
Comparison of Root Ecological Stoichiometry between Non-Growing Season and Growing Season of Grassland on the Chang Tang Plateau
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Collection and Analysis
2.3. Statistical Analyses
3. Results
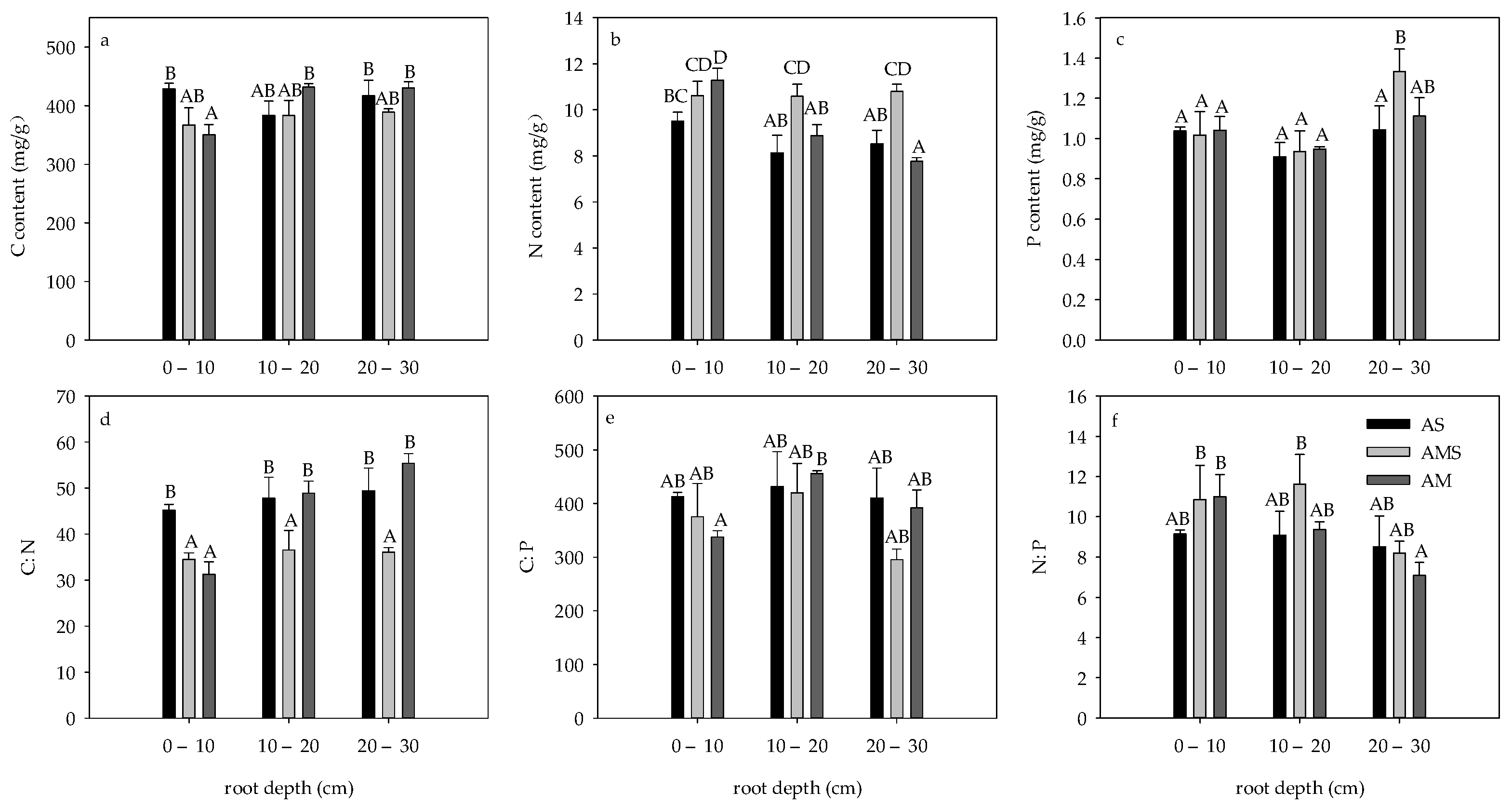
3.1. Contents and Ratios of C, N and P in Roots in April
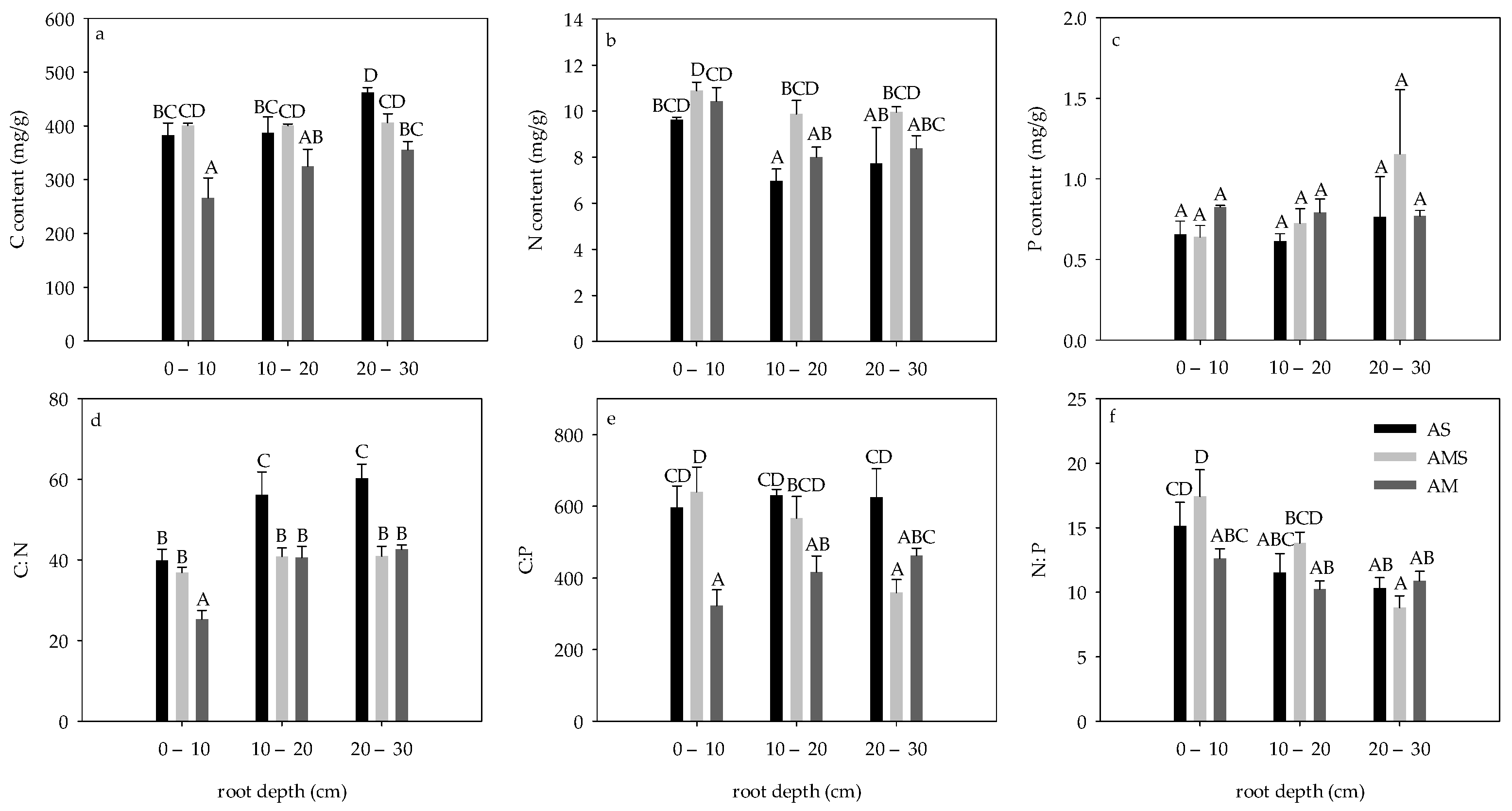
3.2. Contents and Ratios of C, N and P in Roots in August
3.3. Difference of Root C:N:P Stoichiometry between April and August
3.4. Patterns of C, N, P, AN and AP Contents of Soil
3.5. Relationship between Root C:N:P Stoichiometry and Soil Nutrients Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hans, S.; Felix, L.; Karl, A. Nutrient redistribution by grazing cattle drives patterns of topsoil N and P stocks in a low-input pasture ecosystem. Nutr. Cycl. Agroecosyst. 2010, 88, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Bardgett, R.D.; Rvander, W.; Jonsdottir, I.S.; Quirk, H.; Dutton, S. Temporal variability in plant and soil nitrogen pools in a high-Arctic ecosystem. Soil Biol. Biochem. 2007, 39, 2129–2137. [Google Scholar] [CrossRef]
- Yu, H.; Gao, R.Y.; Yang, W.J.; Yang, L.S.; Li, S.Y.; Lin, Y.M.; Wang, D.J.; Li, J. Carbon, nitrogen, and phosphorus contents of leaf, root, and soil and their relationships in dominant herbaceous plants in dry-hot valley. Chin. J. Appl. Environ. Biol. 2022, 28, 727–735. [Google Scholar] [CrossRef]
- Ellsworth, P.Z.; Sternberg, L.S.L. Linking soil nutrient availability, fine root production and turnover, and species composition in a seasonally dry plant community. Plant Soil. 2019, 442, 49–63. [Google Scholar] [CrossRef]
- Liu, Y.; He, J.W.; Yu, H.; Lin, Y.M.; Wang, D.J. Nutrients (C, N, P) contents and stoichiometric ratios of fine root, coarse root and leaf in dominant shrubs in dry-hot valley. Mt. Res. 2020, 38, 29–39. [Google Scholar] [CrossRef]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Wang, S.Q.; Yu, G.R. Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus elements. Acta Ecol. Sin. 2008, 28, 3937–3947. [Google Scholar] [CrossRef]
- Wu, J.S.; Shen, Z.X.; Zhang, X.Z.; Shi, P.L. Biomass allocation patterns of alpine grassland species and functional groups along a precipitation gradient on the Northern Tibetan Plateau. J. Mount. Sci. 2013, 10, 1097–1108. [Google Scholar] [CrossRef]
- Li, N.; Wang, G.X.; Yang, Y.; Gao, Y.H.; Liu, G.S. Plant production, and carbon and nitrogen source pools, are strongly intensified by experimental warming in alpine ecosystems in the Qinghai-Tibet Plateau. Soil Biol. Biochem. 2011, 43, 942–953. [Google Scholar] [CrossRef]
- Johnson, M.T.; Agrawal, A.A. Plant genotype and environment interact to shape a diverse arthropod community on evening primrose (Oenothera biennis). Ecology 2005, 86, 874–885. [Google Scholar] [CrossRef]
- Tian, L.; Zehao, Z.; Jingkuan, S.; Zhanyong, F.; Yinghan, Z.; Wenjing, X. Seasonal Variation Characteristics of C, N, and P Stoichiometry and Water Use Efficiency of Messerschmidia sibirica and Its Relationship With Soil Nutrients. Front. Ecol. Evol. 2022, 10, 1–11. [Google Scholar] [CrossRef]
- Du, C.; Gao, Y. Grazing exclusion alters ecological stoichiometry of plant and soil in degraded alpine grassland. Agric. Ecosyst. Environ. 2021, 308, 107256. [Google Scholar] [CrossRef]
- Lin, Z.L.; Gao, Y.; Zhu, T.X.; Gao, K. Effects of nitrogen application rate and density on the stoichiometric characteristics of C, N, and P in oat leaves. Pratac. Sci. 2020, 37, 1107–1114. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, J.; Yuan, X.; Zhu, B. Effects of warming on carbon and nitrogen cycling in alpine grassland ecosystems on the Tibetan Plateau: A meta-analysis. Geoderma 2020, 370, 114363. [Google Scholar] [CrossRef]
- Liu, C.; Li, W.; Xu, J.; Wei, W.; Xue, P.; Yan, H. Response of soil nutrients and stoichiometry to grazing management in alpine grassland on the Qinghai-Tibet Plateau. Soil Tillage Res. 2021, 206, 104822. [Google Scholar] [CrossRef]
- Hong, J.; Wang, X.; Wu, J. Effects of soil fertility on the N:P stoichiometry of herbaceous plants on a nutrient-limited alpine steppe on the northern Tibetan Plateau. Plant Soil 2015, 391, 179–194. [Google Scholar] [CrossRef]
- Shi, L.; Li, Q.; Fu, X.; Kou, L.; Dai, X.; Wang, H. Foliar, root and rhizospheric soil C:N:P stoichiometries of overstory and understory species in subtropical plantations. Catena 2021, 198, 105020. [Google Scholar] [CrossRef]
- Piao, S.; Tan, K.; Nan, H.; Ciais, P.; Fang, J.; Wang, T.; Vuichard, N.; Zhu, B. Impacts of climate and CO2 changes on the vegetation growth and carbon balance of Qinghai–Tibetan grasslands over the past five decades. Glob. Planet. Change 2012, 98–99, 73–80. [Google Scholar] [CrossRef]
- Wang, J.L.; Chang, T.J.; Li, P.; Cheng, H.H.; Fang, H.L. The vegetation carbon reserve and its spatial distribution configuration of grassland ecosystem in Tibet. Acta Ecol. Sin. 2009, 29, 931–938. [Google Scholar]
- Ma, X.X.; Yan, Y.; Lu, X.Y.; Wang, X.D. Dynamics of belowground biomass and its relationship with soil moisture in alpine grassland on the north Tibetan Plateau. Ecol. Environ. Sci. 2016, 25, 189–195. [Google Scholar] [CrossRef]
- Piao, S.L.; Fang, J.Y.; He, J.S.; Xiao, Y. Spatial Distribution of grassland biomass in China. Chin. J. Plant Ecol. 2004, 4, 491–498. [Google Scholar] [CrossRef]
- Cai, Y.J.; Wang, X.D.; Ding, W.X.; Tian, L.L.; Zhao, H.; Lu, X.Y. Potential short-term effects of yak and Tibetan sheep dung on greenhouse gas emissions in two alpine grassland soils under laboratory conditions. Biol. Fertil. Soils 2013, 49, 1215–1226. [Google Scholar] [CrossRef]
- Wang, H.Y.; Ding, R.; Wang, Z.H.; Yang, F.J. Effects of nitrogen and phosphorus addition on C: N:P ecological stoichi-ometry in leaves and roots of different canopy species in Hulun buir grassland. Acta Prataculturae Sin. 2020, 29, 37–45. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, E103–E122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.W.; Zhang, W.Y.; Zhong, H.P. Plant root N and P levels and their relationship to geographical and climate factors in a Chinese grassland transect. Acta Prataculturae Sin. 2014, 23, 69–76. [Google Scholar] [CrossRef]
- Zhao, W.; Reich, P.B.; Yu, Q.; Zhao, N.; Yin, C.; Zhao, C.; Li, D.; Hu, J.; Li, T.; Yin, H. Shrub type dominates the vertical distribution of leaf C: N: P stoichiometry across an extensive altitudinal gradient. Biogeosciences 2017, 15, 1–45. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhou, H.; Ma, Z. Phylogenetic structure of alpine steppe plant communities along a precipitation and temperature gradient on the Tibetan Plateau. Glob. Ecol. Conserv. 2020, 24, e01379. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, J.; Bai, W.; Zhang, Y.; Zhang, W.H.; Weiser, M. The response of root traits to precipitation change of herbaceous species in temperate steppes. Funct. Ecol. 2019, 33, 2030–2041. [Google Scholar] [CrossRef]
- Wang, J.L.; Zhong, Z.M.; Wang, Z.H.; Zhang, X.Z.; Shen, Z.X.; Hu, X.X. Distribution characteristics and its influence factors of vegetation C/N value of alpine steppe ecosystem in Tibetan Plateau. J. Plant Resour. Environ. 2013, 22, 20–28. [Google Scholar] [CrossRef]
- Tessier, J.T.; Raynal, D.J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Zhang, C.; Tian, H.; Liu, J.; Wang, S.; Liu, M.; Pan, S.; Shi, X. Pools and distributions of soil phosphorus in China. Glob. Biogeochem. Cycles 2005, 19, GB1020. [Google Scholar] [CrossRef] [Green Version]
- Braakhekke, W.G.; Hooftman, D. The resource balance hypothesis of plant species diversity in grassland. J. Veg. Sci. 1999, 10, 187–200. [Google Scholar] [CrossRef]
- Sardans, J.; Grau, O.; Chen, H.; Janssens, I.A.; Ciais, P.; Piao, S. Changes in nutrient concentrations of leaves and roots in response to global change factors. Glob. Change Biol. 2017, 23, 3849–3856. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Type | Depth (cm) | Root C | Root N | Root P | Root C:N | Root C:P | Root N:P |
|---|---|---|---|---|---|---|---|
| AS | 0–10 | 1.884 | −0.283 | 4.520 * | 1.741 | −3.049 | −3.219 * |
| 10–20 | −0.104 | 1.276 | 3.507 * | −1.150 | −2.965 * | −1.278 | |
| 20–30 | −1.632 | 0.476 | 1.005 | −1.775 | −2.202 | −1.035 | |
| AMS | 0–10 | −1.138 | −0.371 | 2.733 | −1.256 | −2.843 * | −2.442 |
| 10–20 | −0.666 | 0.900 | 1.543 | −0.901 | −1.806 | −1.286 | |
| 20–30 | −1.049 | 2.104 | 0.428 | −1.842 | −1.520 | −0.567 | |
| AM | 0–10 | 2.051 | 1.095 | 3.112 | 1.700 | 0.321 | −1.225 |
| 10–20 | 3.307 * | 1.359 | 1.846 | 2.167 | 0.861 | −1.180 | |
| 20–30 | 4.017 * | −1.058 | 3.526 * | 5.411 ** | −1.829 | −3.923 * |
| Month | Root C | Root N | Root P | Root C:N | Root C:P | Root N:P | |
|---|---|---|---|---|---|---|---|
| Soil TC | April | 0.112 | −0.133 | −0.160 | 0.147 | 0.178 | 0.012 |
| August | −0.197 | 0.162 | −0.055 | −0.226 | −0.098 | 0.129 | |
| Soil TN | April | −0.082 | 0.045 | −0.212 | −0.068 | 0.102 | 0.160 |
| August | −0.174 | 0.289 | −0.084 | −0.311 | 0.065 | 0.400 * | |
| Soil TP | April | −0.362 | 0.310 | −0.072 | −0.357 | −0.134 | 0.254 |
| August | −0.561 ** | 0.186 | −0.057 | −0.486 * | −0.315 | 0.107 | |
| Soil AN | April | −0.233 | 0.197 | −0.058 | −0.256 | −0.074 | 0.187 |
| August | −0.086 | 0.374 | −0.219 | −0.357 | 0.247 | 0.618 ** | |
| Soil AP | April | −0.139 | −0.003 | 0.024 | −0.047 | −0.142 | −0.038 |
| August | −0.489 * | 0.146 | −0.025 | −0.440 * | −0.151 | 0.262 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Yan, Y.; Hong, J.; Wang, X. Comparison of Root Ecological Stoichiometry between Non-Growing Season and Growing Season of Grassland on the Chang Tang Plateau. Int. J. Environ. Res. Public Health 2022, 19, 14628. https://doi.org/10.3390/ijerph192214628
Ma X, Yan Y, Hong J, Wang X. Comparison of Root Ecological Stoichiometry between Non-Growing Season and Growing Season of Grassland on the Chang Tang Plateau. International Journal of Environmental Research and Public Health. 2022; 19(22):14628. https://doi.org/10.3390/ijerph192214628
Chicago/Turabian StyleMa, Xingxing, Yan Yan, Jiangtao Hong, and Xiaodan Wang. 2022. "Comparison of Root Ecological Stoichiometry between Non-Growing Season and Growing Season of Grassland on the Chang Tang Plateau" International Journal of Environmental Research and Public Health 19, no. 22: 14628. https://doi.org/10.3390/ijerph192214628