

Sustained Cytotoxic Response of Peripheral Blood Mononuclear Cells from Unvaccinated Individuals Admitted to the ICU Due to Critical COVID-19 Is Essential to Avoid a Fatal Outcome
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Ethical Statement
2.3. Cells
2.4. Pseudotyped SARS-CoV-2 Infection Assay
2.5. Flow Cytometry Analysis
2.6. Detection of SARS-CoV-2 RNAemia
2.7. Statistical Analysis
3. Results
3.1. Characteristics of the Participants
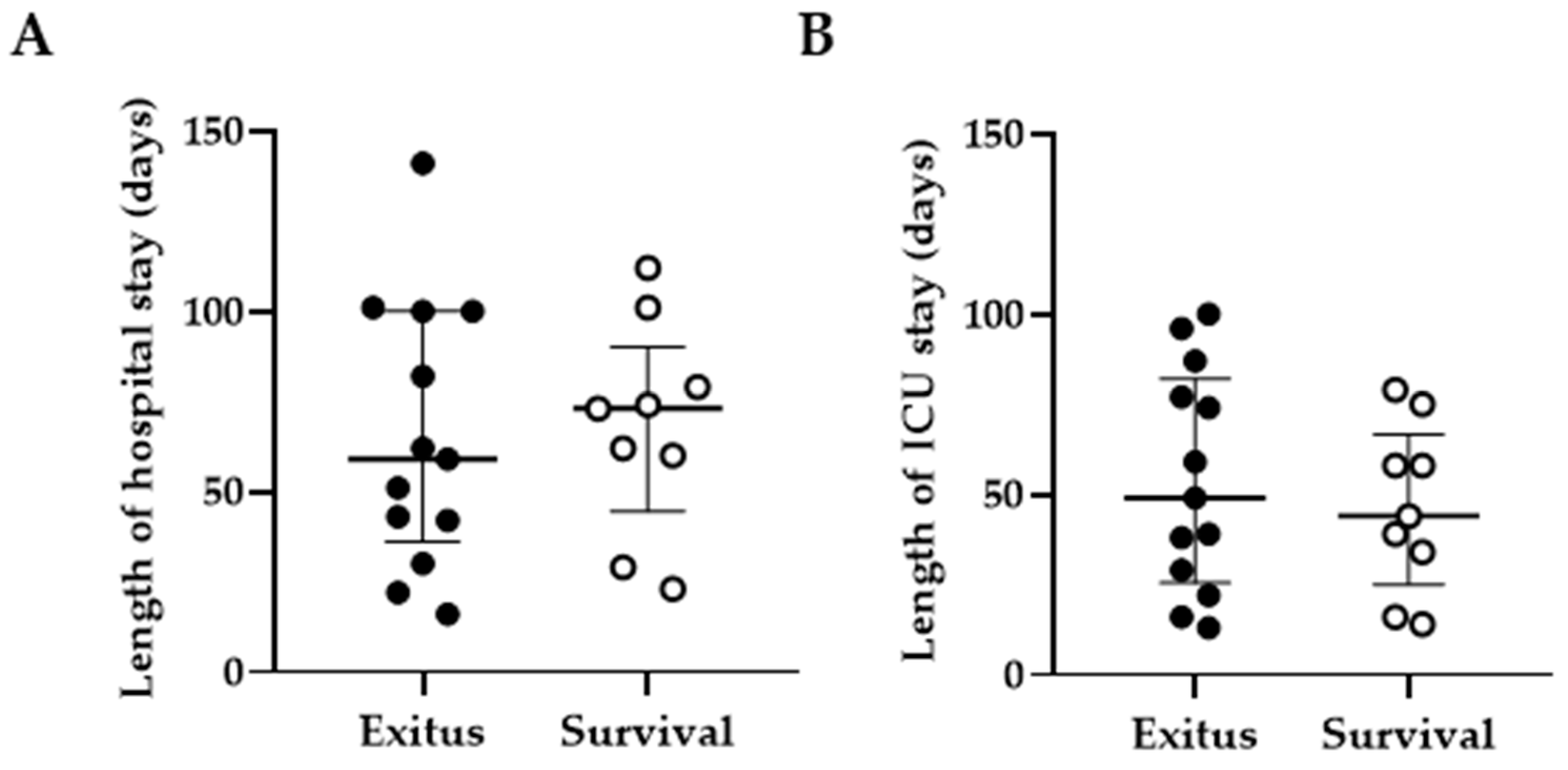
3.2. Length of Hospital and ICU Stay
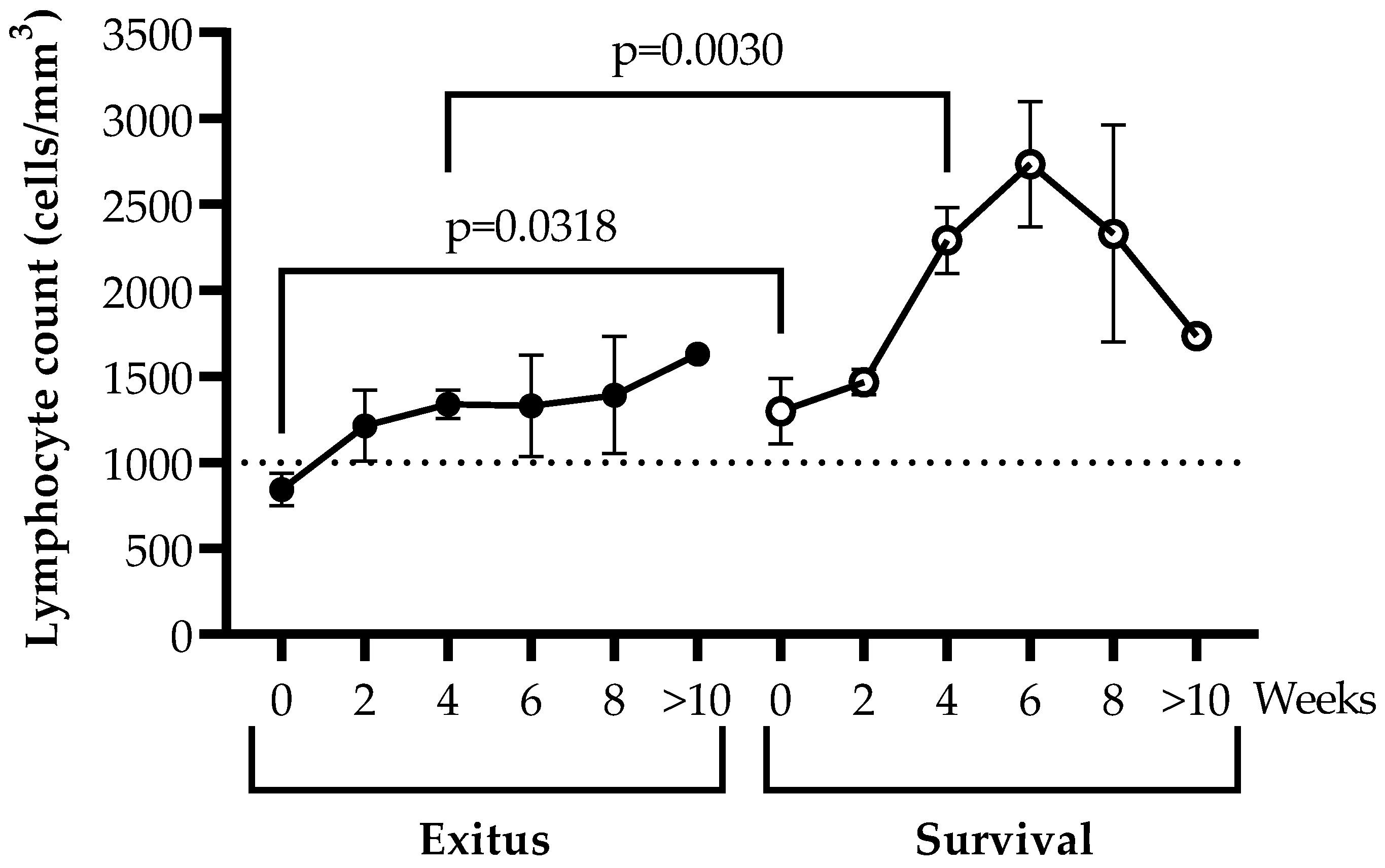
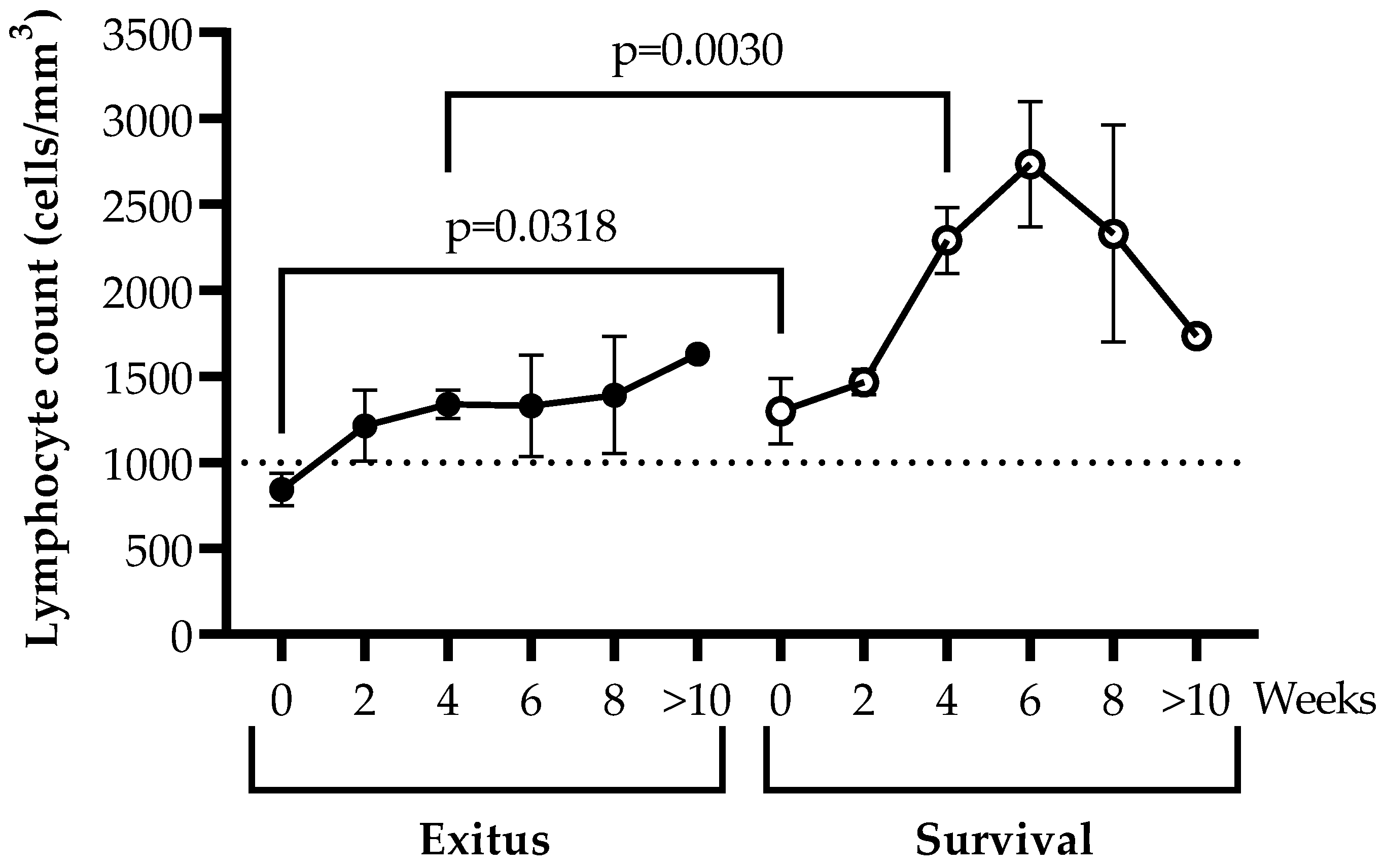
3.3. Levels of Peripheral Blood Lymphocytes
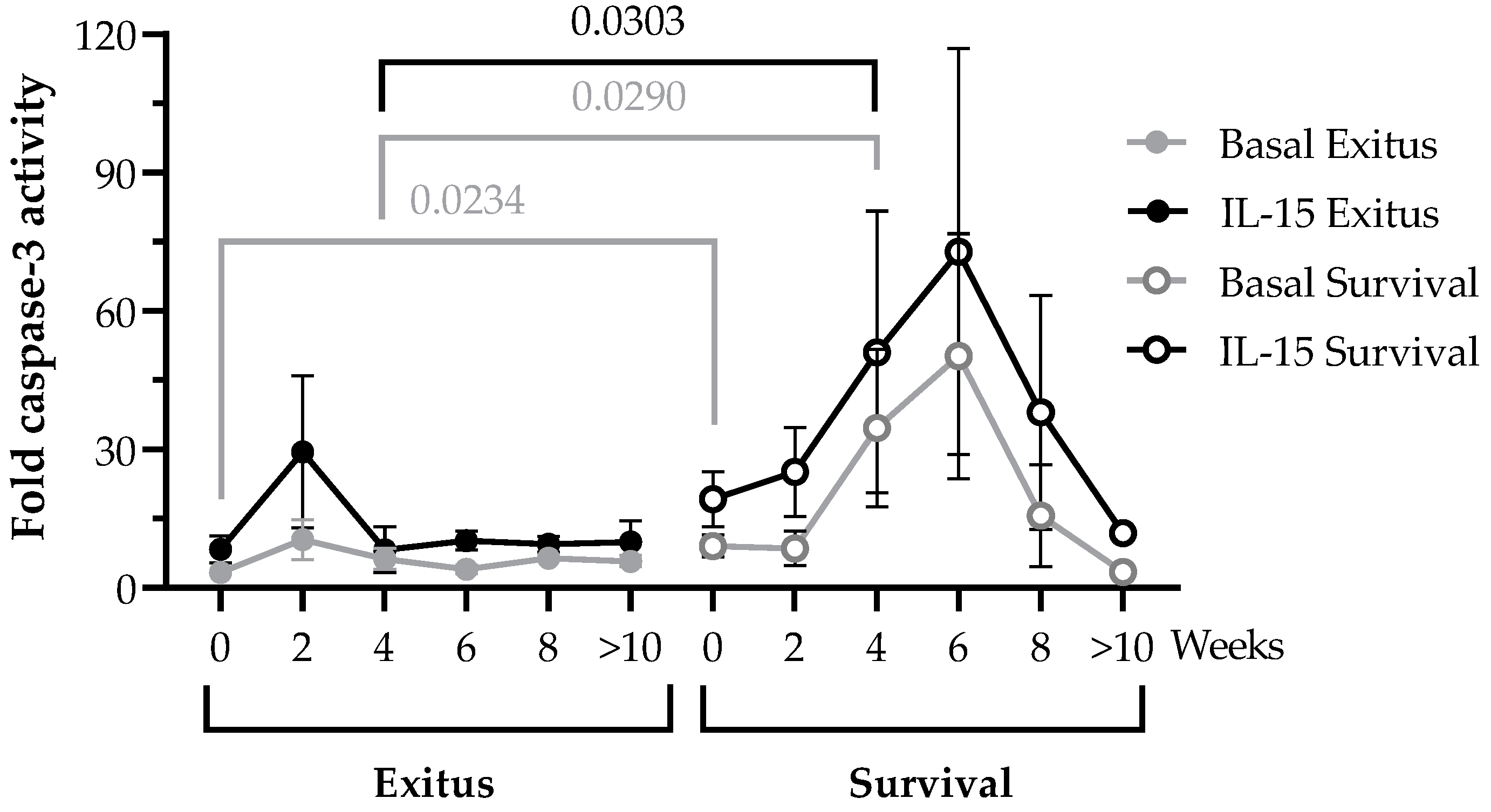
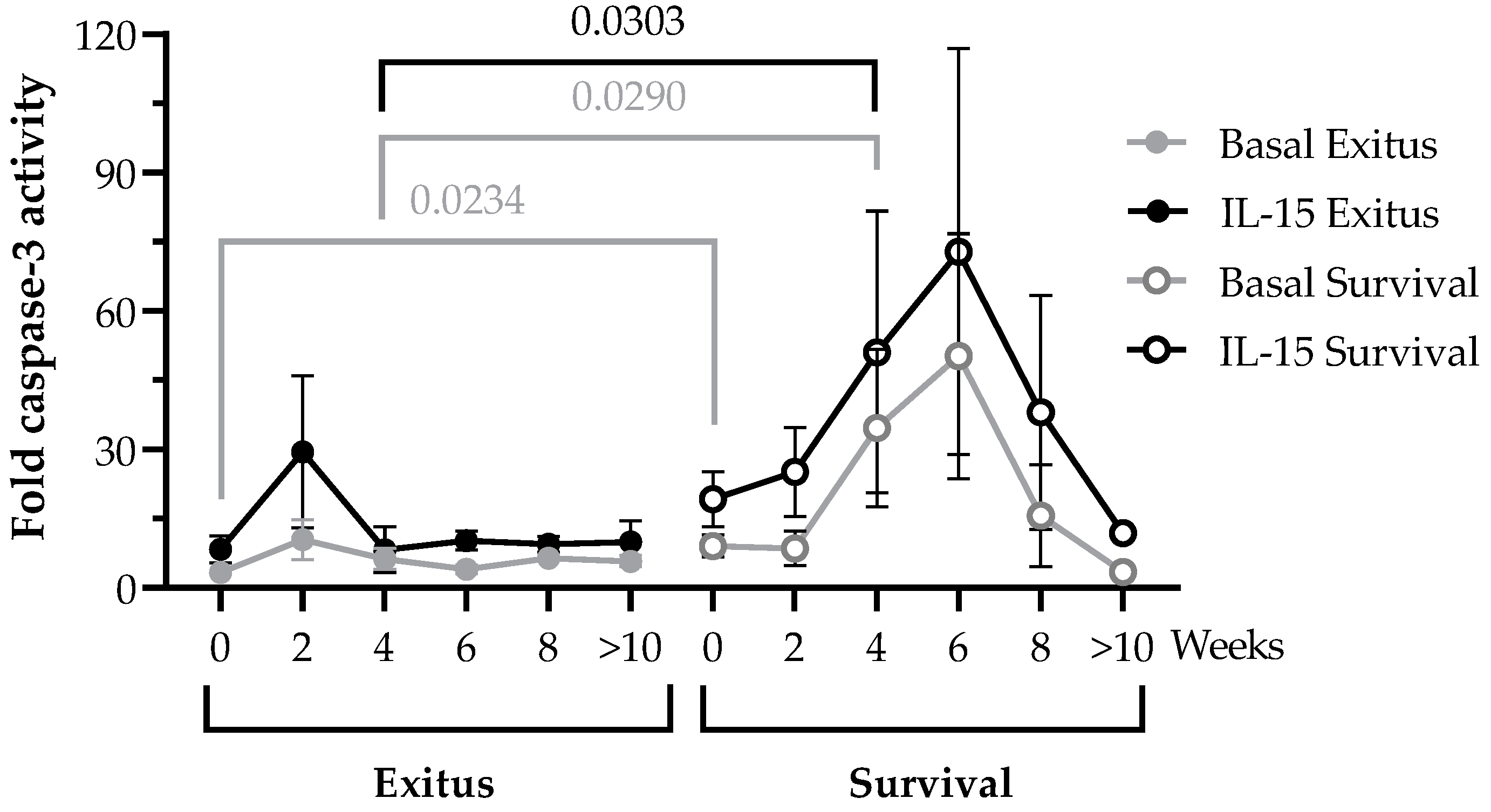
3.4. PBMCs Cytotoxic Activity against Vero E6 Cells Infected with Pseudotyped SARS-CoV-2
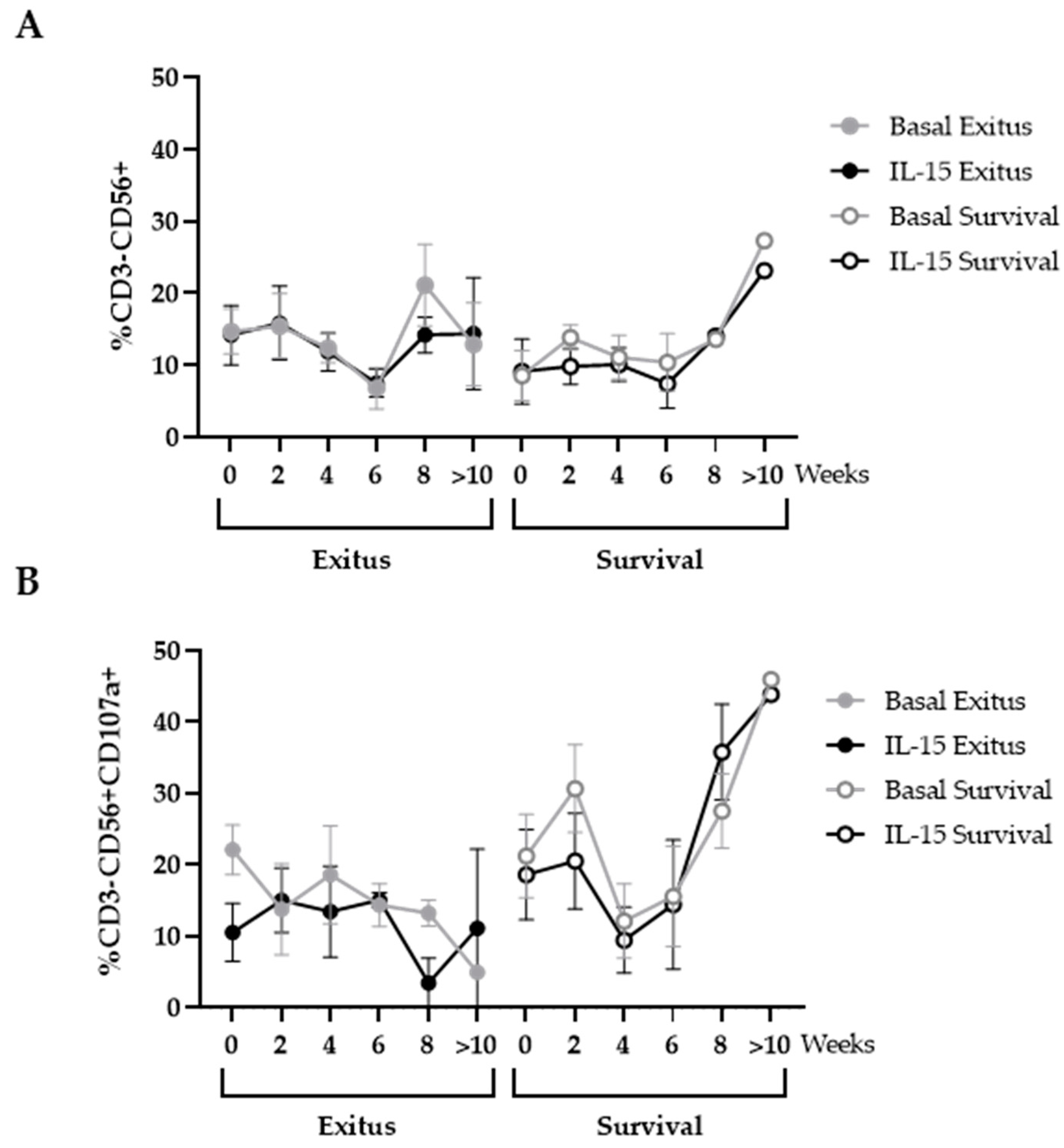
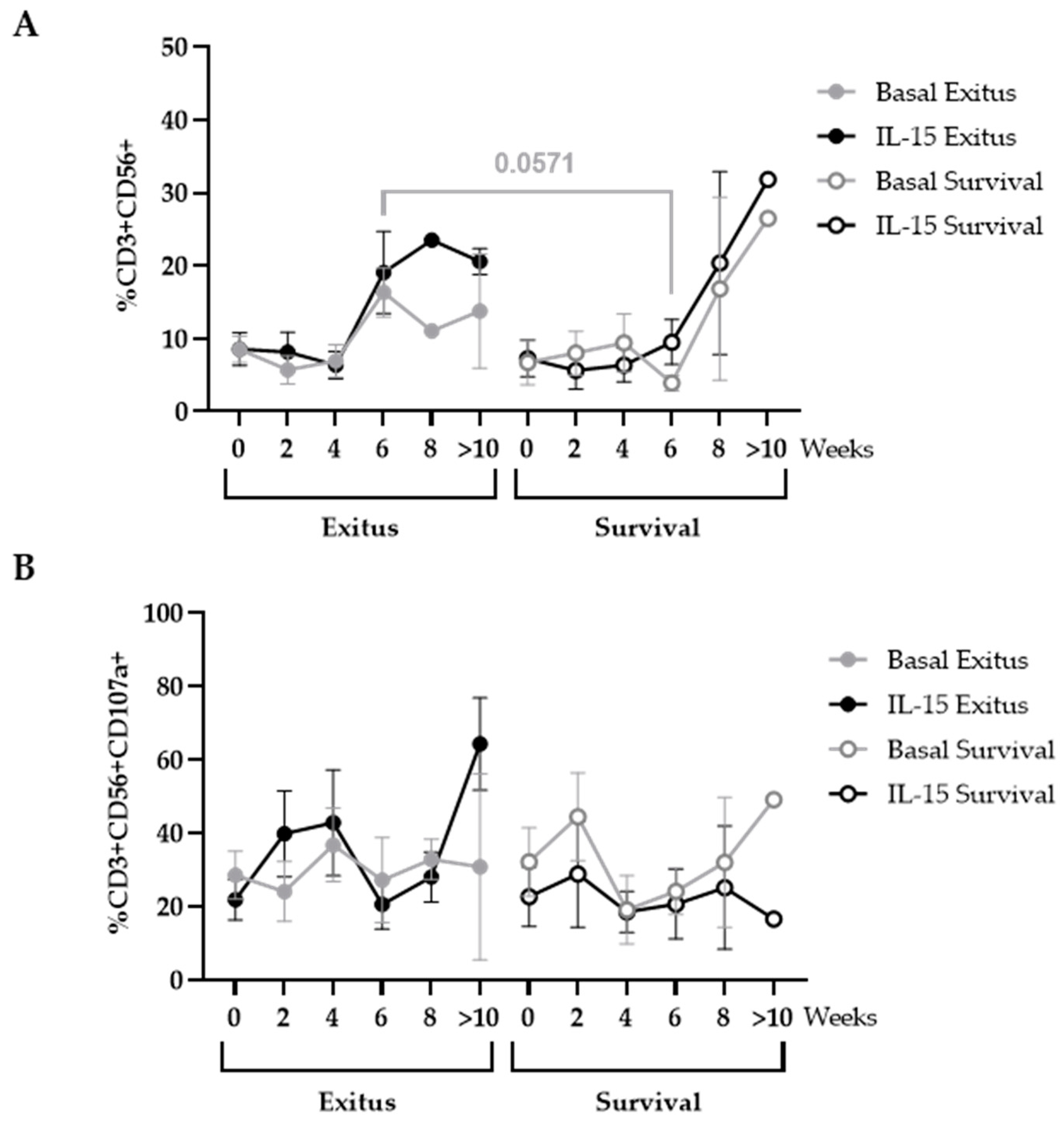
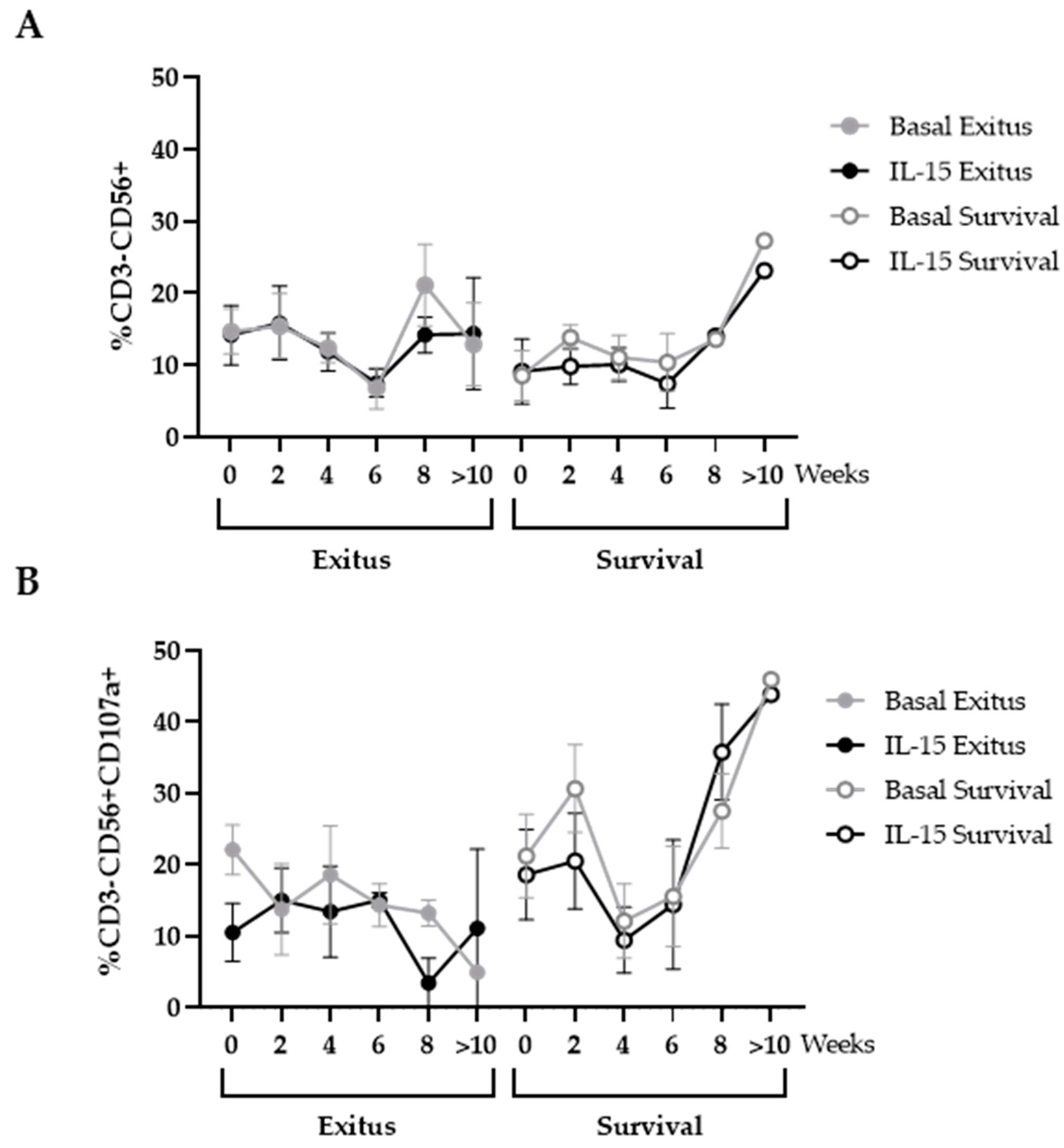
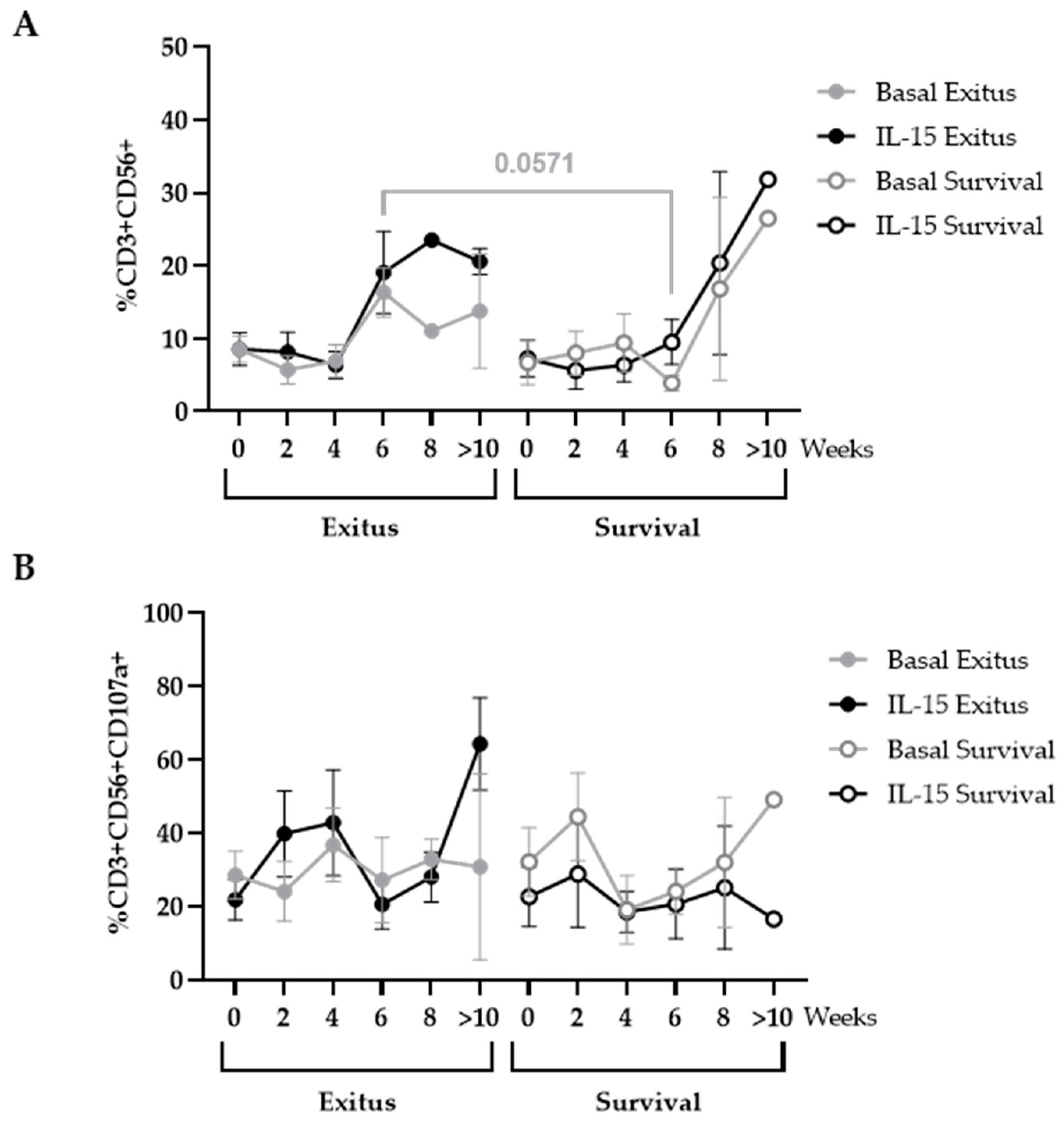
3.5. Levels of NK and NKT Cells
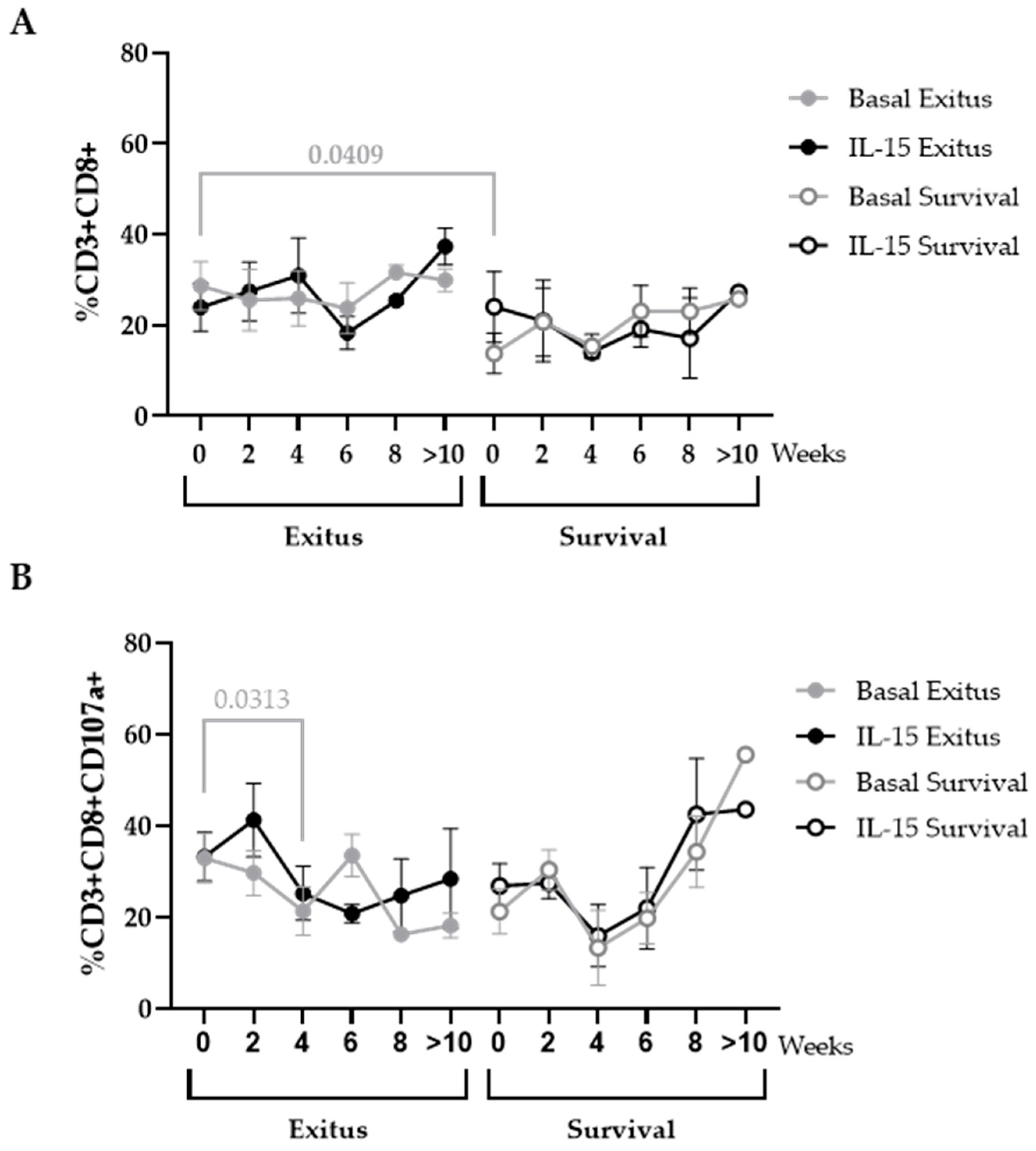
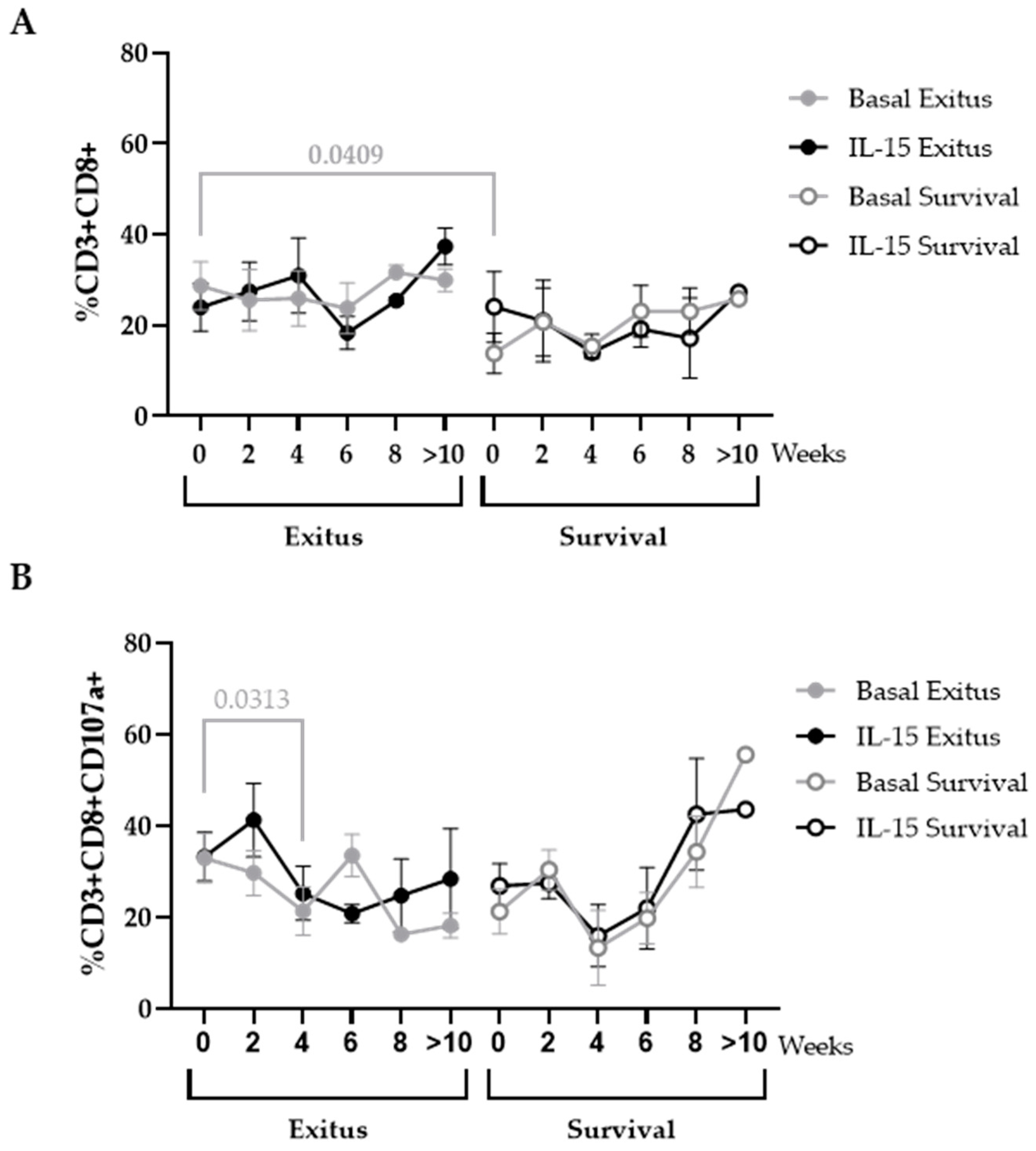
3.6. Levels of CD8+ T Cells
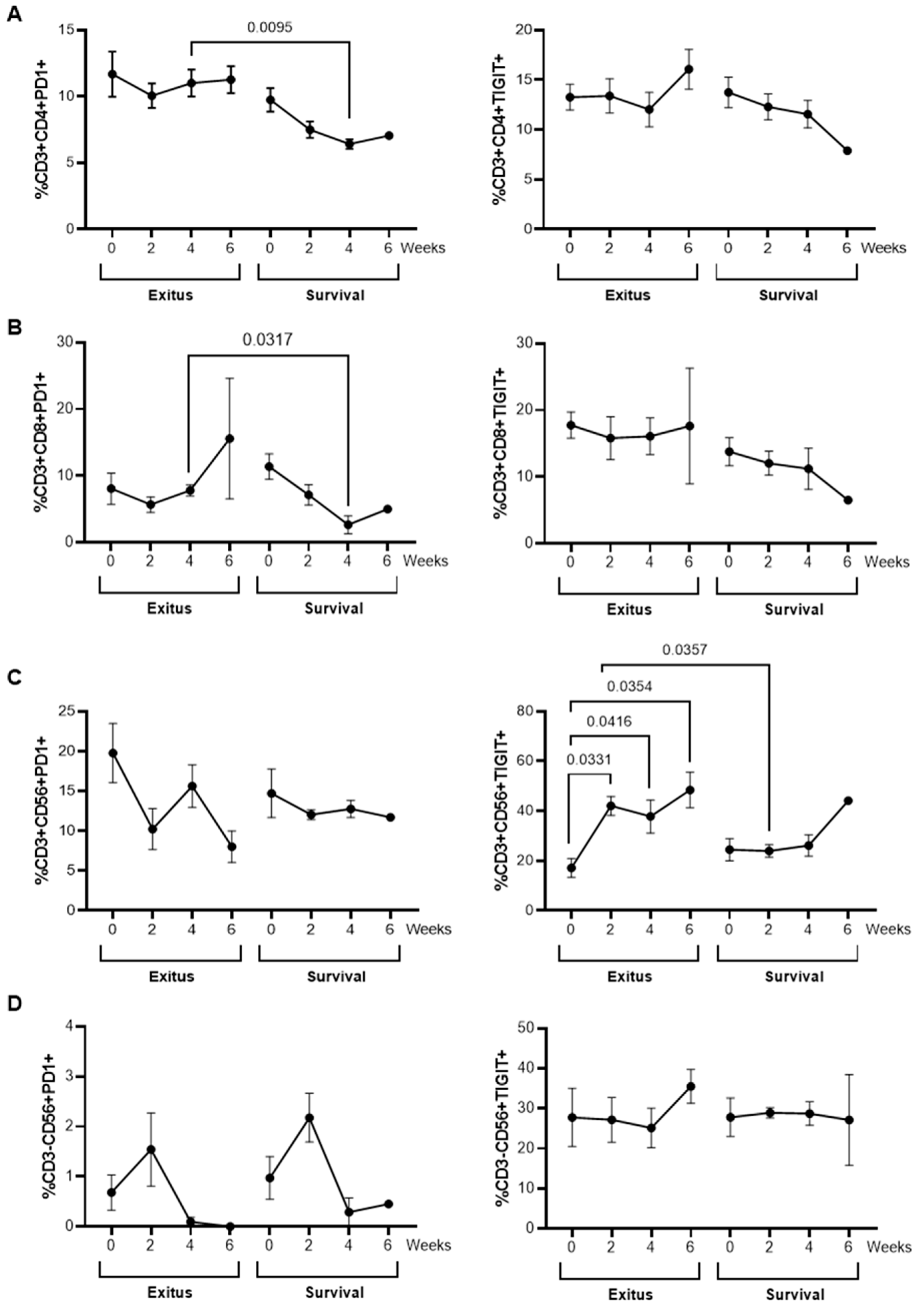
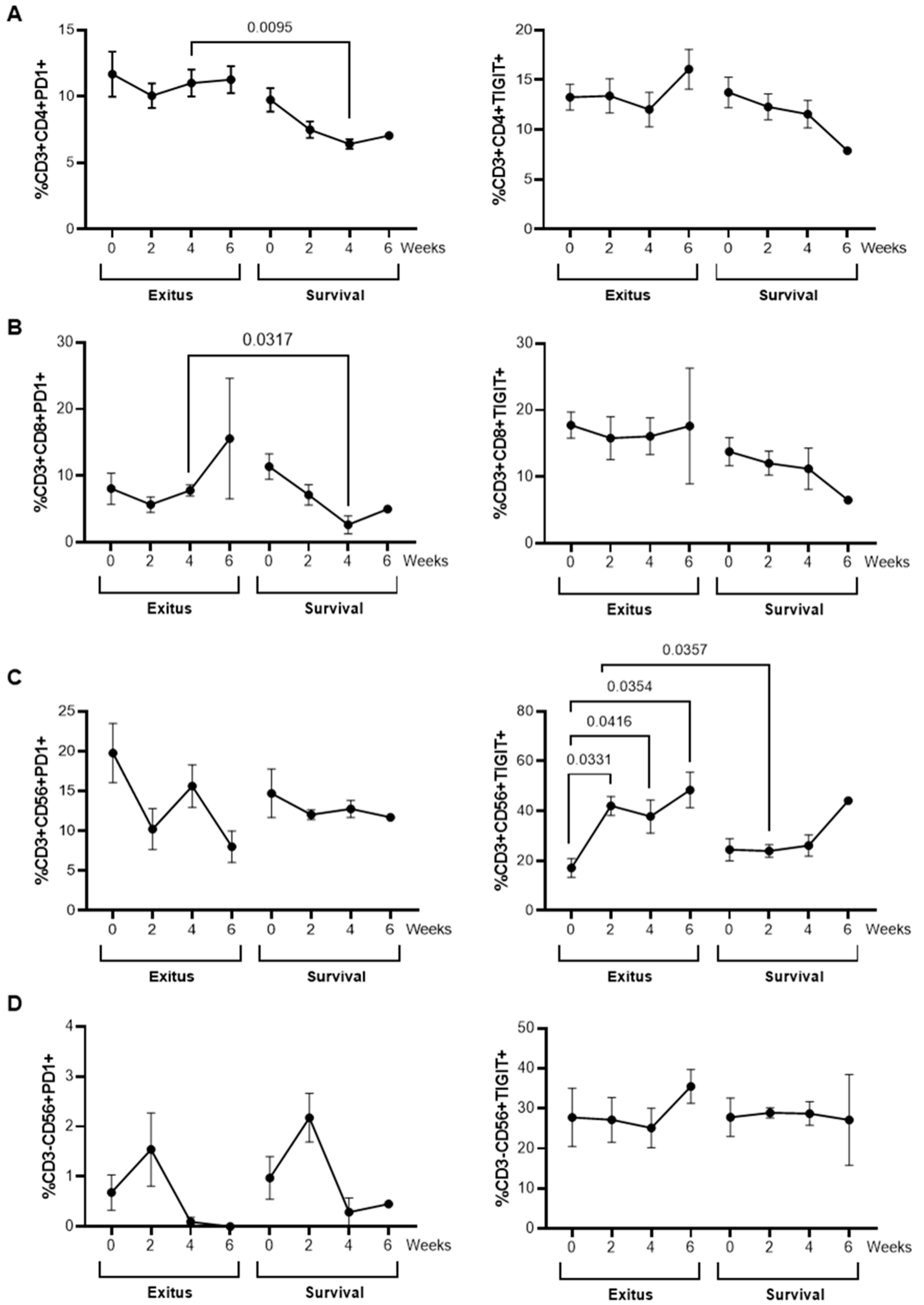
3.7. Levels of Exhaustion Markers
3.8. Quantification of SARS-CoV-2 RNAemia in Plasma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atzrodt, C.L.; Maknojia, I.; McCarthy, R.D.P.; Oldfield, T.M.; Po, J.; Ta, K.T.L.; Stepp, H.E.; Clements, T.P. A Guide to COVID-19: A global pandemic caused by the novel coronavirus SARS-CoV-2. FEBS J. 2020, 287, 3633–3650. [Google Scholar] [CrossRef] [PubMed]
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 18 October 2022).
- Capalbo, C.; Aceti, A.; Simmaco, M.; Bonfini, R.; Rocco, M.; Ricci, A.; Napoli, C.; Rocco, M.; Alfonsi, V.; Teggi, A.; et al. The exponential phase of the COVID-19 pandemic in central Italy: An integrated care pathway. Int. J. Environ. Res. 2020, 17, 3792. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haybar, H.; Kazemnia, K.; Rahim, F. Underlying chronic disease and COVID-19 infection: A state-of-the-art review. Jundishapur J. Chronic Dis. Care 2020, 9, e103452. [Google Scholar] [CrossRef]
- Hamed, S.M.; Elkhatib, W.F.; Khairalla, A.S.; Noreddin, A.M. Global dynamics of SARS-CoV-2 clades and their relation to COVID-19 epidemiology. Sci. Rep. 2021, 11, 8435. [Google Scholar] [CrossRef]
- Erdinc, B.; Sahni, S.; Gotlieb, V. Hematological manifestations and complications of COVID-19. Adv. Clin. Exp. Med. 2021, 30, 101–107. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The cytokine storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef]
- Ramos-Casals, M.; Brito-Zerón, P.; López-Guillermo, A.; Khamashta, M.A.; Bosch, X. Adult haemophagocytic syndrome. Lancet 2014, 383, 1503–1516. [Google Scholar] [CrossRef]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J.; on behalf of the HLH Across Speciality Collaboration, UK. Across Speciality. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Li, M.-Y.; Li, L.; Zhang, Y.; Wang, X.-S. Expression of the SARS-CoV-2 cell receptor gene ACE2 in a wide variety of human tissues. Infect. Dis. Poverty 2020, 9, 45. [Google Scholar] [CrossRef]
- Jiang, Y.; Rubin, L.; Peng, T.; Liu, L.; Xing, X.; Lazarovici, P.; Zheng, W. Cytokine storm in COVID-19: From viral infection to immune responses, diagnosis and therapy. Int. J. Biol. Sci. 2022, 18, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Carrington, E.M.; Zhang, Y.; Heinzel, S.; Lew, A.M. Life and death of activated T cells: How are they different from naïve T cells? Front. Immunol. 2017, 8, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, I.; Pranata, R. Lymphopenia in severe coronavirus disease-2019 (COVID-19): Systematic review and meta-analysis. J. Intensive Care 2020, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Gao, Y.; Wang, G.; Song, G.; Liu, S.; Sun, D.; Xu, Y.; Tian, Z. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 533–535. [Google Scholar] [CrossRef] [Green Version]
- Rha, M.-S.; Jeong, H.W.; Ko, J.-H.; Choi, S.J.; Seo, I.-H.; Lee, J.S.; Sa, M.; Kim, A.R.; Joo, E.-J.; Ahn, J.Y.; et al. PD-1-expressing SARS-CoV-2-specific CD8+ T cells are not exhausted, but functional in patients with COVID-19. Immunity 2021, 54, 44–52. [Google Scholar] [CrossRef]
- Vigón, L.; Fuertes, D.; García-Pérez, J.; Torres, M.; Rodríguez-Mora, S.; Mateos, E.; Corona, M.; Saez-Marín, A.J.; Malo, R.; Navarro, C.; et al. Impaired Cytotoxic Response in PBMCs from Patients with COVID-19 Admitted to the ICU: Biomarkers to Predict Disease Severity. Front. Immunol. 2021, 12, 665329. [Google Scholar] [CrossRef]
- WHO. Interim Statement on Decision-Making Considerations for the Use of Variant Updated COVID-19 Vaccines. Available online: https://www.who.int/news/item/17-06-2022-interim-statement-on-decision-making-considerations-for-the-use-of-variant-updated-covid-19-vaccines (accessed on 15 August 2022).
- Mishra, S.K.; Tripathi, T. One year update on the COVID-19 pandemic: Where are we now? Acta Trop. 2021, 214, 105778. [Google Scholar] [CrossRef]
- Li, X.; Geng, M.; Peng, Y.; Meng, L.; Lu, S. Molecular immune pathogenesis and diagnosis of COVID-19. J. Pharm. Anal. 2020, 10, 102–108. [Google Scholar] [CrossRef]
- TRC Group. Dexamethasone in hospitalized patients with COVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- Zhou, X.; Yu, J.; Cheng, X.; Zhao, B.; Manyam, G.C.; Zhang, L.; Schluns, K.; Li, P.; Wang, J.; Sun, S.-C. The deubiquitinase Otub1 controls the activation of CD8+ T cells and NK cells by regulating IL-15-mediated priming. Nat. Immunol. 2019, 20, 879–889. [Google Scholar] [CrossRef]
- Parums, D.V. Current status of oral antiviral drug treatments for SARS-CoV-2 infection in non-hospitalized patients. Med. Sci. Monit. 2022, 28, e935952-1. [Google Scholar] [CrossRef] [PubMed]
- Couzin-Frankel, J. Antiviral pills could change pandemic’s course. Am. Assoc. Adv. Sci. 2021, 12, 799–800. [Google Scholar] [CrossRef] [PubMed]
- Muntasell, A.; Ochoa, M.C.; Cordeiro, L.; Berraondo, P.; de Cerio, A.L.-D.; Cabo, M.; López-Botet, M.; Melero, I. Targeting NK-cell checkpoints for cancer immunotherapy. Curr. Opin. Immunol. 2017, 45, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Patidar, M.; Yadav, N.; Dalai, S.K. Interleukin 15: A key cytokine for immunotherapy. Cytokine Growth Factor Rev. 2016, 31, 49–59. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of molnupiravir-induced SARS-CoV-2 mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.-P.; Lee, J.-C.; Chiu, C.-W.; Lee, C.-C.; Tsai, P.-J.; Hsu, I.-L.; Ko, W.-C. Oral Nirmatrelvir/Ritonavir Therapy for COVID-19: The Dawn in the Dark? Antibiotics 2022, 11, 220. [Google Scholar] [CrossRef]
- Guo, Y.; Luan, L.; Patil, N.K.; Sherwood, E.R. Immunobiology of the IL-15/IL-15Rα complex as an antitumor and antiviral agent. Cytokine Growth Factor Rev. 2017, 38, 10–21. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, J.; Bai, X.; Handley, M.; Shan, F. Biological effects of IL-15 on immune cells and its potential for the treatment of cancer. Int. Immunopharmacol. 2021, 91, 107318. [Google Scholar] [CrossRef]
- Waldmann, T.A.; Dubois, S.; Miljkovic, M.D.; Conlon, K.C. IL-15 in the Combination Immunotherapy of Cancer. Front. Immunol. 2020, 11, 868. [Google Scholar] [CrossRef]
- Dubois, S.P.; Miljkovic, M.D.; Fleisher, T.A.; Pittaluga, S.; Hsu-Albert, J.; Bryant, B.R.; Petrus, M.N.; Perera, L.P.; Müller, J.R.; Shih, J.H.; et al. Short-course IL-15 given as a continuous infusion led to a massive expansion of effective NK cells: Implications for combination therapy with antitumor antibodies. J. Immunother. Cancer 2021, 9, e002193. [Google Scholar] [CrossRef]
- Conlon, K.C.; Potter, E.L.; Pittaluga, S.; Lee, C.-C.R.; Miljkovic, M.D.; Fleisher, T.A.; Dubois, S.; Bryant, B.R.; Petrus, M.; Perera, L.P.; et al. IL15 by Continuous Intravenous Infusion to Adult Patients with Solid Tumors in a Phase I Trial Induced Dramatic NK-Cell Subset. Clin. Cancer Res. 2019, 25, 4945–4954. [Google Scholar] [CrossRef]
- Romee, R.; Cooley, S.; Berrien-Elliott, M.M.; Westervelt, P.; Verneris, M.R.; Wagner, J.E.; Weisdorf, D.J.; Blazar, B.R.; Ustun, C.; DeFor, T.E.; et al. First-in-human phase 1 clinical study of the IL-15 superagonist complex ALT-803 to treat relapse after transplantation. Blood Am. J. Hematol. 2018, 131, 2515–2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.S.; Davis, Z.B.; Helgeson, E.; Reilly, C.; Thorkelson, A.; Anderson, J.; Lima, N.S.; Jorstad, S.; Hart, G.T.; Lee, J.H.; et al. Safety and virologic Impact of the IL-15 superagonist N-803 in people living with HIV: A phase 1 trial. Nat. Med. 2022, 28, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Berenguer, J.; Ryan, P.; Rodríguez-Baño, J.; Jarrín, I.; Carratalà, J.; Pachón, J.; Yllescas, M.; Arriba, J.R.; Muñoz, E.A.; Gil Divasson, P.; et al. Characteristics and predictors of death among 4035 consecutively hospitalized patients with COVID-19 in Spain. Clin. Microbiol. Infect. 2020, 26, 1525–1536. [Google Scholar] [CrossRef]
- Garcia-Perez, J.; Sanchez-Palomino, S.; Perez-Olmeda, M.; Fernandez, B.; Alcami, J. A new strategy based on recombinant viruses as a tool for assessing drug susceptibility of human immunodeficiency virus type 1. J. Med. Virol. 2007, 79, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volz, E.; Hill, V.; McCrone, J.T.; Price, A.; Jorgensen, D.; O’Toole, Á.; Southgate, J.; Johnson, R.; Jackson, B.; Nascimento, F.F.; et al. Evaluating the effects of SARS-CoV-2 spike mutation D614G on transmissibility and pathogenicity. Cell 2021, 184, 64–75. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking changes in SARS-CoV-2 spike: Evidence that D614G increases infectivity of the COVID-19 virus. Cell 2020, 182, 812–827. [Google Scholar] [CrossRef]
- Nandagopal, N.; Ali, A.K.; Komal, A.K.; Lee, S.-H. The critical role of IL-15–PI3K–mTOR pathway in natural killer cell effector functions. Front. Immunol. 2014, 5, 187. [Google Scholar] [CrossRef]
- Rosenthal, R.; Groeper, C.; Bracci, L.; Adamina, M.; Feder-Mengus, C.; Zajac, P.; Iezzi, G.; Bolli, M.; Weber, W.P.; Frey, D.M.; et al. Differential Responsiveness to IL-2, IL-7, and IL-15 Common Receptor γ Chain Cytokines by Antigen-specific Peripheral Blood Naive or Memory Cytotoxic CD8+ T Cells from Healthy Donors and Melanoma Patients. J. Immunother. 2009, 32, 252–261. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Diagnostic Testing for SARS-CoV-2: Interim Guidance, 11 September 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Becker, L.A. Effect Size (ES); University of Colorado Colorado Springs: Colorado Springs, CO, USA, 2000. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge: Abingdon, UK, 1988. [Google Scholar] [CrossRef]
- Moreno-Perez, O.; Ribes, I.; Boix, V.; Martinez-García, M.; Otero-Rodriguez, S.; Reus, S.; Sánchez-Martínez, R.; Ramos, J.M.; Chico-Sánchez, P.; Merino, E. Hospitalized patients with breakthrough COVID-19: Clinical features and poor outcome predictors. Int. J. Infect. Dis. 2022, 118, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Lee, E.; Park, W.B.; Choe, P.G.; Song, K.-H.; Kim, E.S.; Park, S.-W. Breakthrough COVID-19 Infection during the Delta Variant Dominant Period: Individualized Care Based on Vaccination Status Is Needed. J. Korean Med. Sci. 2022, 37, e252. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, H.; Alsrhani, A.; Zafar, A.; Javed, H.; Junaid, K.; Abdalla, A.E.; Abosalif, K.O.; Ahmed, Z.; Younas, S. COVID-19 and comorbidities: Deleterious impact on infected patients. J. Infect. Public Health 2020, 13, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, R.B.; Botelho, B.G.; de Hollanda, J.V.G.; Ferreira, L.V.L.; de Andrade, L.Z.J.; Oei, S.S.M.L.; Mello, T.D.S.; Muxfeldt, E.S. COVID-19 and the cardiovascular system: A comprehensive review. J. Hum. Hypertens. 2021, 35, 4–11. [Google Scholar] [CrossRef]
- Zhang, J.J.; Lee, K.S.; Ang, L.W.; Leo, Y.S.; Young, B.E. Risk factors for severe disease and efficacy of treatment in patients infected with COVID-19: A systematic review, meta-analysis, and meta-regression analysis. Clin. Infect. Dis. 2020, 71, 2199–2206. [Google Scholar] [CrossRef]
- Kermali, M.; Khalsa, R.K.; Pillai, K.; Ismail, Z.; Harky, A. The role of biomarkers in diagnosis of COVID-19–A systematic review. Life Sci. 2020, 254, 117788. [Google Scholar] [CrossRef]
- Malik, P.; Patel, U.; Mehta, D.; Patel, N.; Kelkar, R.; Akrmah, M.; Gabrilove, J.L.; Sacks, H. Biomarkers and outcomes of COVID-19 hospitalisations: Systematic review and meta-analysis. BMJ Evid. Based Med. 2021, 26, 107–108. [Google Scholar] [CrossRef]
- Li, Y.; Schneider, A.M.; Mehta, A.; Sade-Feldman, M.; Kays, K.R.; Gentili, M.; Charland, N.C.; Gonye, A.L.; Gushterova, I.; Khanna, H.K.; et al. SARS-CoV-2 viremia is associated with distinct proteomic pathways and predicts COVID-19 outcomes. J. Clin. Investig. 2021, 131, e148635. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, D.A.; Roy-Vallejo, E.; Cruz, N.D.Z.; Ramírez, A.M.; Rodríguez-García, S.C.; Arevalillo-Fernández, N.; Galván-Román, J.M.; García-Rodrigo, L.F.; Vega-Piris, L.; Llano, M.C.; et al. Detection of SARS-CoV-2 RNA in serum is associated with increased mortality risk in hospitalized COVID-19 patients. Sci. Rep. 2021, 11, 13134. [Google Scholar] [CrossRef]
- Serafim, R.B.; Póvoa, P.; Souza-Dantas, V.; Kalil, A.C.; Salluh, J.I. Clinical course and outcomes of critically ill patients with COVID-19 infection: A systematic review. Clin. Microbiol. Infect. 2021, 27, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, H.; Zhou, Y.; Wu, X.; Zhao, Y.; Lu, Y.; Tan, W.; Yuan, M.; Ding, X.; Zou, J.; et al. Risk factors associated with disease severity and length of hospital stay in COVID-19 patients. J. Infect. 2020, 81, e95–e97. [Google Scholar] [CrossRef]
- Luo, X.H.; Zhu, Y.; Mao, J.; Du, R.C. T cell immunobiology and cytokine storm of COVID-19. Scand. J. Immunol. 2021, 93, e12989. [Google Scholar] [CrossRef] [PubMed]
- Harwood, O.; O’Connor, S. Therapeutic potential of IL-15 and N-803 in HIV/SIV infection. Viruses 2021, 13, 1750. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Wherry, E.J. CD8 T cell dysfunction during chronic viral infection. Curr. Opin. Immunol. 2007, 19, 408–415. [Google Scholar] [CrossRef]
- McMahan, K.; Yu, J.; Mercado, N.B.; Loos, C.; Tostanoski, L.H.; Chandrashekar, A.; Liu, J.; Peter, L.; Atyeo, C.; Zhu, A.; et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 2021, 590, 630–634. [Google Scholar] [CrossRef]
- Zhuang, Z.; Lai, X.; Sun, J.; Chen, Z.; Zhang, Z.; Dai, J.; Liu, D.; Li, Y.; Li, F.; Wang, Y.; et al. Mapping and role of T cell response in SARS-CoV-2–infected mice. J. Exp. Med. 2021, 218, e20202187. [Google Scholar] [CrossRef]
- Westmeier, J.; Paniskaki, K.; Karaköse, Z.; Werner, T.; Sutter, K.; Dolff, S.; Overbeck, M.; Limmer, A.; Liu, J.; Zheng, X.; et al. Impaired cytotoxic CD8+ T cell response in elderly COVID-19 patients. MBio 2020, 11, e02243-20. [Google Scholar] [CrossRef]
- Laing, A.G.; Lorenc, A.; del Barrio, I.D.M.; Das, A.; Fish, M.; Monin, L.; Muñoz-Ruiz, M.; McKenzie, D.R.; Hayday, T.S.; Francos-Quijorna, I.; et al. A dynamic COVID-19 immune signature includes associations with poor prognosis. Nat. Med. 2020, 26, 1623–1635. [Google Scholar] [CrossRef]
- Kuri-Cervantes, L.; Pampena, M.B.; Meng, W.; Rosenfeld, A.M.; Ittner, C.A.G.; Weisman, A.R.; Agyekum, R.S.; Mathew, D.; Baxter, A.E.; Vella, L.A.; et al. Comprehensive mapping of immune perturbations associated with severe COVID-19. Sci. Immunol. 2020, 5, eabd7114. [Google Scholar] [CrossRef]
- Aktas, E.; Kucuksezer, U.C.; Bilgic, S.; Erten, G.; Deniz, G. Relationship between CD107a expression and cytotoxic activity. Cell. Immunol. 2009, 254, 149–154. [Google Scholar] [CrossRef]
- Björkström, N.K.; Strunz, B.; Ljunggren, H.-G. Natural killer cells in antiviral immunity. Nat. Rev. Immunol. 2022, 22, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Bi, J. NK cell dysfunction in patients with COVID-19. Cell. Mol. Immunol. 2022, 19, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Gustine, J.N.; Jones, D. Immunopathology of Hyperinflammation in COVID-19. Am. J. Clin. Pathol. 2021, 191, 4–17. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zhou, Y.; Li, Z.; Farooq, M.A.; Ajmal, I.; Zhang, H.; Zhang, L.; Tao, L.; Yao, J.; Du, B.; et al. Co-expression of IL-7 improves NKG2D-based CAR T cell therapy on prostate cancer by enhancing the expansion and inhibiting the apoptosis and exhaustion. Cancers 2020, 12, 1969. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exitus (n = 13) | Survival (n = 10) | |
|---|---|---|
| Demographic characteristics | ||
| Age, median (IQR) | 65.0 (62.0–69.0) | 63.0 (59.0–68.5) |
| Sex: male, n (%) | 11 (84.6) | 6 (60.0) |
| Hospital stay | ||
| Hospitalization days, median (IQR) | 59.0 (36.0–100.0) | 73.0 (44.5–90.0) |
| Days in ICU (IQR) | 49.0 (25.5–82.0) | 44.0 (25.0–66.5) |
| Comorbidities | ||
| One or more comorbidity, n (%) | 9 (69.23) | 6 (60.0) |
| DM, n (%) | 4 (30.8) | 2 (20.0) |
| DL, n (%) | 6 (54.5) | 6 (60.0) |
| HT, n (%) | 5 (41.7) | 4 (40.0) |
| Treatments | ||
| Corticoids, n (%) | 10 (76.9) | 9 (90.0) |
| Antibiotics, n (%) | 10 (76.9) | 9 (90.0) |
| Anticoagulants, n (%) | 10 (76.9) | 9 (90.0) |
| Tocilizumab, n (%) | 3 (23.1) | 1 (10.0) |
| Remdesivir, n (%) | 1 (7.7) | 1 (10.0) |
| Signs and symptoms | ||
| Pneumonia, n (%) | 13 (100) | 10 (100) |
| Cough and expectoration, n (%) | 11 (84.6) | 9 (90.0) |
| Dyspnea, n (%) | 11 (84.6) | 9 (90.0) |
| Fever, n (%) | 12 (92.3) | 8 (80.0) |
| Blood biochemistry data at baseline | ||
| CRP (mg/L) (IQR) | 108.0 (30.2–202.0) | 123.4 (58.1–172.6) |
| LDH (U/L) (IQR) | 423.5 (319.8–535.8) | 388.0 (365.0–470.0) |
| Lymphocytes (cells/μL) (IQR) | 860.0 (620.0–1090.0) | 1040.0 (930.0–1500.0) |
| Monocytes (cells/μL) (IQR) | 490.0 (290.0–700.0) | 670.0 (462.5–770.0) |
| Platelets (103 cells/μL) (IQR) | 191.0 (142.0–277.0) | 284.0 (182.5–450.7) |
| Fibrinogen (mg/dL) (IQR) | 740.0 (608.8–740.0) | 740.0 (601.0–740.0) |
| Ventilation | ||
| NIV, n (%) | 1 (7.7) | 0 (0) |
| IV, n (%) | 12 (92.3) | 10 (100.0) |
| Reservoir, n (%) | 1 (7.7) | 2 (20.0) |
| Nasal glasses, n (%) | 1 (7.7) | 5 (50.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casado-Fernández, G.; Corona, M.; Torres, M.; Saez, A.J.; Ramos-Martín, F.; Manzanares, M.; Vigón, L.; Mateos, E.; Pozo, F.; Casas, I.; et al. Sustained Cytotoxic Response of Peripheral Blood Mononuclear Cells from Unvaccinated Individuals Admitted to the ICU Due to Critical COVID-19 Is Essential to Avoid a Fatal Outcome. Int. J. Environ. Res. Public Health 2023, 20, 1947. https://doi.org/10.3390/ijerph20031947
Casado-Fernández G, Corona M, Torres M, Saez AJ, Ramos-Martín F, Manzanares M, Vigón L, Mateos E, Pozo F, Casas I, et al. Sustained Cytotoxic Response of Peripheral Blood Mononuclear Cells from Unvaccinated Individuals Admitted to the ICU Due to Critical COVID-19 Is Essential to Avoid a Fatal Outcome. International Journal of Environmental Research and Public Health. 2023; 20(3):1947. https://doi.org/10.3390/ijerph20031947
Chicago/Turabian StyleCasado-Fernández, Guiomar, Magdalena Corona, Montserrat Torres, Adolfo J. Saez, Fernando Ramos-Martín, Mario Manzanares, Lorena Vigón, Elena Mateos, Francisco Pozo, Inmaculada Casas, and et al. 2023. "Sustained Cytotoxic Response of Peripheral Blood Mononuclear Cells from Unvaccinated Individuals Admitted to the ICU Due to Critical COVID-19 Is Essential to Avoid a Fatal Outcome" International Journal of Environmental Research and Public Health 20, no. 3: 1947. https://doi.org/10.3390/ijerph20031947
APA StyleCasado-Fernández, G., Corona, M., Torres, M., Saez, A. J., Ramos-Martín, F., Manzanares, M., Vigón, L., Mateos, E., Pozo, F., Casas, I., García-Gutierrez, V., Rodríguez-Mora, S., & Coiras, M. (2023). Sustained Cytotoxic Response of Peripheral Blood Mononuclear Cells from Unvaccinated Individuals Admitted to the ICU Due to Critical COVID-19 Is Essential to Avoid a Fatal Outcome. International Journal of Environmental Research and Public Health, 20(3), 1947. https://doi.org/10.3390/ijerph20031947