
Comparative Detection of Immunoglobulin Isotypes and Subclasses against Toxoplasma gondii Soluble Antigen in Serum and Colostrum Samples from Puerperal Women

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Patients
2.2. Human Colostrum Samples
2.3. Blood Samples
2.4. Maintenance of T. gondii
2.5. Indirect ELISA for Detection of Anti-T. gondii IgG Total and Subclasses
2.6. Capture ELISA for Detection of Anti-T. gondii IgM and IgA
2.7. Statistical Analysis
3. Results
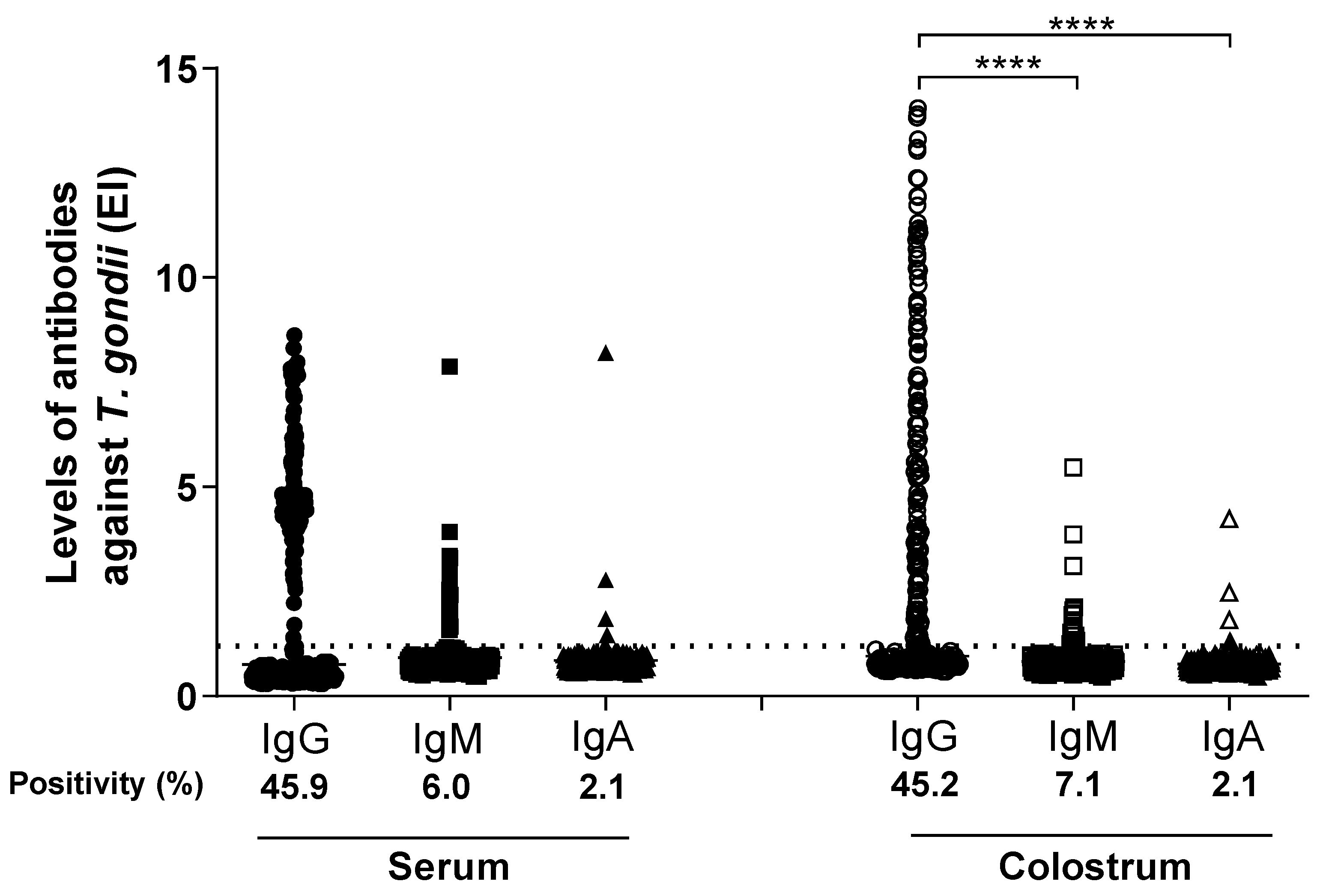
3.1. Detection of T. gondii-Specific IgG, IgM, and IgA Antibodies in Paired Serum and Colostrum Samples
3.2. Detection of T. gondii-Specific IgG1, IgG3, and IgG4 Subclasses in Paired Serum and Colostrum Samples
3.3. Correlation between Anti-T. gondii IgG1, IgG3, and IgG4 Subclass Levels in Serum and Colostrum Samples
3.4. Associations between IgG1, IgG3, and IgG4 Subclasses and T. gondii-Specific IgG, IgM, and IgA Isotypes in Serum and Colostrum Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Dubey, J.P.; Lago, E.G.; Gennari, S.M.; Su, C.; Jones, J.L. Toxoplasmosis in humans and animals in Brazil: High prevalence, high burden of disease, and epidemiology. Parasitology 2012, 139, 1375–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francisco, F.M.; Souza, S.L.P.; Gennari, S.M.; Pinheiro, S.R.; Muradian, V.; Soares, R.M. Seroprevalence of toxoplasmosis in a low-income community in the São Paulo municipality, SP, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2006, 48, 167–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, F.L.; Amendoeira, M.R.R.; Bastos, O.M.P.; Mattos, D.P.B.G.; Fonseca, A.B.M.; Nicolau, J.L.; Neves, L.B.; Millar, P.R. Prevalence and risk factors for Toxoplasma gondii infection among pregnant and postpartum women attended at public healthcare facilities in the City of Niterói, State of Rio de Janeiro, Brazil. Rev. Soc. Bras. Med. Trop. 2013, 46, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spalding, S.M.; Amendoeira, M.R.R.; Klein, C.H.; Ribeiro, L.C. Serological screening and toxoplasmosis exposure factors among pregnant women in South of Brazil. Rev. Soc. Bras. Med. Trop. 2005, 38, 173–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, A.D.M.; Oliveira, L.A.R.; Oliveira, M.F.B.; Santos, R.C.S.; Araújo, R.M.; Alves, J.A.B.; Pinheiro, K.S.; Gurgelli, R.Q.; Mussi-Pinhata, M.M. Soroprevalência de anticorpos para toxoplasmose, rubéola, citomegalovírus, sífilis e HIV em gestantes sergipanas. Rev. Soc. Bras. Med. Trop. 2009, 42, 532–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, M.A.S.; Matos, C.C.B.; Spegiorin, L.C.J.F.; Oliani, D.C.M.V.; Oliani, A.H.; Mattos, L.C. Seropositivity rates for toxoplasmosis, rubella, syphilis, cytomegalovirus, hepatitis and HIV among pregnant women receiving care at a public health service, Sao Paulo state, Brazil. Braz. J. Infect. Dis. 2010, 14, 601–605. [Google Scholar] [CrossRef] [Green Version]
- Segundo, G.R.S.; Silva, D.A.O.; Mineo, J.R.; Ferreira, M.S. Congenital toxoplasmosis in Uberlandia, MG, Brazil. J. Trop. Pediatr. 2004, 50, 50–53. [Google Scholar] [CrossRef] [Green Version]
- Omonijo, A.O.; Kalinda, C.; Mukaratirwa, S. Toxoplasma gondii infections in animals and humans in Southern Africa: A systematic review and meta-analysis. Pathogens 2022, 11, 183. [Google Scholar] [CrossRef]
- López Ureña, N.M.; Chaudhry, U.; Calero Bernal, R.; Cano Alsua, S.; Messina, D.; Evangelista, F.; Betson, M.; Lalle, M.; Jokelainen, P.; Ortega Mora, L.M.; et al. Contamination of soil, water, fresh produce, and bivalve mollusks with Toxoplasma gondii oocysts: A systematic review. Microorganisms 2022, 10, 517. [Google Scholar] [CrossRef]
- Ahmadpour, E.; Rahimi, M.T.; Ghojoghi, A.; Rezaei, F.; Hatam-Nahavandi, K.; Oliveira, S.M.R.; de Lourdes Pereira, M.; Majidiani, H.; Siyadatpanah, A.; Elhamirad, S.; et al. Toxoplasma gondii infection in marine animal species, as a potential source of food contamination: A systematic review and meta-analysis. Acta Parasitol. 2022, 67, 592–605. [Google Scholar] [CrossRef]
- Moreno-Mesonero, L.; Amorós, I.; Moreno, Y.; Alonso, J.L. Simultaneous detection of less frequent waterborne parasitic protozoa in reused wastewater using amplicon sequencing and qPCR techniques. J. Environ. Manag. 2022, 314, 115029. [Google Scholar] [CrossRef]
- Liu, L.; Liu, T.; Yu, L.; Cai, Y.; Zhang, A.; Xu, X.; Luo, Q.; Hu, Y.; Song, W.; Lun, Z.; et al. rROP2 (186–533): A novel peptide antigen for detection of IgM antibodies against Toxoplasma gondii. Foodborne Pathog. Dis. 2012, 9, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, W.Y.; Nam, H.W.; Youn, J.H.; Kim, D.J.; Kong, Y.; Kang, S.Y.; Cho, S.Y. Detection of antibodies in serum and cerebrospinal fluid to Toxoplasma gondii by indirect latex agglutination test and enzyme-linked immunosorbent assay. Kisaengchunghak Chapchi 1992, 30, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Araujo-Andrade, C.; Pichardo-Molina, J.L.; Barbosa-Sabanero, G.; Frausto-Reyes, C.; Torres-Lopez, A. Detection of the presence of antibodies against Toxoplasma gondii in human colostrum by Raman spectroscopy and principal component analysis. J. Biomed. Opt. 2007, 12, 034006. [Google Scholar] [CrossRef] [PubMed]
- Lamounier, J.A.; Vieira, G.O.; Gouvêa, L.C. Composição do leite humano—Fatores nutricionais. In Aleitamento Materno; Rego, J.D., Ed.; São Paulo: Atheneu, Brazil, 2002; pp. 47–58. [Google Scholar]
- Lawrence, R.A.; Lawrence, R.M. Breastfeeding: A Guide for the Medical Profession; Elsevier Mosby: Amsterdam, The Netherlands, 2005; p. 1152. [Google Scholar]
- Euclydes, M.P. Nutrição do Lactente, Base Científica Para uma Alimentação Saudável, 3rd ed.; Suprema: Viçosa, Brazil, 2005; p. 548. [Google Scholar]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 4974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellote, C.; Casillas, R.; Ramirez-Santana, C.; Perez-Cano, F.J.; Castell, M.; Moretones, M.G.; López-Sabater, M.C.; Franch, A. Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. J. Nutr. 2011, 141, 1181–1187. [Google Scholar] [CrossRef]
- Neto, M.T. Aleitamento materno e infecção ou da importância do mesmo na sua prevenção. Acta Pediátrica Port. 2006, 37, 23–27. [Google Scholar]
- Chirico, G.; Marzollo, R.; Cortinovis, S.; Fonte, C.; Gasparoni, A. Antiinfective Properties of Human Milk. J. Nutr. 2008, 138, 1801S–1806S. [Google Scholar] [CrossRef] [Green Version]
- Ladomenou, F.; Moschandreas, J.; Kafatos, A.; Tselentis, Y.; Galanakis, E. Protective effect of exclusive breastfeeding against infections during infancy: A prospective study. Arch. Dis. Child. 2010, 95, 1004–1008. [Google Scholar] [CrossRef]
- Iyengar, S.R.; Walker, W.A. Immune factors in breast milk and the development of atopic disease. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 641–647. [Google Scholar] [CrossRef]
- Chong-Neto, H.; Solé, D.; Camargos, P.; Rosário, N.; Sarinho, S.; Chong-Silva, D.C.; Kiertsman, B.; Pastorino, A.C.; Sano, F.; Urrutia-Pereira, M.; et al. Diretrizes da Associação Brasileira de Alergia e Imunologia e Sociedade Brasileira de Pediatria para sibilância e asma no pré-escolar. Arq. Asma Alerg. Imunol. 2018, 2, 163–208. [Google Scholar] [CrossRef]
- Silva, N.A.; Honorio-França, A.C.; Giachini, F.R.; Mores, L.; Souza, E.G.; França, E.L. Bioactive factors of colostrum and human milk exhibit a day-night variation. Am. J. Immunol. 2013, 9, 68–74. [Google Scholar] [CrossRef]
- Oliveira, A.C.M.; Borges, H.D.S.; Carvalho, F.R.; Macedo, A.G.J.; Mota, C.M.; Oliveira, A.M.M.; Santiago, F.M.; Araújo, C.G.A.; Silva, D.A.O.; Mineo, T.W.P.; et al. Evaluation of colostrum as an alternative biological sample for the diagnosis of human congenital toxoplasmosis. BMC Infect. Dis. 2015, 15, 519. [Google Scholar] [CrossRef]
- Brasil Agência Nacional de Vigilância Sanitária. Banco de Leite Humano: Funcionamento, Prevenção e Controle de Riscos; Anvisa: Brasília, Brazil, 2008; p. 160. [Google Scholar]
- Ribeiro, D.P.; Freitas, M.M.P.; Cardoso, M.R.D.; Pajuaba, A.C.A.M.; Silva, N.M.; Mineo, T.W.P.; Silva, J.S.; Mineo, J.R.; Deise, A.O.; Silva, D.A.O. CpG-ODN combined with Neospora caninum lysate, but not with excreted-secreted antigen, enhances protection against infection in mice. Vaccine 2009, 27, 2570–2579. [Google Scholar] [CrossRef]
- Mineo, J.R.; Camargo, M.E.; Ferreira, A.W.; Almeida, G. Research on IgM anti-Toxoplasma gondii antibodies by using a reverse immunoenzymatic techique. Rev. Inst. Med. Trop. Sao Paulo 1986, 28, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.B.; Nakane, P.K. Recent developments in the periodate method of conjugating horseradish peroxidase (HRPO) to antibodies. In Immunofluorescence and Related Staining Techniques; Knapp, W., Holubar, K., Wick, G., Eds.; Elsevier North-Holland Biomedical Press: Amsterdam, The Netherlands, 1978; pp. 215–224. [Google Scholar]
- Grey, H.M.; Kunkel, H.G. H chain subgroups of myeloma proteins and normal 7S gamma-globulin. J. Exp. Med. 1964, 120, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Technical Series; World Health Organization: Geneva, Switzerland, 1966; p. WHA35. [Google Scholar]
- Lambin, P. Les sous classes d’immunoglobulines IgG Aspects biochimiques et cliniques. Rev. Française Transfus D’hémobiologie 1989, 32, 357–376. [Google Scholar] [CrossRef]
- Smith, T.F. IgG subclasses. Adv. Pediatr. 1992, 39, 101–126. [Google Scholar]
- van de Winkel, J.G.; Anderson, C.L. Biology of human immunoglobulin G Fc receptors. J. Leukoc. Biol. 1991, 49, 511–524. [Google Scholar] [CrossRef]
- Xu, Y.; Oomen, R.; Klein, M.H. Residue at position 331 in the IgG1 and IgG4 CH2 domains contributes to their differential ability to bind and activate complement. J. Biol. Chem. 1994, 269, 3469–3474. [Google Scholar] [CrossRef]
- Brekke, O.H.; Michaelsen, T.E.; Sandlie, I. The structural requirements for complement activation by IgG: Does it hinge on the hinge? Immunol. Today 1995, 16, 85–90. [Google Scholar] [CrossRef]
- Huskinson, J.; Stepick-Biek, P.N.; Araujo, F.G.; Thulliez, P.; Suzuki, Y.; Remington, J.S. Toxoplasma antigens recognized by immunoglobulin G subclasses during acute and chronic infection. J. Clin. Microbiol. 1989, 27, 2031–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canedo-Solares, I.; Galván-Ramirez, M.L.; Luna-Pastén, H.; Pérez, L.R.R.; Ortiz-Alegria, L.B.; Rico-Torres, C.P.; Vela-Amieva, M.; Pérez-Andrade, M.; Figueroa-Damián, R.; Dolores Correa, D. Congenital toxoplasmosis: Specific IgG subclasses in mother/newborn pairs. Pediatr. Infect. Dis. J. 2008, 27, 469–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, L.M.; Rangel, A.L.P.; Peixe, R.G.; Silva-dos-Santos, P.P.; Lemos, E.M.; Martins-Filho, O.A.; Bahia-Oliveira, L.M.G. Specific IgM, IgG and IgG1 directed against Toxoplasma gondii detected by flow cytometry and their potential as serologic tools to support clinical indirect fundoscopic presumed diagnosis of ocular disease. J. Immunol. Methods 2015, 417, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Buffolano, W.; Beghetto, E.; Del Pezzo, M.; Spadoni, A.; Di Cristina, M.; Petersen, E.; Gargano, N. Use of recombinant antigens for early postnatal diagnosis of congenital toxoplasmosis. J. Clin. Microbiol. 2005, 43, 5916–5924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparoni, A.; Avanzini, A.; Ravagni Probizer, F.; Chirico, G.; Rondini, G.; Severi, F. IgG subclasses compared in maternal and cord serum and breast milk. Arch. Dis. Child. 1992, 67, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Camejo, C.; Puyol, A.; Fazio, L.; Rodríguez, A.; Villamil, E.; Andina, E.; Cordobez, V.; Díaz, H.; Lemos, M.; Siré, G.; et al. Antibody profile of colostrum and the effect of processing in human milk banks: Implications in immunoregulatory properties. J. Hum. Lact. 2018, 34, 137–147. [Google Scholar] [CrossRef]
- Canedo-Solares, I.; Gomez-Chavez, F.; Luna-Pasten, H.; Ortiz-Alegria, L.B.; Flores-Garcia, Y.; Figueroa-Damian, R.; Macedo-Romero, C.A.; Correa, D. What do anti-Toxoplasma gondii IgA and IgG subclasses in human saliva indicate? Parasite Immunol. 2018, 40, e12526. [Google Scholar] [CrossRef]
- Santana, S.S.; Silva, D.A.O.; Vaz, L.D.; Pirovani, C.P.; Barros, G.B.; Lemos, E.M.; Dietze, R.; Mineo, J.R.; Cunha-Junior, J.P. Analysis of IgG subclasses (IgG1 and IgG3) to recombinant SAG2A protein from Toxoplasma gondii in sequential serum samples from patients with toxoplasmosis. Immunol. Lett. 2012, 143, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Souza-e-Silva, C.H.; Vasconcelos-Santos, D.V.; Andrade, G.Q.; Carellos, E.V.M.; Romanelli, R.M.C.; Resende, L.M.; Januário, J.N.; Carneiro, M.; Carneiro, A.C.A.V.; Vitor, R.W.A. Association between IgG subclasses against Toxoplasma gondii and clinical signs in newborns with congenital toxoplasmosis. Pediatr. Infect. Dis. J. 2013, 32, 13–16. [Google Scholar] [CrossRef]
- Spiegelberg, H.L. Biological activities of immunoglobulins of different classes and subclasses. Adv. Immunol. 1974, 19, 259–294. [Google Scholar] [PubMed]
- Hamilton, R.G. Human IgG subclass measurements in the clinical laboratory. Clin. Chem. 1987, 33, 1707–1725. [Google Scholar] [CrossRef] [PubMed]
- Andrade, G.Q.; Januário, J.N.; Carneiro, A.C.A.V.; Carneiro, M.; Vasconcelos-Santos, D.V.; Vitor, R.W.A. Early diagnosis of congenital toxoplasmosis in newborn infants using IgG subclasses against two Toxoplasma gondii recombinant proteins. Mem. Inst. Oswaldo Cruz 2012, 107, 342–347. [Google Scholar]
- Aalberse, R.C.; van der Gaag, R.; van Leeuwen, J. Serologic aspects of IgG4 antibodies. I. Prolonged immunization results in an IgG4-restricted response. J. Immunol. 1983, 130, 722–726. [Google Scholar] [PubMed]
- Pastorino, A.C.; Jacob, C.M.A.; Grumach, A.S. Subclasses de IgG e sua importância em pediatria. Pediatria 1994, 16, 60–66. [Google Scholar]
- Stone, J.H.; Zen, Y.; Deshpande, V. IgG4-related disease. N. Engl. J. Med. 2012, 366, 539–551. [Google Scholar] [CrossRef]
- Correa, D.; Canedo-Solares, I.; Ortiz-Alegria, L.B.; Caballero-Ortega, H.; Rico-Torres, C.P. Congenital and acquired toxoplasmosis: Diversity and role of antibodies in different compartments of the host. Parasite Immunol. 2007, 29, 651–660. [Google Scholar] [CrossRef]
- Nirula, A.; Glaser, S.M.; Kalled, S.L.; Taylor, F.R. What is IgG4? A review of the biology of a unique immunoglobulin subtype. Curr. Opin. Rheumatol. 2011, 23, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Meira, C.S.; Vidal, J.E.; Costa-Silva, T.A.; Motoie, G.; Gava, R.; Hiramoto, R.M.; Pereira-Chioccola, V.L. IgG4 specific to Toxoplasma gondii excretory/secretory antigens in serum and/or cerebrospinal fluid support the cerebral toxoplasmosis diagnosis in HIV-infected patients. J. Immunol. Methods. 2013, 395, 21–28. [Google Scholar] [CrossRef]
- Kim, K.; Keller, M.A.; Heiner, D.C. Immunoglobulin G subclasses in human colostrum, milk and saliva. Acta Paediatr. 1992, 81, 113–118. [Google Scholar] [CrossRef]
- Mehta, P.D.; Mehta, S.P.; Isaacs, C.E. Distribution of IgG subclasses in human colostrum and milk. Immunol. Lett. 1989, 22, 235–238. [Google Scholar] [CrossRef]
- Guglietta, S.; Beghetto, E.; Spadoni, A.; Buffolano, W.; Del Porto, P.; Gargano, N. Age-dependent impairment of functional helper T cell responses to immunodominant epitopes of Toxoplasma gondii antigens in congenitally infected individuals. Microbes Infect. 2007, 9, 127–133. [Google Scholar] [CrossRef]
- Carlier, Y.; Truyens, C.; Deloron, P.; Peyron, F. Congenital parasitic infections: A review. Acta Trop. 2012, 121, 55–70. [Google Scholar] [CrossRef]
- Redline, R.W. Inflammatory responses in the placenta and umbilical cord. Semin. Fetal Neonatal Med. 2006, 11, 296–301. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum n (%) | Colostrum n (%) | ||||||
|---|---|---|---|---|---|---|---|
| Association of Isotypes IgG/IgM/IgA | IgG1 | IgG3 | IgG4 | IgG1 | IgG3 | IgG4 | |
| +/+/+ | n = 2 | 2 (100) | 2 (100) | 2 (100) | 1 (50.0) | 2 (100) | 2 (100) |
| +/+/− | n = 10 | 10 (100) | 5 (50.0) | 4 (40.0) | 7 (70.0) | 9 (90.0) | 3 (30.0) |
| +/−/− | n = 111 | 109 (98.2) | 59 (53.2) | 50 (44.1) | 60 (54.1) | 85 (76.6) | 39 (35.1) |
| Total | 121 (98.4) | 66 (53.6) | 56 (45.5) | 68 (55.2) | 96 (78.0) | 44 (35.8) |
| Chronic Phase | Early Acute Phase 0–3 Months of Infection | Convalescent Phase 3–12 Months of Infection | |
|---|---|---|---|
| IgG+ | IgG3/IgG1 > 1 IgM+IgA+ or IgM+IgA− | IgG3/IgG1 < 1 IgM+IgA+ or IgM+IgA− | |
| Serum n (%) | 117 (90.0%) | 1 (0.77%) | 12 (9.23%) |
| Colostrum n (%) | 115 (88.46%) | 6 (4.62%) | 9 (6.92%) |
| SERUM | COLOSTRUM | |||||||
|---|---|---|---|---|---|---|---|---|
| Case | IgG3/IgG1 Ratio | IgM (EI) | IgA (EI) | Diagnosis Status | IgG3/IgG1 Ratio | IgM (EI) | IgA (EI) | Diagnosis Status |
| 1 | 0.31 | Neg (0.59) | Neg (0.55) | Chronic infection | 0.14 | Pos (1.26) | Neg (0.78) | Convalescent phase 3–12 months of infection |
| 2 | 0.25 | Pos (1.58) | Neg (0.67) | Convalescent phase 3–12 months of infection | 0.43 | Pos (1.32) | Neg (0.82) | Convalescent phase 3–12 months of infection |
| 3 | 1.16 | Pos (3.14) | Neg (0.69) | Early acute infection 0–3 months of infection | 0.88 | Pos (1.35) | Neg (0.99) | Convalescent phase 3–12 months of infection |
| 4 | 0.24 | Pos (3.36) | Neg (1.01) | Convalescent phase 3–12 months of infection | 2.86 | Pos (3.11) | Neg (0.81) | Early acute infection 0–3 months of infection |
| 5 | 0.75 | Pos (2.76) | Pos (1.85) | Convalescent phase 3–12 months of infection | 7.07 | Pos (3.86) | Pos (1.82) | Early acute infection 0–3 months of infection |
| 6 | 0.21 | Pos (1.93) | Neg (1.07) | Convalescent phase 3–12 months of infection | 0.64 | Pos (1.28) | Neg (0.71) | Convalescent phase 3–12 months of infection |
| 7 | 0.34 | Pos (2.29) | Pos (2.77) | Convalescent phase 3–12 months of infection | 0.29 | Pos (1.24) | Pos (1.30) | Convalescent phase 3–12 months of infection |
| 8 | 0.55 | Pos (1.62) | Neg (0.64) | Convalescent phase 3–12 months of infection | 0.50 | Pos (1.52) | Neg (0.83) | Convalescent phase 3–12 months of infection |
| 9 | 0.15 | Pos (3.30) | Neg (1.12) | Convalescent phase 3–12 months of infection | 1.03 | Pos (1.82) | Neg (0.82) | Early acute infection 0–3 months of infection |
| 10 | 0.12 | Neg (0.63) | Neg (0.86) | Chronic infection | 0.32 | Pos (1.94) | Neg (0.82) | Convalescent phase 3–12 months of infection |
| 11 | 0.63 | Pos (1.66) | Neg (0.88) | Convalescent phase 3–12 months of infection | 1.23 | Pos (1.29) | Neg (0.86) | Early acute infection 0–3 months of infection |
| 12 | 0.12 | Pos (2.18) | Neg (0.77) | Convalescent phase 3–12 months of infection | 0.28 | Pos (1.22) | Neg (0.86) | Convalescent phase 3–12 months of infection |
| 13 | 0.27 | Pos (2.28) | Neg (1.01) | Convalescent phase 3–12 months of infection | 3.11 | Pos (1.61) | Neg (0.90) | Early acute infection 0–3 months of infection |
| 14 | 0.19 | Neg (1.18) | Neg (0.88) | Chronic infection | 0.43 | Pos (1.35) | Neg (0.72) | Convalescent phase 3–12 months of infection |
| 15 | 0.30 | Pos (1.91) | Neg (0.87) | Convalescent phase 3–12 months of infection | 4.02 | Pos (1.25) | Neg (0.58) | Early acute infection 0–3 months of infection |
| 16 | 0.12 | Pos (2.15) | Neg (1.08) | Convalescent phase 3–12 months of infection | 0.29 | Neg (0.97) | Neg (0.91) | Chronic infection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borges, H.D.S.; Oliveira-Scussel, A.C.M.; Oliveira, Â.M.M.; Abdallah, V.O.S.; Pajuaba, A.C.A.M.; Mineo, J.R. Comparative Detection of Immunoglobulin Isotypes and Subclasses against Toxoplasma gondii Soluble Antigen in Serum and Colostrum Samples from Puerperal Women. Int. J. Environ. Res. Public Health 2022, 19, 7953. https://doi.org/10.3390/ijerph19137953
Borges HDS, Oliveira-Scussel ACM, Oliveira ÂMM, Abdallah VOS, Pajuaba ACAM, Mineo JR. Comparative Detection of Immunoglobulin Isotypes and Subclasses against Toxoplasma gondii Soluble Antigen in Serum and Colostrum Samples from Puerperal Women. International Journal of Environmental Research and Public Health. 2022; 19(13):7953. https://doi.org/10.3390/ijerph19137953
Chicago/Turabian StyleBorges, Hellen Dayane Silva, Ana Carolina Morais Oliveira-Scussel, Ângela Maria Morais Oliveira, Vânia Olivetti Steffen Abdallah, Ana Cláudia Arantes Marquez Pajuaba, and José Roberto Mineo. 2022. "Comparative Detection of Immunoglobulin Isotypes and Subclasses against Toxoplasma gondii Soluble Antigen in Serum and Colostrum Samples from Puerperal Women" International Journal of Environmental Research and Public Health 19, no. 13: 7953. https://doi.org/10.3390/ijerph19137953