
Resistance to Arsenite and Arsenate in Saccharomyces cerevisiae Arises through the Subtelomeric Expansion of a Cluster of Yeast Genes

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. Comparative Genomic Hybridization (CGH)
2.3. Strain Genome Sequencing, Analysis, and Assembly
2.4. Identification of Potential Duplications through Whole-Genome Sequencing
2.5. Pulsed-Field Gel Electrophoresis and Southern Blotting Hybridization
2.6. Transcriptional Analysis
3. Results
3.1. Resistance to Arsenite and Arsenate in Natural Saccharomyces cerevisiae Strains
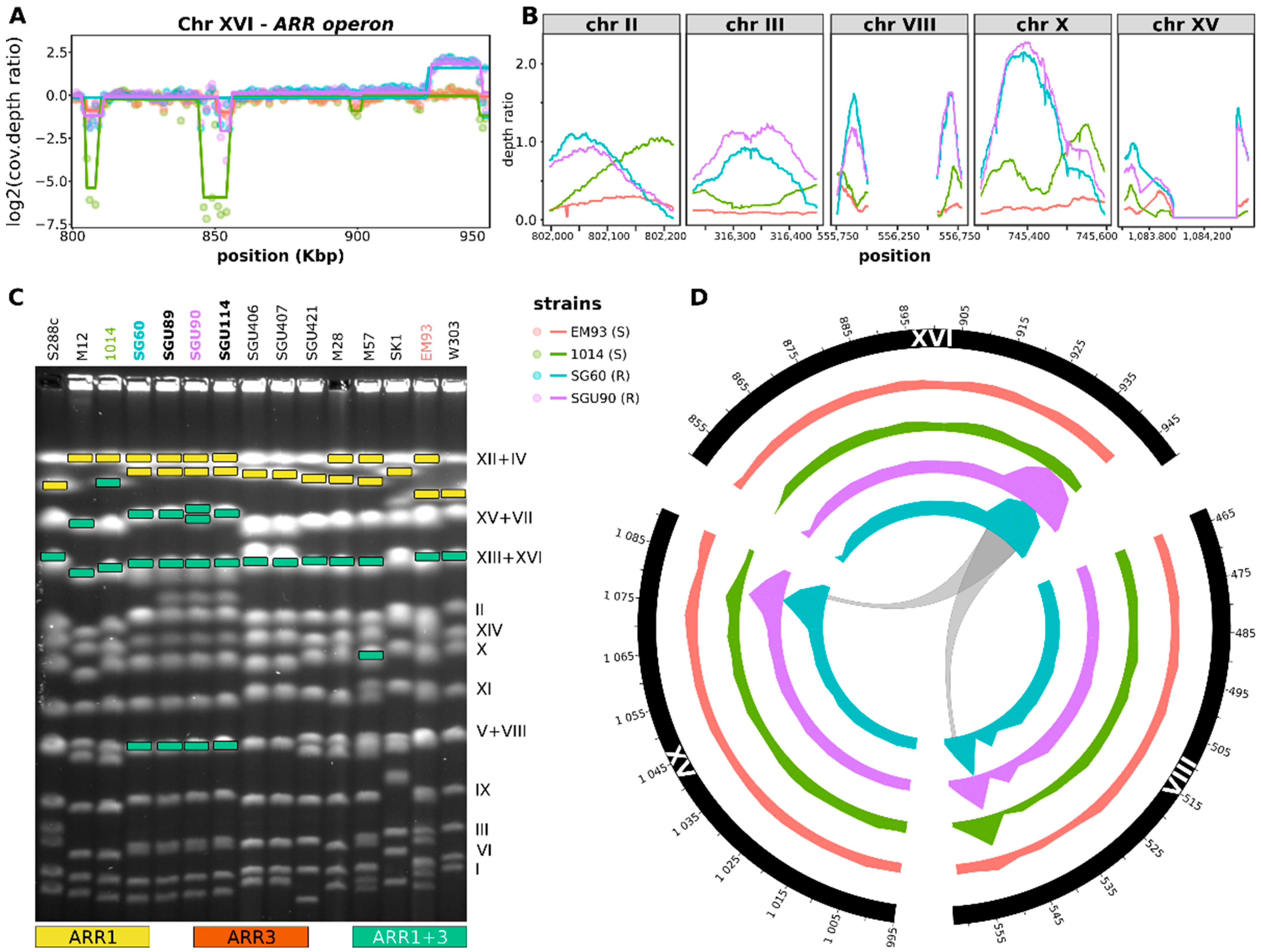
3.2. Comparative Genomic Hybridization (CGH) Reveals the Presence of Multiple Copies of the ARR1, ARR2, and ARR3 Genes in Arsenic-Resistant Strains
3.3. Transcriptomic Profiling Highlights Patterns Associated with the Genetic Makeup of Arsenic-Resistant Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duker, A.; Carranza, E.; Hale, M. Arsenic Geochemistry and Health. Environ. Int. 2005, 31, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Bowell, R.J.; Alpers, C.N.; Jamieson, H.E.; Nordstrom, D.K.; Majzlan, J. The Environmental Geochemistry of Arsenic—An Overview. Rev. Miner. Geochem. 2014, 79, 1–16. [Google Scholar] [CrossRef]
- World Health Organization. WHO Exposure to Lead: A Major Public Health Concern. Available online: https://www.who.int/ipcs/features/arsenic.pdf?ua=1 (accessed on 1 December 2021).
- Watson, W.H. Molecular Mechanisms in Arsenic Toxicity. In Advances in Molecular Toxicology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 35–75. [Google Scholar]
- Islam, K.; Wang, Q.Q.; Naranmandura, H. Molecular Mechanisms of Arsenic Toxicity. In Advances in Molecular Toxicology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 77–107. [Google Scholar]
- Kitchin, K.T.; Wallace, K. The Role of Protein Binding of Trivalent Arsenicals in Arsenic Carcinogenesis and Toxicity. J. Inorg. Biochem. 2008, 102, 532–539. [Google Scholar] [CrossRef]
- Wu, J.; Tisa, L.S.; Rosen, B.P. Membrane Topology of the ArsB Protein, the Membrane Subunit of an Anion-Translocating ATPase. J. Biol. Chem. 1992, 267, 12570–12576. [Google Scholar] [CrossRef]
- Gladysheva, T.B.; Oden, K.L.; Rosen, B.P. Properties of the Arsenate Reductase of Plasmid R773. Biochemistry 1994, 33, 7288–7293. [Google Scholar] [CrossRef] [PubMed]
- Tisa, L.S.; Rosen, B.P. Molecular Characterization of an Anion Pump. The ArsB Protein Is the Membrane Anchor for the ArsA Protein. J. Biol. Chem. 1990, 265, 190–194. [Google Scholar] [CrossRef]
- Takemaru, K.-I.; Mizuno, M.; Sato, T.; Takeuchi, M.; Kobayashi, Y. Complete Nucleotide Sequence of a Skin Element Excised by DNA Rearrangement during Sporulation in Bacillus Subtilis. Microbiology 1995, 141, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Diorio, C.; Cai, J.; Marmor, J.; Shinder, R.; DuBow, M.S. An Escherichia Coli Chromosomal Ars Operon Homolog Is Functional in Arsenic Detoxification and Is Conserved in Gram-Negative Bacteria. J. Bacteriol. 1995, 177, 2050–2056. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.; Silver, S. Regulation and Expression of the Arsenic Resistance Operon from Staphylococcus Aureus Plasmid PI258. J. Bacteriol. 1992, 174, 3684–3694. [Google Scholar] [CrossRef] [Green Version]
- Rosenstein, R.; Peschel, A.; Wieland, B.; Götz, F. Expression and Regulation of the Antimonite, Arsenite, and Arsenate Resistance Operon of Staphylococcus Xylosus Plasmid PSX267. J. Bacteriol. 1992, 174, 3676–3683. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, C.; Ji, G.; Ramirez, J.; Silver, S. Resistance to Arsenic Compounds in Microorganisms. FEMS Microbiol. Rev. 1994, 15, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Bobrowicz, P.; Wysocki, R.; Owsianik, G.; Goffeau, A.; Ułaszewski, S. Isolation of Three Contiguous Genes, ACR1, ACR2 and ACR3, Involved in Resistance to Arsenic Compounds in the Yeast Saccharomyces Cerevisiae. Yeast 1997, 13, 819–828. [Google Scholar] [CrossRef]
- Kumar, N.V.; Yang, J.; Pillai, J.K.; Rawat, S.; Solano, C.; Kumar, A.; Grøtli, M.; Stemmler, T.L.; Rosen, B.P.; Tamás, M.J. Arsenic Directly Binds to and Activates the Yeast AP-1-like Transcription Factor Yap8. Mol. Cell. Biol. 2016, 36, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, R.; Rosen, B.P. Saccharomyces Cerevisiae ACR2 Gene Encodes an Arsenate Reductase. FEMS Microbiol. Lett. 1998, 168, 127–136. [Google Scholar] [CrossRef]
- Maciaszczyk-Dziubinska, E.; Migocka, M.; Wysocki, R. Acr3p Is a Plasma Membrane Antiporter That Catalyzes As(III)/H+ and Sb(III)/H+ Exchange in Saccharomyces Cerevisiae. Biochim. Biophys. Acta-Biomembr. 2011, 1808, 1855–1859. [Google Scholar] [CrossRef] [Green Version]
- Chwastowski, J.; Chwastowski, H. The kinetic reduction of Cr(VI) by yeast Saccharomyces cerevisiae, Phaffia rhodozyma and their protoplasts. Acta Biochim. Pol. 2013, 60, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Dymond, J.S. Chapter Twelve—Preparation of Genomic DNA from Saccharomyces cerevisiae. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 529, pp. 153–160. [Google Scholar] [CrossRef]
- Pollack, J.R.; Perou, C.M.; Sorlie, T.; Alizadeh, A.A.; Rees, C.; Eise, M.B.; Pergamenschikov, A.; Williams, C.F.; van de Rijn, M.; Jeffrey, S.S.; et al. Genome-Wide Analysis of DNA Copy Number Variation in Breast Cancer Using DNA Microarrays. Nat. Genet. 1999, 23, 69. [Google Scholar] [CrossRef]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A Toolkit for Quality Control of next Generation Sequencing Data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce Framework for Analyzing next-Generation DNA Sequencing Data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A Program for Annotating and Predicting the Effects of Single Nucleotide Polymorphisms, SnpEff. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackman, S.D.; Vandervalk, B.P.; Mohamadi, H.; Chu, J.; Yeo, S.; Hammond, S.A.; Jahesh, G.; Khan, H.; Coombe, L.; Warren, R.L.; et al. ABySS 2.0: Resource-Efficient Assembly of Large Genomes Using a Bloom Filter. Genome Res. 2017, 27, 768–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, C.B.; Jackman, S.D.; Birol, I.; Jones, S.J.M. ABySS-Explorer: Visualizing Genome Sequence Assemblies. IEEE Trans. Vis. Comput. Graph. 2009, 15, 881–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanke, M.; Waack, S. Gene Prediction with a Hidden Markov Model and a New Intron Submodel. Bioinformatics 2003, 19, ii215–ii225. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Venkatraman, E.S.; Olshen, A.B. A Faster Circular Binary Segmentation Algorithm for the Analysis of Array CGH Data. Bioinformatics 2007, 23, 657–663. [Google Scholar] [CrossRef] [Green Version]
- Hage, A.E.; Houseley, J. Resolution of Budding Yeast Chromosomes Using Pulsed-Field Gel Electrophoresis. In DNA Electrophoresis; Humana Press: Totowa, NJ, USA, 2013; pp. 195–207. [Google Scholar]
- Louis, E.J. Whole Chromosome Analysis. In Methods in Microbiology: Yeast Gene Analysis; Tuite, M.F., Brown, A.J.P., Eds.; Academic Press: London, UK, 1998; pp. 15–31. [Google Scholar]
- Townsend, J.P.; Cavalieri, D.; Hartl, D.L. Population genetic variation in genome-wide gene expression. Mol. Biol. Evol. 2003, 20, 955–963. [Google Scholar] [CrossRef]
- Mortimer, R.K.; Johnston, J.R. Genealogy of principal strains of the yeast genetic stock center. Genetics 1986, 113, 35–43. [Google Scholar] [CrossRef]
- Kane, S.M.; Roth, R. Carbohydrate metabolism during ascospore development in yeast. J. Bacteriol. 1974, 118, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Stern, M.; Jensen, R.; Herskowitz, I. Five SWI genes are required for expression of the HO gene in yeast. J. Mol. Biol. 1984, 178, 853–868. [Google Scholar] [CrossRef]
- Winston, F.; Dollard, C.; Ricupero-Hovasse, S.L. Construction of a set of convenient Saccharomyces cerevisiae strains that are isogenic to S288C. Yeast 1995, 11, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Stefanini, I.; Dapporto, L.; Legras, J.L.; Calabretta, A.; Di Paola, M.; De Filippo, C.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S.; et al. Role of social wasps in Saccharomyces cerevisiae ecology and evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef] [Green Version]
- Cavalieri, D.; Barberio, C.; Casalone, E.; Pinzauti, F.; Sebastiani, F.; Mortimer, R.; Polsinelli, M. Genetic and molecular diversity in Saccharomyces cerevisiae natural populations. Food Technol. Biotechnol. 1998, 36, 45–50. [Google Scholar]
- Ewing, A.D. Transposable Element Detection from Whole Genome Sequence Data. Mob. DNA 2015, 6, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klemm, S.L.; Shipony, Z.; Greenleaf, W.J. Chromatin Accessibility and the Regulatory Epigenome. Nat. Rev. Genet. 2019, 20, 207–220. [Google Scholar] [CrossRef]
- Rathod, J.; Tu, H.-P.; Chang, Y.-I.; Chu, Y.-H.; Tseng, Y.-Y.; Jean, J.-S.; Wu, W.-S. YARG: A Repository for Arsenic-Related Genes in Yeast. PLoS ONE 2018, 13, e0201204. [Google Scholar] [CrossRef]
- Garelick, H.; Jones, H.; Dybowska, A.; Valsami-Jones, E. Arsenic Pollution Sources. In Reviews of Environmental Contamination; Springer: Cham, Switzerland, 2009; pp. 17–60. [Google Scholar]
- Garbinski, L.D.; Rosen, B.P.; Chen, J. Pathways of Arsenic Uptake and Efflux. Environ. Int. 2019, 126, 585–597. [Google Scholar] [CrossRef]
- Knight, S.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional Microbial Signatures Positively Correlate with Differential Wine Phenotypes: Evidence for a Microbial Aspect to Terroir. Sci. Rep. 2015, 5, 14233. [Google Scholar] [CrossRef] [Green Version]
- Akache, B.; Turcotte, B. New Regulators of Drug Sensitivity in the Family of Yeast Zinc Cluster Proteins. J. Biol. Chem. 2002, 277, 21254–21260. [Google Scholar] [CrossRef] [Green Version]
- Abou Zeid, A.Z. Production of Cycloheximide by Streptomyces sp. Acta Microbiol. Pol. B 1972, 4, 83–88. [Google Scholar]
- Haugen, A.C.; Kelley, R.; Collins, J.B.; Tucker, C.J.; Deng, C.; Afshari, C.A.; Brown, J.M.; Ideker, T.; Van Houten, B. Integrating Phenotypic and Expression Profiles to Map Arsenic-Response Networks. Genome Biol. 2004, 5, R95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menezes, R.A.; Amaral, C.; Delaunay, A.; Toledano, M.; Rodrigues-Pousada, C. Yap8p Activation in Saccharomyces Cerevisiae under Arsenic Conditions. FEBS Lett. 2004, 566, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Di, Y.; Tamás, M.J. Regulation of the Arsenic-Responsive Transcription Factor Yap8p Involves the Ubiquitin-Proteasome Pathway. J. Cell Sci. 2007, 120, 256–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.-X.; Li, J.; Aigrain, L.; Hallin, J.; Persson, K.; Oliver, K.; Bergström, A.; Coupland, P.; Warringer, J.; Lagomarsino, M.C.; et al. Contrasting Evolutionary Genome Dynamics between Domesticated and Wild Yeasts. Nat. Genet. 2017, 49, 913–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, S.; Fonseca, H.M.A.C.; Oliveira, S.M.R.; Oliveira, H.; Gupta, V.K.; Sharma, B.; de Lourdes Pereira, M. Environmental and health hazards of chromated copper arsenate-treated wood: A review. Int. J. Environ. Res. Public Health 2021, 18, 5518. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Strain | Sodium Arsenite [mM] | Sodium Arsenate [mM] | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1.25 | 2.5 | 5 | 10 | 15 | 6.25 | 12.5 | 25 | 50 | 75 | |
| 1014 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| BY4742 * | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| EM93 * | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| M12 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| M28 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| M57 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| SG60 | + | + | + | 0 | 0 | ++ | + | 0 | 0 | 0 |
| SGU89 | + | + | + | 0 | 0 | ++ | 0 | 0 | 0 | 0 |
| SGU90 | + | + | + | 0 | 0 | ++ | + | 0 | 0 | 0 |
| SGU114 | + | + | + | 0 | 0 | ++ | 0 | 0 | 0 | 0 |
| SGU406 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| SGU407 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| SK1 * | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| W303 * | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefanini, I.; Di Paola, M.; Liti, G.; Marranci, A.; Sebastiani, F.; Casalone, E.; Cavalieri, D. Resistance to Arsenite and Arsenate in Saccharomyces cerevisiae Arises through the Subtelomeric Expansion of a Cluster of Yeast Genes. Int. J. Environ. Res. Public Health 2022, 19, 8119. https://doi.org/10.3390/ijerph19138119
Stefanini I, Di Paola M, Liti G, Marranci A, Sebastiani F, Casalone E, Cavalieri D. Resistance to Arsenite and Arsenate in Saccharomyces cerevisiae Arises through the Subtelomeric Expansion of a Cluster of Yeast Genes. International Journal of Environmental Research and Public Health. 2022; 19(13):8119. https://doi.org/10.3390/ijerph19138119
Chicago/Turabian StyleStefanini, Irene, Monica Di Paola, Gianni Liti, Andrea Marranci, Federico Sebastiani, Enrico Casalone, and Duccio Cavalieri. 2022. "Resistance to Arsenite and Arsenate in Saccharomyces cerevisiae Arises through the Subtelomeric Expansion of a Cluster of Yeast Genes" International Journal of Environmental Research and Public Health 19, no. 13: 8119. https://doi.org/10.3390/ijerph19138119