
Mercury Sources, Emissions, Distribution and Bioavailability along an Estuarine Gradient under Semiarid Conditions in Northeast Brazil


Abstract
:1. Introduction
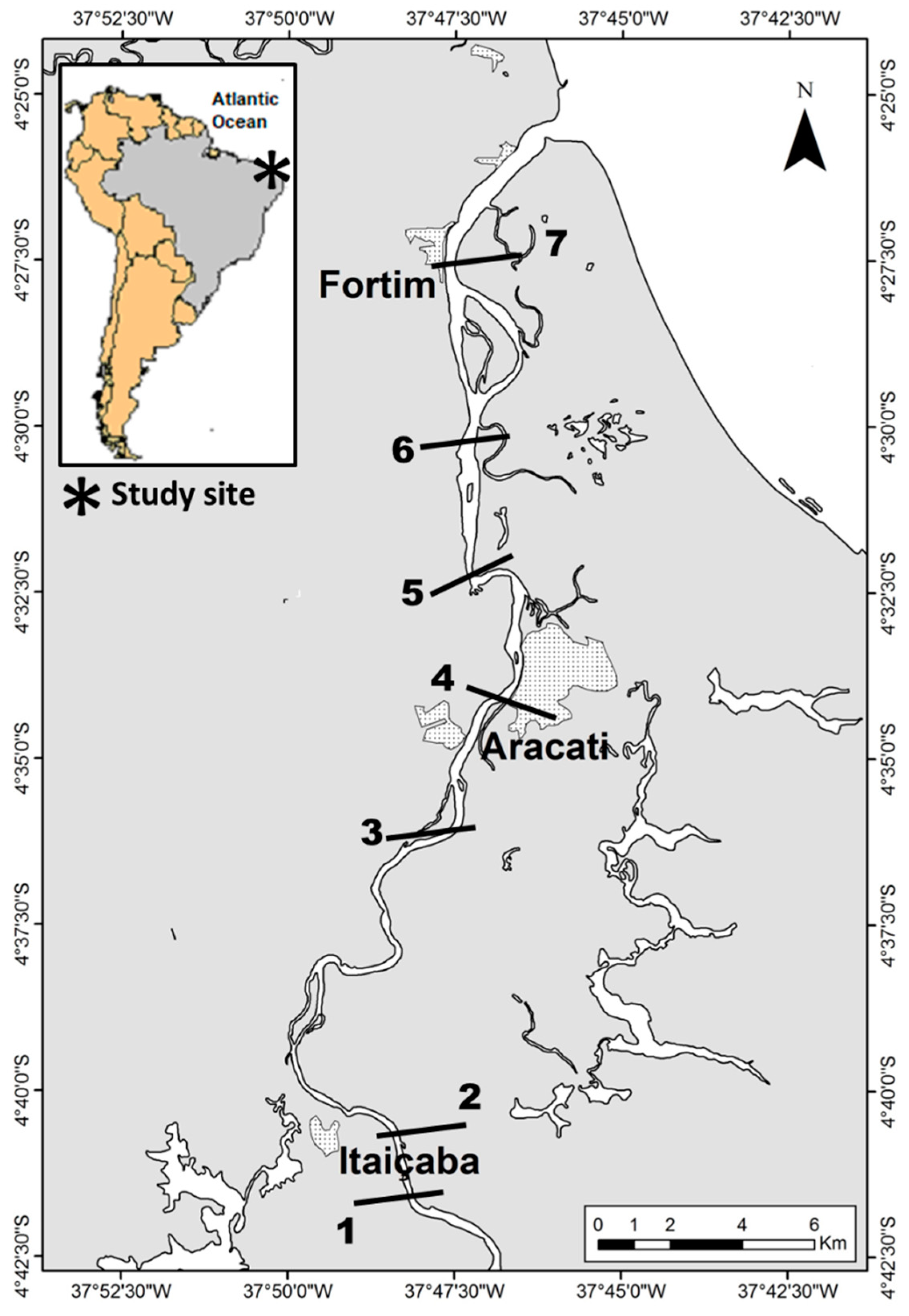
2. Material and Methods
3. Results and Discussion
3.1. Sources and Emissions of Hg
3.2. Hydrochemistry
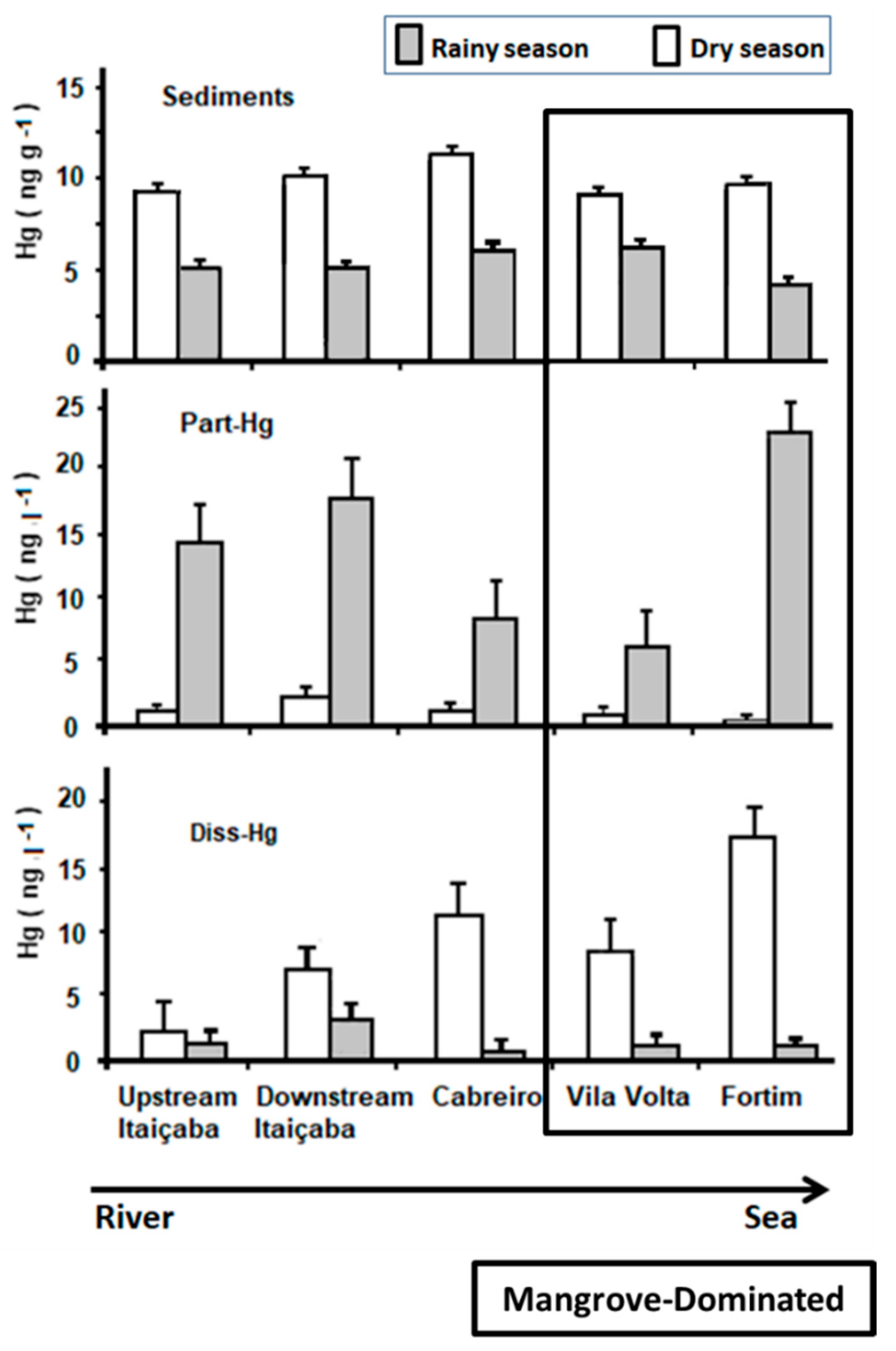
3.3. Mercury in Sediments and Water
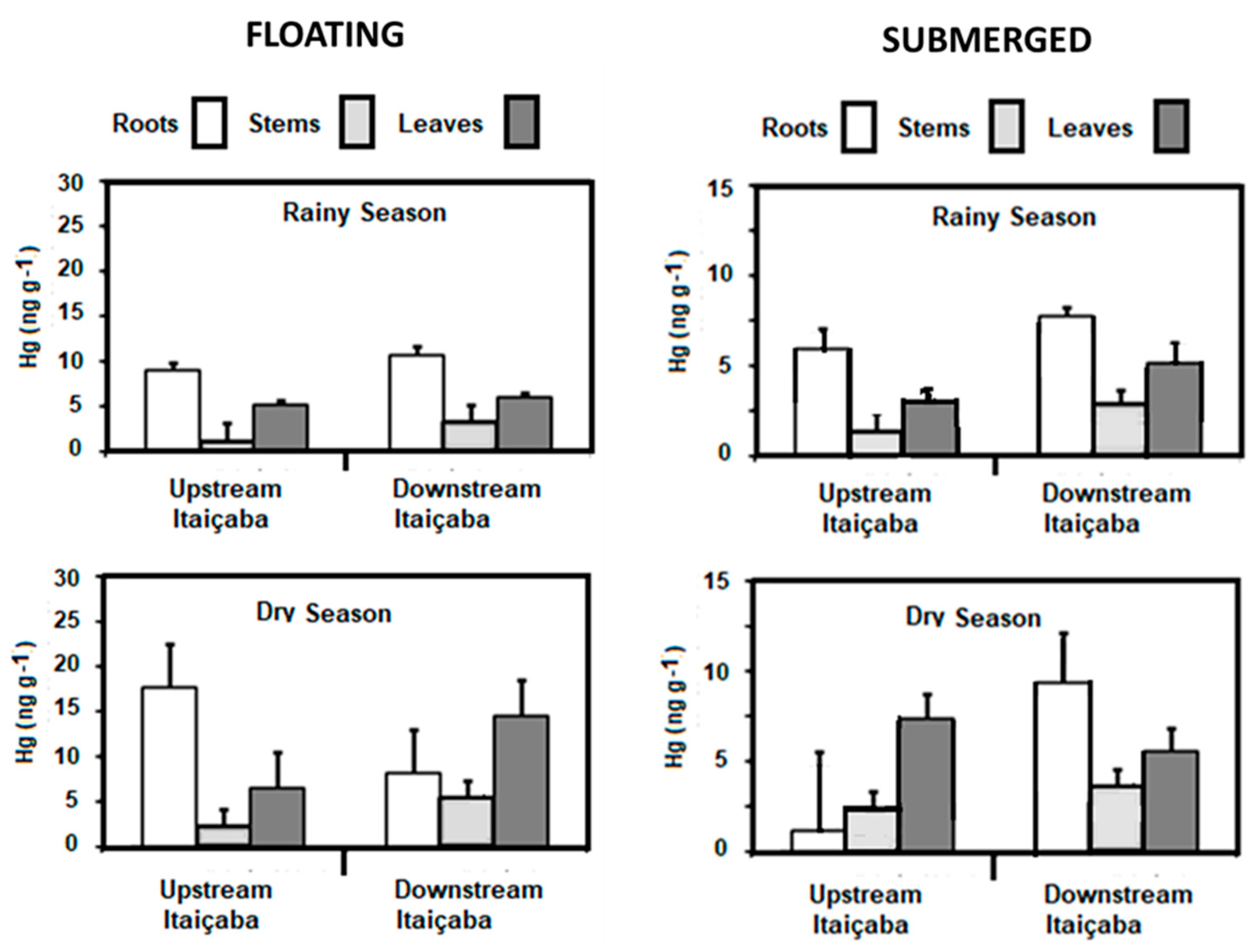
3.4. Mercury in Aquatic Macrophytes
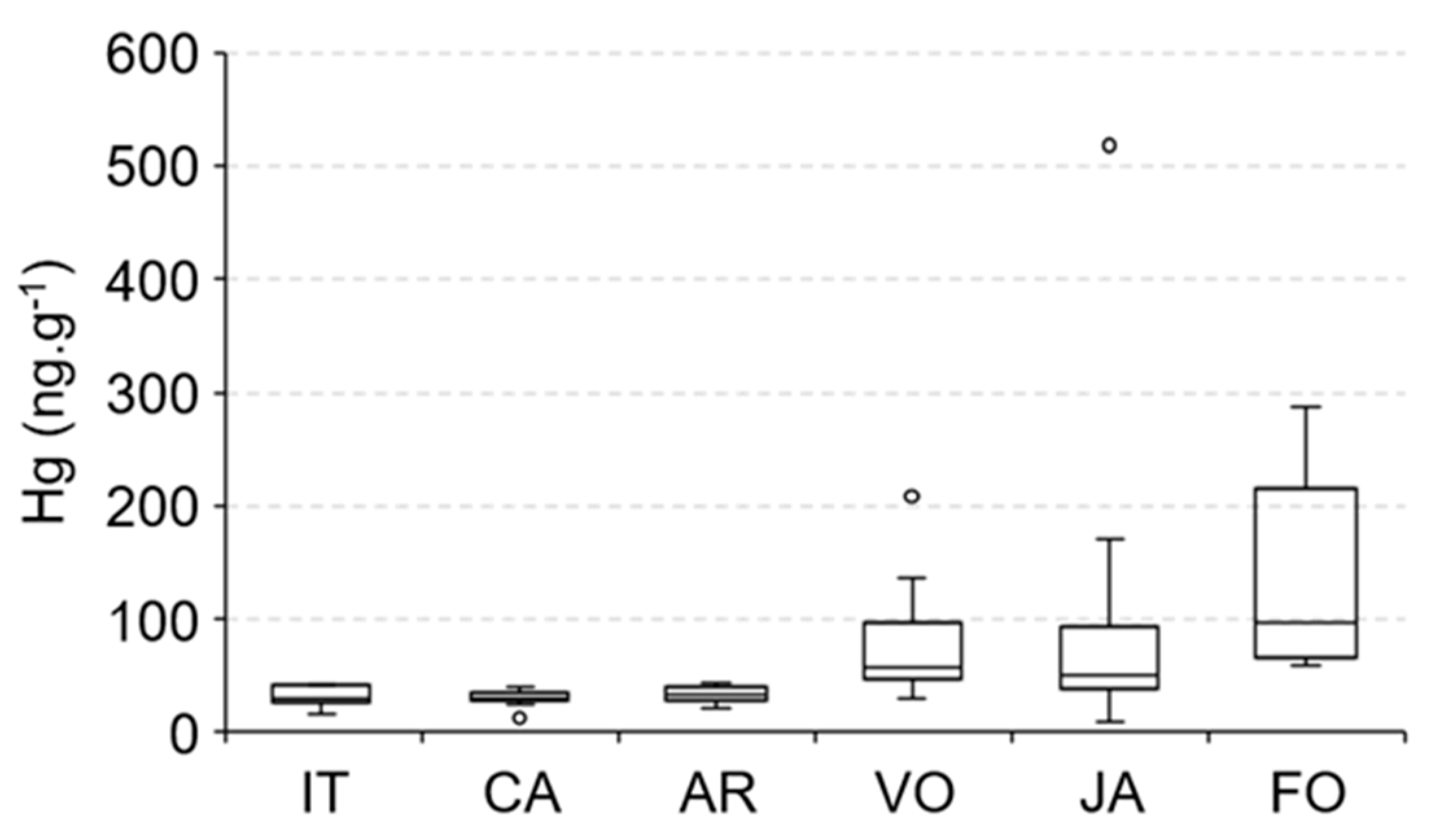
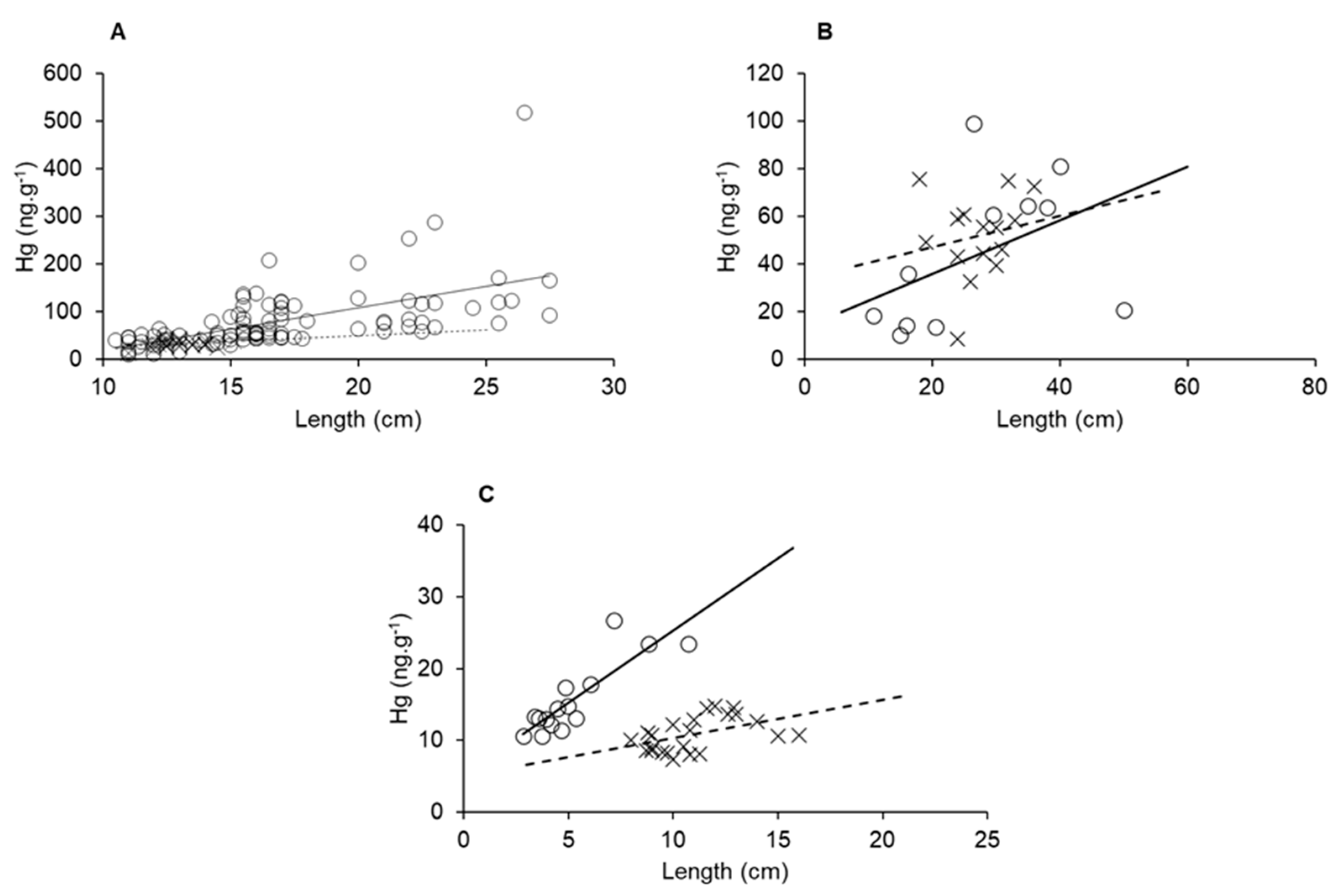
3.5. Mercury in Aquatic Fauna
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergamaschi, B.A.; Krabbenhoft, D.P.; Aiken, G.R.; Patino, E.; Rumbold, D.G.; Orem, W.H. Tidally driven export of dissolved organic carbon, total mercury, and methylmercury from a mangrove-dominated estuary. Environ. Sci. Technol. 2012, 46, 1371–1378. [Google Scholar] [CrossRef]
- Correia, R.R.S.; Guimarães, J.R.D. Mercury methylation and sulfate reduction rates in mangrove sediments, Rio de Janeiro, Brazil: The role of different microorganism consortia. Chemosphere 2017, 167, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, K.F.; Lacerda, L.D.; Peres, T.F.; Bezerra, M.F.; Dias, F.J.S. Emission factor and balance of mercury in fish farms in an artificial reservoir in NE-Brazil. Environ. Sci. Pollut. Res. 2015, 22, 18278–18287. [Google Scholar] [CrossRef] [PubMed]
- Stern, G.A.; Macdonald, R.W.; Outridge, P.M.; Wilson, S.; Chételat, J.; Cole, A.; Hintelmann, H.; Loseto, L.L.; Steffen, A.; Wang, F.; et al. How does climate change influence arctic mercury? Sci. Total Environ. 2012, 414, 22–42. [Google Scholar] [CrossRef]
- Lu, Y.; Yuan, J.; Lu, X.; Su, C.; Zhang, Y.; Wang, C.; Cao, X.; Li, Q.; Su, J.; Ittekkot, V.; et al. Major threats of pollution and climate change to global coastal. Sci. Total Environ. 2018, 239, 670–680. [Google Scholar] [CrossRef] [Green Version]
- Lacerda, L.D.; Marins, R.V.; Dias, F.J.S. An Arctic Paradox: Response of fluvial Hg inputs and its bioavailability to global climate change in an extreme coastal environment. Front. Earth Sci. 2020, 8, 93. [Google Scholar] [CrossRef]
- Dias, F.J.S.; Lacerda, L.D.; Marins, R.V.; de Paula, F.C.F. Comparative analysis of rating curve and ADP estimates of instantaneous water discharge through estuaries in two contrasting Brazilian rivers. Hydrol. Proc. 2011, 25, 2188–2201. [Google Scholar] [CrossRef]
- Lacerda, L.D.; Dias, F.J.S.; Marins, R.V.; Soares, T.M.; Godoy, J.M.; Godoy, M.D.P. Pluriannual watershed discharges of Hg into the tropical semi-arid estuary of the Jaguaribe River, NE Brazil. J. Braz. Chem. Soc. 2013, 24, 1719–1731. [Google Scholar] [CrossRef]
- Alongi, D.M. The impact of climate change on mangrove forests. Curr. Clim. Change Rep. 2015, 1, 30–39. [Google Scholar] [CrossRef]
- Godoy, M.D.P.; Lacerda, L.D. Mangroves response to climate change: A review of recent findings on mangrove extension and distribution. An. Acad. Brasil. Ciênc. 2015, 87, 651–667. [Google Scholar] [CrossRef]
- Ward, R.D.; Lacerda, L.D. Response of mangrove ecosystems to sea level change. In Dynamic Sedimentary Environments of Mangrove Coasts; Sidik, F., Friess, D.A., Eds.; Elsevier Publ. Co.: Amsterdan, The Netherlands, 2021; pp. 235–253. [Google Scholar]
- Saniewska, D.; Beldowska, M.; Beldowski, J.; Saniewski, M.; Szubska, M.; Romanowski, A.; Falkowska, L. The impact of land use and season on the riverine transport of mercury into the marine coastal zone. Environ. Monit. Assess. 2014, 186, 7593–7604. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, L.S.; Pestana, I.A.; Rocha, A.R.M.; Meneguelli-Souza, A.C.; Lima, C.A.I.; Almeida, M.G.; Bastos, W.R.; Souza, C.M.M. Drought promotes increases in total mercury and methylmercury concentrations in fish from the lower Paraíba do Sul River, southeastern Brazil. Chemosphere 2018, 202, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, L.D.; Ward, R.; Borges, R.; Ferreira, A.C. Mangrove trace metal biogeochemistry response to global climate change. Front. For. Glob. Change 2022, 5, 817992. [Google Scholar] [CrossRef]
- Pei, L.; Zhong, H.; Duana, D.; Pana, K. A review on mercury biogeochemistry in mangrove sediments: Hotspots of methylmercury production? Sci. Total Environ. 2019, 680, 140–150. [Google Scholar] [CrossRef]
- Godoy, M.D.P.; Meireles, A.J.A.; Lacerda, L.D. Mangrove response to land use change in estuaries along the semiarid coast of Ceará, Brazil. J. Coast. Res. 2018, 34, 524–533. [Google Scholar] [CrossRef]
- Alvala, R.C.S.; Cunha, A.P.; Brito, S.S.B.; Seluchi, M.E.; Marengo, J.A.; Moraes, O.L.L. Drought monitoring in the Brazilian Semiarid region. An. Acad. Bras. Ciênc. 2019, 91, e20170209. [Google Scholar] [CrossRef] [Green Version]
- Marengo, J.A.; Alves, L.M.; Alvala, R.C.S.; Cunha, A.P.; Brito, S.; Moraes, O.L.L. Climatic characteristics of the 2010–2016 drought in the semiarid Northeast Brazil region. An. Acad. Bras. Ciênc. 2018, 90, 1973–1985. [Google Scholar] [CrossRef]
- Costa, B.G.B.; Soares, T.M.; Torres, R.F.; Lacerda, L.D. Mercury distribution in a mangrove tidal creek affected by intensive shrimp farming. Bull. Environ. Cont. Toxicol. 2013, 90, 537–541. [Google Scholar] [CrossRef]
- Costa, B.G.B.; Lacerda, L.D. Mercury (Hg) in fish consumed by the local population of the Jaguaribe River Lower Basin, Northeast Brazil. Environ. Sci. Pollut. Res. 2014, 21, 13335–13341. [Google Scholar] [CrossRef]
- Moura, V.L.; Lacerda, L.D. Contrasting mercury bioavailability in the marine and fluvial dominated areas of the Jaguaribe River, Ceará, Brazil. Bull. Environ. Contamin. Toxicol. 2018, 101, 49–54. [Google Scholar] [CrossRef]
- Rios, J.H.L.; Marins, R.V.; Oliveira, K.F.; Lacerda, L.D. Long-Term (2002–2015) changes in mercury contamination in NE Brazil depicted by the mangrove oyster Crassostraea rhizophorae (Guilding, 1828). Bull. Environ. Contam. Toxicol. 2016, 97, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.K.; Tripathi, B.D.; Kim, K.H. Removal and accumulation of mercury by aquatic macrophytes from an open cast coal mine effluent. J. Hazard Mater. 2009, 172, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Lafabrie, C.; Major, K.M.; Major, C.S.; Cebrián, J. Trace metal contamination of the aquatic plant Hydrilla verticillata and associated sediment in a coastal Alabama creek (Gulf of Mexico—USA). Mar. Pollut. Bull. 2013, 68, 147–151. [Google Scholar] [CrossRef]
- Tong, Y.; Zhang, W.; Hu, D.; Ou, L.; Hu, X.; Yang, T.; Wei, W.; Ju, L.; Wang, X. Behavior of mercury in an urban river and its accumulation in aquatic plants. Environ. Earth Sci. 2013, 68, 1089–1097. [Google Scholar] [CrossRef]
- Tong, Y.; Zhang, W.; Chen, C.; Chen, L.; Wang, W.; Hu, X.; Wang, H.; Hu, D.; Ou, L.; Wang, X.; et al. Fate modeling of mercury species and fluxes estimation in an urban river. Environ. Pollut. 2014, 184, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Sitarska, M.; Traczewska, S.; Stanicka-Łotocka, A.; Filyarovskaya, V.; Zamorska-Wojdyła, D. Accumulation of mercury in the biomass of selected pleustophytes. Environ. Protect. Engineer. 2014, 40, 165–174. [Google Scholar] [CrossRef]
- Paraquetti, H.H.M.; Ayres, G.A.; Molisani, M.M.; Lacerda, L.D. Mercury distribution, speciation and flux in the Sepetiba Bay tributaries, SE Brazil. Water Res. 2004, 38, 1439–1448. [Google Scholar] [CrossRef]
- Paula Filho, F.J.; Marins, R.V.; Lacerda, L.D.; Aguiar, J.E.; Peres, T.F. Background values for evaluation of heavy metal contamination in sediments in the Parnaíba River Delta estuary, NE/Brazil. Mar. Pollut. Bull. 2015, 91, 424–428. [Google Scholar] [CrossRef]
- Paula Filho, F.J.; Marins, R.V.; Lacerda, L.D. Natural and anthropogenic emissions of N and P to the Parnaíba River Delta in NE Brazil. Estuar. Coast. Shelf Sci. 2015, 166, 34–44. [Google Scholar] [CrossRef]
- Oliveira, D.C.M.; Correia, R.R.S.; Marinho, C.C.; Guimarães, J.R.D. Mercury methylation in sediments of a Brazilian mangrove under different vegetation covers and salinities. Chemosphere 2015, 127, 214–221. [Google Scholar] [CrossRef]
- Dai, A.; Qian, T.; Trenberth, K.E. Changes in continental freshwater discharge from 1948 to 2004. J. Clim. 2009, 22, 2273–2792. [Google Scholar] [CrossRef]
- Dias, F.J.S.; Castro, B.M.; Lacerda, L.D.; Miranda, L.B.; Marins, R.V. Physical characteristics and discharges of suspended particulate matter at the continent-ocean interface in an estuary located in a semiarid region in northeastern Brazil. Estuar. Coast. Shelf Sci. 2016, 180, 258–274. [Google Scholar] [CrossRef]
- Dias, F.J.S.; Castro, B.M.; Lacerda, L.D. Tidal and low-frequency currents off the Jaguaribe River estuary (4oS, 37o4′W), northeastern Brazil. Ocean Dyn. 2018, 68, 967–985. [Google Scholar] [CrossRef]
- Bloom, N.S.; Crecelius, E.C. Determination of mercury in seawater at sub-nanogram per liter levels. Mar. Chem. 1983, 14, 49–59. [Google Scholar] [CrossRef]
- Marins, R.V.; Paraquetti, H.H.M.; Ayres, G.A. Alternativa analítica para a especiação físico-química de mercúrio em águas costeiras tropicais. Quím. Nova 2002, 25, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Sial, A.N.; Lacerda, L.D.; Ferreira, V.P.; Frei, R.; Marquillas, R.A.; Barbosa, J.A.; Gaucher, C.; Windmöller, C.C.; Pereira, N.S. Mercury as a proxy for volcanic activity during extreme environmental turnover: The Cretaceous-Paleogene transition. Paleogeogr Palaeoclim. Palaeoecol. 2013, 387, 153–164. [Google Scholar] [CrossRef]
- Lacerda, L.D.; Sena, D.D. Estimativas de Cargas de Nitrogênio, Fósforo e Metais Pesados de Interesse Ambiental para as Bacias Inferiores do Litoral do Estado do Ceará. Secretaria de Meio Ambiente do Ceará, Programa Zoneamento Ecológico e Econômico do Litoral do Ceará, Fortaleza, Brazil. 2005. Available online: https://www.semace.gov.ce/zee/produtos/EstimativasCargas.pdf (accessed on 20 January 2022).
- Lacerda, L.D.; Soares, T.M.; Costa, B.G.B.; Godoy, M.D.P. Mercury Emission factors from intensive shrimp aquaculture and its relative importance to the Jaguaribe River Estuary, NE Brazil. Bull. Environ. Cont. Toxicol. 2011, 87, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Marins, R.V.; Lacerda, L.D.; Mounier, S.; Paraquetti, H.H.M.; Marques, W.S. Caracterização hidroquímica, distribuição e especiação de mercúrio nos estuários dos Rios Ceará e Pacotí, Região Metropolitana de Fortaleza, Ceará, Brasil. Geochim. Bras. 2002, 16, 37–48. Available online: https://www.geobrasiliensis.org.br/geobrasiliensis/article/view/497/pdf (accessed on 24 April 2022).
- Marins, R.V.; Paula Filho, F.J.; Maia, S.R.R.; Lacerda, L.D.; Marques, W.S. Distribuição de mercúrio total como indicador de poluição urbana e industrial na costa brasileira. Quím. Nova 2004, 27, 763–770. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.C.; Lacerda, L.D.; Morais, J.O. Spatial and temporal variability of hydrographic parameters measured in the estuary of the Jaguaribe river (semi-arid region, northeast, Brazil). Rev. Bras. Geofís. 2012, 30, 245–261. [Google Scholar] [CrossRef] [Green Version]
- Dias, F.J.S.; Marins, R.V.; Maia, L.P. Hydrology of a well-mixed estuary at the semi-arid Northeastern Brazilian coast. Acta Limnol. Brasil. 2009, 21, 377–385. [Google Scholar]
- Dias, F.J.S.; Marins, R.V.; Maia, L.P. Impact of drainage basin changes on suspended matter and particulate copper and zinc discharges to the ocean from the Jaguaribe River in the semiarid NE Brazilian coast. J. Coast. Res. 2013, 29, 1137–1145. [Google Scholar] [CrossRef]
- Marins, R.V.; Paula Filho, F.J.; Eschrique, A.S.; Lacerda, L.D. Anthropogenic sources and distribution of phosphorus in sediments from in the Jaguaribe river estuary, NE Brazil. Braz. J. Biol. 2011, 71, 673–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marins, R.V.; Paula Filho, F.J.; Rocha, C.A.S. Phosphorus geochemistry as a proxy of environmental estuarine processes at the Jaguaribe River, northeastern Brazil. Quím. Nova 2007, 30, 1208–1214. [Google Scholar] [CrossRef] [Green Version]
- Marins, R.V.; Lacerda, L.D.; Araújo, I.C.S.; Fonseca, L.V.; Silva, F.A.T.F. Phosphorus and suspended matter retention in mangroves affected by shrimp farm effluents in NE Brazil. An. Acad. Bras. Ciênc. 2020, 92, e20200758. [Google Scholar] [CrossRef]
- Vaisman, A.G.; Marins, R.V.; Lacerda, L.D. Characterization of the mangrove oyster, Crassostrea rhizophorae, as a bio-monitor for mercury in tropical estuarine systems, Northeast Brazil. Bull. Environ. Cont. Toxicol. 2005, 74, 582–588. [Google Scholar] [CrossRef]
- Doile, T.W.; Krauss, K.W.; Conner, W.H.; From, A.S. Predicting the retreat and migration of tidal forests along the northern Gulf of Mexico under sea-level rise. For. Ecol. Manag. 2010, 259, 770–777. [Google Scholar] [CrossRef]
- Mounier, S.J.; Lacerda, L.D.; Marins, R.V. Determining the influence of urbanization on mangrove zones of Northeastern Brazil: Characterization of Ceará State coastal zone organic matter inputs. In Threats to Mangrove Forests; Makowski, C., Ed.; Springer: Berlin, Germany, 2018; pp. 199–222. [Google Scholar]
- Coulibaly, S.; Atse, B.C.; Koffi, K.M.; Sylla, S.; Konan, K.J.; Kouassi, N.J. Seasonal accumulations of some heavy metal in water, sediment and tissues of Black Chinned Tilapia Sarotherodon melanotheron from Bietri Bay in Ebrie Lagoon, Ivory Coast. Bull. Environ. Cont. Toxicol. 2012, 128, 12–522. [Google Scholar] [CrossRef]
- Cardoso, P.G.; Pereira, E.; Duarte, A.C.; Azeiteiro, U.M. Temporal characterization of mercury accumulation at different trophic levels and implications for metal biomagnification along a coastal food web. Mar. Pollut. Bull. 2014, 87, 39–47. [Google Scholar] [CrossRef]
- Jiang, T.; Skyllberg, U.; Björn, E.; Green, N.W.; Tang, J.H.; Wang, D.Y.; Gao, J.; Li, C.X. Characteristics of dissolved organic matter (DOM) and relationship with dissolved mercury in Xiaoqing River-Laizhou Bay estuary, Bohai Sea, China. Environ. Pollut. 2017, 223, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molisani, M.M.; Rocha, R.; Machado, W.; Barreto, R.C.; Lacerda, L.D. Mercury contents in aquatic macrophytes from two reservoirs in the Paraíba do Sul: Guandú river system, SE Brazil. Braz. J. Biol. 2006, 66, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Abbas, Z.; Rizwan, M.; Zaheer, I.E.; Yavaş, İ.; Ünay, A.; Abdel-DAIM, M.M.; Bin-Jumah, M.; Hasanuzzaman, M.; Kalderis, D. Application of floating aquatic plants in phytoremediation of heavy metals polluted water: A review. Sustainability 2020, 12, 1927. [Google Scholar] [CrossRef]
- Marins, R.V.; Lacerda, L.D.; Gonçalves, G.O.; Paiva, E.C. Effects of root metabolism on the post-depositional mobilization of mercury in salt marsh soils. Bull. Environ. Contam. Toxicol. 1997, 58, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, Z.M. Mercury toxicity, molecular response and tolerance in higher plants. Biometals 2012, 25, 847–857. [Google Scholar] [CrossRef]
- Wang, Y.D.; Greger, M. Clonal difference in mercury tolerance, accumulation and distribution in willow. J. Environm. Qual. 2004, 33, 1779–1785. [Google Scholar] [CrossRef]
- Barletta, M.; Lucena, L.R.R.; Costa, M.F.; Barbosa-Cintra, S.C.T.; Cysneros, F.J.A. The interaction rainfall vs. weight as determinant of total mercury concentration in fish from a tropical estuary. Environ. Pollut. 2012, 167, 1–6. [Google Scholar] [CrossRef]
- Lacerda, L.D.; Marins, R.V.; Cavalcante, M. Mercury mobilization due to global climate and regional land use changes in the Jaguaribe River Estuary, NE Brazil. In Vulnerabilidad de las zonas costeras de Latinoamérica al cambio climático; Botello, A.V., Villanueva, S., Gutiérrez, J., Galaviz, J.L.R., Eds.; UNAM, Mexico D.F.: Ciudad de México, México, 2017; pp. 333–344. [Google Scholar]
- Chennuri, K.; Chakraborty, P.; Jayachandran, S.; Mohakud, S.K.; Ishita, I.; Ramtek, D.; Padalkar, P.P.; Babu, P.C.; Babu, K.R. Operationally defined mercury (Hg) species can delineate Hg bioaccumulation in mangrove sediment systems: A case study. Sci. Total Environ. 2020, 701, 134842. [Google Scholar] [CrossRef]
- Bank, M.S.; Chesney, E.; Shine, J.P.; Maage, A.; Senn, D.B. Mercury bioaccumulation and trophic transfer in sympatric snapper species from the Gulf of Mexico. Ecol. Appl. 2007, 17, 2100–2110. [Google Scholar] [CrossRef]
- Sackett, D.K.; Cope, W.G.; Rice, J.A.; Aday, D.D. The Influence of Fish Length on Tissue Mercury Dynamics: Implications for Natural Resource Management and Human Health Risk. Int. J. Environ. Res. Publ. Health 2013, 10, 638–659. [Google Scholar] [CrossRef] [Green Version]
- Razavi, N.R.; Cushman, S.F.; Halfman, J.D.; Massey, T. Mercury bioaccumulation in stream food webs of the Finger Lakes in central New York State, USA. Ecotoxicol. Environ. Safety 2019, 172, 265–272. [Google Scholar] [CrossRef]
- Julio, T.G.; Moura, V.L.; Lacerda, L.D.; Lessa, R.P.T. Mercury concentrations in coastal Elasmobranchs (Hypanus guttatus and Rhizoprionodon porosus) and human exposure in Pernambuco, Northeastern Brazil. An. Acad. Bras. Ciênc. 2022, 94, e20220045. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sediments | Diss-Hg | Part-Hg | |
|---|---|---|---|
| Rainy season | 6.14 ± 0.41 | 1.12 ± 0.28 | 7.46 ± 2.07 |
| Dry season | 9.94 ± 0.40 | 12.32 ± 2.63 | 1.10 ± 0.44 |
| Student’s t p-Value | 0.001 * | 0.003 * | 0.017 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, V.L.; de Lacerda, L.D. Mercury Sources, Emissions, Distribution and Bioavailability along an Estuarine Gradient under Semiarid Conditions in Northeast Brazil. Int. J. Environ. Res. Public Health 2022, 19, 17092. https://doi.org/10.3390/ijerph192417092
Moura VL, de Lacerda LD. Mercury Sources, Emissions, Distribution and Bioavailability along an Estuarine Gradient under Semiarid Conditions in Northeast Brazil. International Journal of Environmental Research and Public Health. 2022; 19(24):17092. https://doi.org/10.3390/ijerph192417092
Chicago/Turabian StyleMoura, Victor Lacerda, and Luiz Drude de Lacerda. 2022. "Mercury Sources, Emissions, Distribution and Bioavailability along an Estuarine Gradient under Semiarid Conditions in Northeast Brazil" International Journal of Environmental Research and Public Health 19, no. 24: 17092. https://doi.org/10.3390/ijerph192417092
APA StyleMoura, V. L., & de Lacerda, L. D. (2022). Mercury Sources, Emissions, Distribution and Bioavailability along an Estuarine Gradient under Semiarid Conditions in Northeast Brazil. International Journal of Environmental Research and Public Health, 19(24), 17092. https://doi.org/10.3390/ijerph192417092