCHO) | 1, 73.30 | 0.00 | 0.951 | 1, 44.48 | 0.01 | 0.932 | 1, 73.29 | 0.00 | 0.953 | 1, 23.29 | 1.72 | 0.202 | 1, 23.52 | 1.74 | 0.200 |
| Positive mood | 1, 73.42 | 0.27 | 0.605 | 1, 38.31 | 0.26 | 0.610 | 1, 73.41 | 0.03 | 0.862 | 1, 24.17 | 4.05 | 0.056 | 1, 23.88 | 3.37 | 0.079 |
| Calmness | 1, 73.78 | 0.60 | 0.442 | 1, 47.31 | 1.22 | 0.276 | 1, 73.77 | 0.08 | 0.781 | 1, 24.38 | 0.69 | 0.416 | 1, 23.92 | 0.37 | 0.550 |
| Recovery | 1, 73.34 | 0.06 | 0.808 | 1, 52.48 | 0.35 | 0.558 | 1, 73.33 | 0.84 | 0.363 | 1, 23.82 | 0.99 | 0.330 | 1, 23.26 | 0.76 | 0.391 |
| Relaxation | 1, 73.15 | 0.50 | 0.484 | 1, 70.58 | 1.20 | 0.277 | 1, 73.13 | 0.31 | 0.581 | 1, 23.22 | 0.21 | 0.651 | 1, 22.30 | 1.11 | 0.304 |
| Motivational state (MOT) | 1, 73.29 | 3.10 | 0.083 | 1, 32.77 | 2.75 | 0.107 | 1, 73.28 | 0.19 | 0.668 | 1, 24.13 | 1.42 | 0.246 | 1, 23.95 | 1.35 | 0.257 |
| Willingness to seek contact | 1, 73.43 | 0.00 | 0.994 | 1, 43.31 | 4.84 | 0.033 * | 1, 73.42 | 0.59 | 0.444 | 1, 24.09 | 3.89 | 0.060 | 1, 23.70 | 0.22 | 0.646 |
| Social acceptance | 1, 73.24 | 0.81 | 0.371 | 1, 31.77 | 0.52 | 0.476 | 1, 73.23 | 0.35 | 0.556 | 1, 24.10 | 1.26 | 0.273 | 1, 23.93 | 0.28 | 0.604 |
| Readiness to strain | 1, 73.42 | 0.11 | 0.744 | 1, 40.17 | 1.45 | 0.236 | 1, 73.42 | 0.12 | 0.732 | 1, 24.14 | 1.47 | 0.237 | 1, 23.81 | 4.88 | 0.037 * |
| Self-confidence | 1, 73.34 | 31.19 | <0.001 *** | 1, 33.99 | 2.21 | 0.147 | 1, 73.33 | 0,33 | 0.570 | 1, 24.16 | 0.06 | 0.803 | 1, 23.95 | 0.78 | 0.385 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurer, A.; Klein, J.; Claus, J.; Upadhyay, N.; Henschel, L.; Martin, J.A.; Scheef, L.; Daamen, M.; Schörkmaier, T.; Stirnberg, R.; et al. Effects of a 6-Month Aerobic Exercise Intervention on Mood and Amygdala Functional Plasticity in Young Untrained Subjects. Int. J. Environ. Res. Public Health 2022, 19, 6078. https://doi.org/10.3390/ijerph19106078
Maurer A, Klein J, Claus J, Upadhyay N, Henschel L, Martin JA, Scheef L, Daamen M, Schörkmaier T, Stirnberg R, et al. Effects of a 6-Month Aerobic Exercise Intervention on Mood and Amygdala Functional Plasticity in Young Untrained Subjects. International Journal of Environmental Research and Public Health. 2022; 19(10):6078. https://doi.org/10.3390/ijerph19106078
Chicago/Turabian StyleMaurer, Angelika, Julian Klein, Jannik Claus, Neeraj Upadhyay, Leonie Henschel, Jason Anthony Martin, Lukas Scheef, Marcel Daamen, Theresa Schörkmaier, Rüdiger Stirnberg, and et al. 2022. "Effects of a 6-Month Aerobic Exercise Intervention on Mood and Amygdala Functional Plasticity in Young Untrained Subjects" International Journal of Environmental Research and Public Health 19, no. 10: 6078. https://doi.org/10.3390/ijerph19106078
APA StyleMaurer, A., Klein, J., Claus, J., Upadhyay, N., Henschel, L., Martin, J. A., Scheef, L., Daamen, M., Schörkmaier, T., Stirnberg, R., Stöcker, T., Radbruch, A., Attenberger, U. I., Reuter, M., & Boecker, H. (2022). Effects of a 6-Month Aerobic Exercise Intervention on Mood and Amygdala Functional Plasticity in Young Untrained Subjects. International Journal of Environmental Research and Public Health, 19(10), 6078. https://doi.org/10.3390/ijerph19106078