
Artisanal Gem Mining in Brazil: Evaluation of Oxidative Stress and Genotoxicity Biomarkers

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Samples
2.2. Oxidative Stress Biomarkers
2.2.1. Total Peroxides in Plasma
2.2.2. Determination of the Antioxidant Enzyme Catalase (CAT) Activity
2.3. Environmental Assessment through Genotoxicity and Mutagenicity Studies in Soil Samples
2.3.1. Allium cepa Test
2.3.2. Ames Test
2.4. Statistical Analysis
3. Results and Discussion
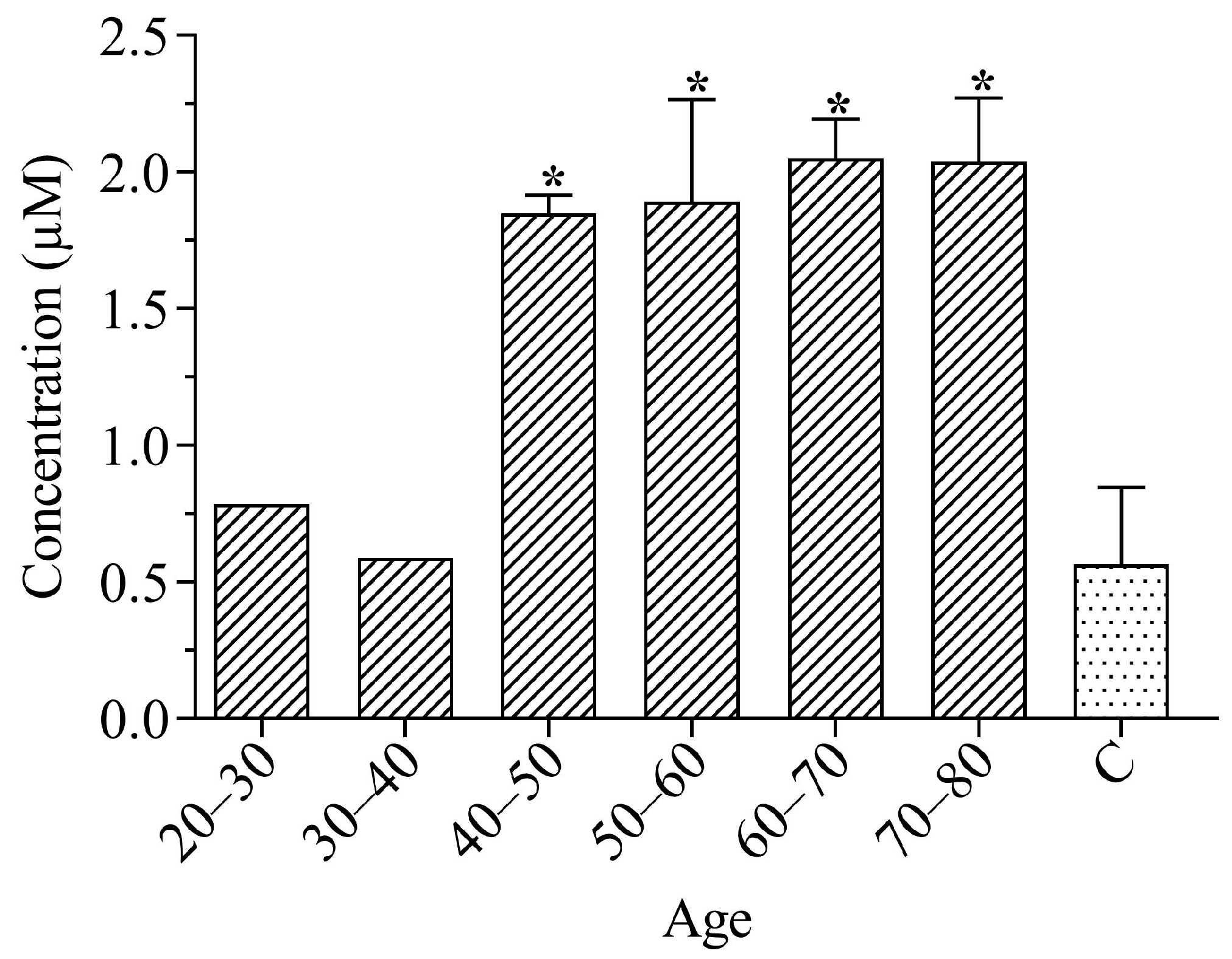
3.1. Total Peroxides in Plasma
3.2. Total Catalase Activity
3.3. Micronutrients
3.4. Evaluation of Cytotoxicity, Genotoxicity and Mutagenicity by the Allium cepa Bioassay
3.4.1. Cytotoxicity Analysis
3.4.2. Analysis of Genotoxicity and Mutagenicity
3.5. Ames Test
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peruzzi, C.; Nascimento, S.; Gauer, B.; Nardi, J.; Sauer, E.; Göethel, G.; Garcia, S.C. Inflammatory and oxidative stress biomarkers at protein and molecular levels in workers occupationally exposed to crystalline silica. Environ. Sci. Pollut. Res. 2019, 26, 1394–1405. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.P.R.; Pêgo, J.S. Riscos Associados aos Garimpos de Gemas no Povoado de Taquaral, Itinga, MG: Dois Contextos Dilemáticos; IFNMG: Araçuaí, Brazil, 2014. [Google Scholar]
- Santos, A.P.R.; Pego, J.S.; Muniz, E.O. Contextualização dos impactos ambientais em áreas de extração de gemas do Povoado de Taquaral, Itinga (MG). Rev. Agrogeoambiental 2016, 8, 61–69. [Google Scholar] [CrossRef]
- Stewart, A.G. Mining is bad for health: A voyage of discovery. Environ. Geochem. Health 2020, 42, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Amorim, L.C.A. Biomarkers for evaluating exposure to chemical agents present in the environment. Rev. Bras. De Epidemiol. 2003, 6, 158–170. [Google Scholar] [CrossRef]
- Bonciu, E.; Firbas, P.; Fontanetti, C.S.; Wusheng, J.; Karaismailoğlu, M.C.; Liu, D.; Menicucci, F.; Pesnya, D.S.; Popescu, I.A.; Romanovsky, A.V.; et al. An evaluation for the standardization of the Allium cepa test as cytotoxicity and genotoxicity assay. Caryologia 2018, 71, 191–209. [Google Scholar] [CrossRef]
- Tagliari, K.C.; Cecchini, R.; Saridakis, H.O. Teste de ames como uma ferramenta para detecção de citotoxidade e mutagenicidade causadas por metais pesados e radicais livres. Semin. Ciências Biológicas E Da Saúde 1999, 18, 41–50. [Google Scholar] [CrossRef]
- Demirci-Cekic, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of oxidative stress and antioxidant defense. J. Pharm. Biomed. Anal. 2022, 209, 114477. [Google Scholar] [CrossRef] [PubMed]
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of oxidative stress in metabolic syndrome and associated diseases. Oxidative Med. Cell. Longev. 2019, 2019, 8267234. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhou, X.; Tan, X.; Huang, X.; Yuan, L.; Zhang, Z.; Yang, Y.; Xu, M.; Wan, Y.; Li, Z. The status of oxidative stress in patients with alcohol dependence: A meta-analysis. Antioxidants 2022, 11, 1919. [Google Scholar] [CrossRef]
- Lu, C.; Luo, J.; Liu, Y.; Yang, X. The oxidative stress responses caused by phthalate acid esters increases mRNA abundance of base excision repair (BER) genes in vivo and in vitro. Ecotoxicol. Environ. Saf. 2021, 208, 111525. [Google Scholar] [CrossRef]
- Garcia-Caparros, P.; Filippis, L.; Gul, A.; Hasanuzzaman, M.; Ozturk, M.; Altay, V.; Lao, M.T. Oxidative stress and antioxidant metabolism under adverse environmental conditions: A review. Bot. Rev. 2021, 87, 421–466. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Estresse oxidativo, envelhecimento e doenças. Interv. Clínicas No Envelhec. 2018, 13, 757–772. [Google Scholar]
- Polidori, M.C.; Mecocci, P. Modeling the dynamics of energy imbalance: The free radical theory of aging and frailty revisited. Free. Radic. Biol. Med. 2022, 181, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Liu, H.; Li, C. Dietary regulation of oxidative stress in chronic metabolic diseases. Foods 2021, 10, 1854. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: Chronic diseases and aging. Arch. Toxicol. 2023, 97, 2499–2574. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.P.R. Dissertação de Mestrado. UFVJM. Available online: http://acervo.ufvjm.edu.br/jspui/bitstream/1/1929/1/ana_paula_rufino_santos.pdf (accessed on 30 July 2021).
- Câmara, V.D.M.; Filhote, M.I.D.F.; Lima, M.I.M.; Alheira, F.V.; Martins, M.S.; Dantas, T.O.; Luiz, R.R. Methodology to prevent mercury exposure among adolescents from goldmine areas in Mariana, state of Minas Gerais, Brazil. Cad. De Saúde Pública 1996, 12, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.W.; Cockerham, L. Toxic metals in the environment. In Basic Environmental Toxicology, 1st ed.; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Ferrari, C.K. Functional foods, herbs and nutraceuticals: Towards biochemical mechanisms of healthy aging. Biogerontology 2004, 5, 275–289. [Google Scholar] [CrossRef]
- Fu, Z.; Xi, S. The effects of heavy metals on human metabolism. Toxicol. Mech. Methods 2020, 30, 167–176. [Google Scholar] [CrossRef]
- Gil, F.; Pla, A. Biomarkers as biological indicators of xenobiotic exposure. J. Appl. Toxicol. Int. J. 2001, 21, 245–255. [Google Scholar] [CrossRef]
- Chandra, S.; Chauhan, L.K.S.; Murthy, R.C.; Saxena, P.N.; Pande, P.N.; Gupta, S.K. Comparative biomonitoring of leachates from hazardous solid waste of two industries using Allium test. Sci. Total Environ. 2005, 347, 46–52. [Google Scholar] [CrossRef]
- Anlar, H.G.; Bacanli, M.; İritaş, S.; Bal, C.; Kurt, T.; Tutkun, E.; Yilmaz, H.; Basaran, N. Effects of occupational silica exposure on oxidative stress and immune system parameters in ceramic workers in Turkey. J. Toxicol. Environ. Health 2017, 80, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.P.R.; Silva, L.Z.; Freire, B.M.; da Silva Faria, M.C.; Batista, B.L.; Rocha, B.A.; Barbosa, F., Jr.; Rodrigues, J.L. Artisanal Gem Mining in Brazil: A Source of Genotoxicity and Exposure to Toxic Elements. Int. J. Environ. Res. Public Health 2023, 20, 2510. [Google Scholar] [CrossRef] [PubMed]
- Faria, M.C.S.; Santos, N.A.; Rodrigues, M.A.C.; Rodrigues, J.L.; Junior, F.B.; Santos, A.C. Effect of diabetes on biodistribution, nephrotoxicity and antitumor activity of cisplatin in mice. Chem. Biol. Interact. 2015, 229, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzym. 1984, 105, 121–126. [Google Scholar]
- Silva, F.M.R.J.; Rocha, J.A.V.; Vargas, V.M.F. Extraction parameters in the mutagenicity assay of soil samples. Sci. Total Environ. 2009, 407, 6017–6023. [Google Scholar]
- Grant, W.F. Chromosome aberration assays in Allium: A report of the US Environmental Protection Agency gene-tox program. Mutat. Res. Rev. Genet. Toxicol. 1982, 99, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Hott, R.C.; Magalhães, T.S.; Maia, L.F.; Santos, K.S.; Rodrigues, G.L.; Oliveira, L.C.; Pereira, M.C.; Faria, M.C.S.; Carli, A.P.; Alves, C.C.S.; et al. Purification of arsenic-contaminated water using iron molybdate filters and monitoring of their genotoxic, mutagenic, and cytotoxic effects through bioassays. Environ. Sci. Pollut. Res. 2021, 28, 5714–5730. [Google Scholar] [CrossRef]
- Mourão, A.O.; Silva, D.F.; Rodriguez, M.; Torres, T.S.; Franco, E.S.; Pádua, V.L.; Faria, M.C.S.; Maia, L.F.O.; Rodrigues, J.L. Degradation of haloacetic acids with the Fenton-like and analysis by GC-MS: Use of bioassays for monitoring of genotoxic, mutagenic and cytotoxic effects. Env. Monit Assess 2019, 191, 513. [Google Scholar] [CrossRef] [PubMed]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. Environ. Mutagen. Relat. Subj. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Sapkota, M.; Burnham, E.L.; Vasure, J.M.; Sweeter, J.M.; Hunter, C.D.; Duryee, M.J.; Klassen, L.W.; Kharbanda, K.K.; Sisson, J.H.; Thiele, G.M.; et al. Malondialdehyde–Acetaldehyde (MAA) Protein Adducts Are Found Exclusively in the Lungs of Smokers with Alcohol Use Disorders and Are Associated with Systemic Anti-MAA Antibodies. Alcohol. Clin. Exp. Res. 2017, 41, 2093–2099. [Google Scholar] [CrossRef]
- Mure, K.; Tomono, S.; Mure, M.; Horinaka, M.; Mutoh, M.; Sakai, T.; Ishikawa, H.; Wakabayashi, K. The combination of cigarette smoking and alcohol consumption synergistically increases reactive carbonyl species in human male plasma. Int. J. Mol. Sci. 2021, 16, 9043. [Google Scholar] [CrossRef]
- Bhagia, L.J. Non-occupational exposure to silica dust. Indian J. Occup. Environ. Med. 2012, 16, 95. [Google Scholar] [CrossRef]
- Satpathy, S.R.; Jala, V.R.; Bodduluri, S.R.; Krishnan, E.; Hegde, B.; Hoyle, G.W.; Fraig, M.; Luster, A.D.; Haribabu, B. Crystalline silica-induced leukotriene B 4-dependent inflammation promotes lung tumour growth. Nat. Commun. 2015, 6, 7064. [Google Scholar] [CrossRef] [PubMed]
- Marques Da Silva, V.; Benjdir, M.; Montagne, P.; Pairon, J.C.; Lanone, S.; Andujar, P. Pulmonary toxicity of silica linked to its micro-or nanometric particle size and crystal structure: A review. Nanomaterials 2022, 12, 2392. [Google Scholar] [CrossRef]
- Guidi, P.; Nigro, M.; Bernardeschi, M.; Lucchesi, P.; Scarcelli, V.; Frenzilli, G. Does the crystal habit modulate the genotoxic potential of silica particles? A cytogenetic evaluation in human and murine cell lines. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2015, 792, 46–52. [Google Scholar] [CrossRef] [PubMed]
- OSHA. Occupational Exposure to Respirable Crystalline Silica—Review of Health Effects Literature and Preliminary Quantitative Risk Assessment; OSHA: Tumwater, WA, USA, 2010. [Google Scholar]
- Nardi, J.; Nascimento, S.; Göethel, G.; Gauer, B.; Sauer, E.; Fão, N.; Cestonaroa, L.; Peruzzia, C.; Souza, J.; Garcia, S.C. Inflammatory and oxidative stress parameters as potential early biomarkers for silicosis. Clínica Chim. Acta 2018, 484, 305–313. [Google Scholar] [CrossRef]
- Amato, A.A.; Rosa, E.C.C.C.; Santos, R.R.C.; Neves, F.D.A.R.; Gris, E.F.; Ferreira, E.A.; Coelho, M.S. Evaluation of oxidative stress and lipoperoxidation (OLP) in patients with type 2 diabetes mellitus (DM2) treated at the University Hospital of Brasília (HUB). Braz. J. Health Rev. 2019, 2, 4236–4256. [Google Scholar]
- Malaei, R.; Ramezani, A.M.; Absalan, G. Analysis of malondialdehyde in human plasma samples through derivatization with 2, 4-dinitrophenylhydrazine by ultrasound-assisted dispersive liquid–liquid microextraction-GC-FID approach. J. Chromatogr. B 2018, 1089, 60–69. [Google Scholar] [CrossRef]
- Nikitina, O.A. Activity of Lipoperoxidation–Antioxidant Protection Reactions in Patients with HIV Infection. Acta Biomed. Sci. 2021, 5, 124–132. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Feldman, N.B.; Lutsenko, S.V. ROS and RNS signalling: Adaptive redox switches through oxidative/nitrosative protein modifications. Free. Radic. Res. 2018, 52, 507–543. [Google Scholar] [CrossRef]
- Khan, K.; Rafiq, M.T.; Bacha, A.U.R.; Nabi, I.; Irshad, M.; Faridullah, F.; Younas, M.; Daud Khan, M.; Aziz, R.; Amin, M.; et al. Assessment of heavy metals and associated oxidative stress in occupationally exposed workers from Bannu and Karak Districts in Pakistan. Environ. Geochem. Health 2023, 45, 5915–5925. [Google Scholar] [CrossRef] [PubMed]
- Pierezan, B.; Webber, B.; Vidmar, M.F.; Martins, C.A.D.Q.; Almeida, C.R.D.; Siqueira, L.D.O. Analysis of the oxidative profile of different biological samples of patients with anterior cruciate ligament injury. Fisioter. E Pesqui. 2017, 24, 198–204. [Google Scholar] [CrossRef]
- Martemucci, G.; Costagliola, C.; Mariano, M.; D’andrea, L.; Napolitano, P.; D’Alessandro, A.G. Free radical properties, source and targets, antioxidant consumption and health. Oxygen 2022, 2, 48–78. [Google Scholar] [CrossRef]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Several lines of antioxidant defense against oxidative stress: Antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch. Toxicol. 2024, 98, 1323–1367. [Google Scholar] [CrossRef]
- Altin, R.; Armutcu, F.; Kart, L.; Gurel, A.; Savranlar, A.; Özdemir, H. Antioxidant response at early stages and low grades of simple coal worker’s pneumoconiosis diagnosed by high resolution computed tomography. Int. J. Hyg. Environ. Health 2004, 207, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Tapak, M.; Sadeghi, S.; Ghazanfari, T.; Mosaffa, N. Chemical exposure and alveolar macrophages responses: The role of pulmonary defense mechanism in inhalation injuries. BMJ Open Respir. Res. 2023, 10, e001589. [Google Scholar] [CrossRef]
- Madkour, L.H. Function of reactive oxygen species (ROS) inside the living organisms and sources of oxidants. Pharm. Sci. Anal. Res. J. 2019, 2, 180023. [Google Scholar]
- Vallyathan, V.; Mega, J.F.; Shi, X.; Dalal, N.S. Enhanced generation of free radicals from phagocytes induced by mineral dusts. Am J Respir Cell Mol Biol. 1992, 6, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.P.S.; Maciel, E.; Ibiapina, C.R.L.S.; Moura, M.C. Implications of serum selenium concentrations on inflamatory bowel diseases: A bibliographic review. Electron. J. Collect. Health 2020, 12, 2101. [Google Scholar]
- Amin, M.N.; Siddiqui, S.A.; Uddin, M.G.; Ibrahim, M.D.; Uddin, S.N.; Adnan, M.T.; Rahaman, M.Z.; Kar, A.; Islam, M.S. Increased oxidative stress, altered trace elements, and macro-minerals are associated with female obesity. Biol. Trace Elem. Res. 2020, 197, 384–393. [Google Scholar] [CrossRef]
- Liu, Z.; Ren, Z.; Zhang, J.; Chuang, C.C.; Kandaswamy, E.; Zhou, T.; Zuo, L. Role of ROS and nutritional antioxidants in human diseases. Front. Physiol. 2018, 9, 477. [Google Scholar] [CrossRef]
- Kawasaki, H. A mechanistic review of silica-induced inhalation toxicity. Inhal. Toxicol. 2015, 27, 363–377. [Google Scholar] [CrossRef]
- Papp, L.V.; Holmgren, A.; Khanna, K.K. Selenium and Selenoproteins in Health and Disease. Antioxid. Redox Signal. 2010, 12, 793–795. [Google Scholar] [CrossRef]
- Sitta, A.; Vanzin, C.S.; Biancini, G.B.; Manfredini, V.; Oliveira, A.B.; Wayhs, C.A.Y.; Vargas, C.R. Evidence that L-carnitine and selenium supplementation reduces oxidative stress in phenylketonuric patients. Cell. Mol. Neurobiol. 2011, 31, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Tinggi, U. Selenium: Its role as antioxidant in human health. Environ. Health Prev. Med. 2008, 13, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.M.D.; Barbosa Júnior, F.; Júnior, A.A.J.; Foss, N.T.; Navarro, A.M.; Frade, M.A.C. Estresse oxidativo e micronutrientes na hanseníase. Rev. Nutr. 2015, 28, 349–357. [Google Scholar] [CrossRef]
- Tonel, D.; Silva, T.O.; Lazarotto, A.K.; Muller, D.A.; Lucca, L.D.; Ichikawa, T.T.D.; Gallina, A.L. Avaliação do status oxidativo, consumo alimentar de zinco e zinco plasmático em indivíduos infectados pelo HIV. Rev. Bras. Análises Clínicas 2018, 50, 153–160. [Google Scholar]
- Valla, F.V.; Bost, M.; Roche, S.; Pitance, M.; Cuerq, C.; Ridout, J.; Ecochard, R.; Ginhoux, T.; Bellon, A.; Ford-Chessel, C.; et al. Multiple micronutrient plasma level changes are related to oxidative stress intensity in critically ill children. Pediatr. Crit. Care Med. 2018, 19, 455–463. [Google Scholar] [CrossRef]
- Viana, A.C.C.; Maia, F.M.M.; Carvalho, N.S.D.; Morais, S.R.D.; Bezerra, A.N.; Mendonça, P.D.S.; Costa, S.L.D.; Daltro, A.F.C.S. Correlation between nutritional assessment and oxidative stress in candidates for liver transplant. Einstein 2019, 18, eAO4039. [Google Scholar] [CrossRef]
- Kumar, N.; Chugh, H.; Sood, D.; Singh, S.; Singh, A.; Awasthi, A.D.; Tomar, R.; Tomar, V.; Chandra, R. Biology of heme: Drug interactions and adverse drug reactions with CYP450. Curr. Top. Med. Chem. 2018, 18, 2042–2055. [Google Scholar] [CrossRef]
- Campos, M.T.G.; Leme, F.D.O.P. Oxidative stress: Pathophysiology and laboratory diagnosis. Pubvet 2017, 12, 1–8. [Google Scholar]
- Admas, T.; Kerisew, B. Assessment of cytotoxicity and genotoxicity potential of effluents from Bahir Dar Tannery using Allium cepa. Adv. Public Health 2022, 2022, 1–10. [Google Scholar] [CrossRef]
- Leme, D.M.; Marin-Morales, M.A. Allium cepa test in environmental monitoring: A review on its application. Mutat. Res. Rev. Mutat. Res. 2009, 682, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Carruyo, I.; Fernández, Y.; Marcano, L.; Montiel, X.; Torrealba, Z. Correlation of toxicity with lead content in root tip cells (Allium cepa L.). Biol. Trace Elem. Res. 2008, 125, 276–285. [Google Scholar] [CrossRef]
- Cunha Neto, A.R.; Silva, I.G.; Calvelli, J.V.B.; Martins, G.E.C.; Carvalho, M.; Barbosa, S. Toxicity of heavy metals that affect germination, development and cell cycle of Allium cepa L. Bull. Environ. Contam. Toxicol. 2023, 111, 22. [Google Scholar] [CrossRef]
- Moura, J.F.; Silva, M.S.; Batista, N.J.C.; Uchoa, V.T.; Alves, W.S. Study the Genotoxicity Of Extract Abelmoschus Esculentus (Okra) By Test Allium cepa. Saúde Foco 2014, 1, 15–28. [Google Scholar]
- Tabassum, J.A.S.; Pandey, D. Metal induced genotoxicity and oxidative stress in plants, assessment methods, and role of various factors in genotoxicity regulation. In Induced Genotoxicity and Oxidative Stress in Plants; Springer: Berlin/Heidelberg, Germany, 2021; pp. 133–149. [Google Scholar]
- Wierzbicka, M. The effect of lead on the cell cycle in the root meristem of Allium cepa L. Protoplasma 1999, 207, 186–194. [Google Scholar] [CrossRef]
- Achary, V.M.M.; Jena, S.; Panda, K.K.; Panda, B.B. Aluminium induced oxidative stress and DNA damage in root cells of Allium cepa L. Ecotoxicol. Environ. Saf. 2008, 70, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Singh, J.; Singh, J.; Singh, S.; Singh, S. Assessment of genotoxic effects of pesticide and vermicompost treated soil with Allium cepa test. Sustain. Environ. Res. 2018, 28, 171–178. [Google Scholar] [CrossRef]
- Sabeen, M.; Mahmood, Q.; Bhatti, Z.A.; Irshad, M.; Bilal, M.; Hayat, M.T.; Usman, I.; Tahir, A.A.; Muhammad, A.; Shahid, N. Allium cepa assay based comparative study of selected vegetables and the chromosomal aberrations due to heavy metal accumulation. Saudi J. Biol. Sci. 2020, 27, 1368–1374. [Google Scholar] [CrossRef]
- Reddy, K.R.; Chirakkara, R.A.; Ribeiro, L.F.M. Synergistic Co-contaminant Effects on Phytoremediation of Polluted Soils. Indian Geotech. J. 2024, 54, 315–328. [Google Scholar] [CrossRef]
- Matos, A.R.; Faria, M.C.S.; Freire, B.M.; Pereira, R.M.; Batista, B.L.; Rodrigues, J.L. Determination of 14 trace elements in blood, serum and urine after environmental disaster in the Doce River basin: Relationship between mining waste and metal concentration in the population. J. Trace Elem. Med. Biol. 2022, 70, 126920. [Google Scholar] [CrossRef] [PubMed]
- Abubacker, M.N.; Sathya, C. Genotoxic Effect of Heavy Metals Cr, Cu, Pb and Zn Using Allium Cepa L. Biosci. Biotechnol. Res. Asia 2017, 14, 1181–1186. [Google Scholar] [CrossRef]
- Galindo, T.; Silva, E.; Rosário, I. Indução de micronúcleos e toxicidade por efluente doméstico em duas populações de Bathygobius soporator (Valenciennes, 1837) (Teleostei, Gobbidae) no Litoral de Salvador (BA), Brasil. Braz. J. Aquat. Sci. Technol. 2012, 16, 1–7. [Google Scholar] [CrossRef]
- Xiong, J.; Yuan, B.F.; Feng, Y.Q. Mass spectrometry for investigating the effects of toxic metals on nucleic acid modifications. Chem. Res. Toxicol. 2019, 32, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Demir, E.; Castranova, V. Genotoxic effects of synthetic amorphous silica nanoparticles in the mouse lymphoma assay. Toxicol. Rep. 2016, 3, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Kopp, B.; Zalko, D.; Audebert, M. Genotoxicity of 11 heavy metals detected as food contaminants in two human cell lines. Environ. Mol. Mutagen. 2018, 59, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Wagner, J.R. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | 20–30 | 30–40 | 40–50 | 50–60 | 60–70 | 70–80 | Control | R2 |
|---|---|---|---|---|---|---|---|---|
| Fe | 569,634.89 | 583,184.53 | 572,925.90 | 549,363.50 * | 377,906.93 | 321,043.61 * | 580,573.83 | 0.08 |
| Cu | 980.42 | 972.83 * | 1154.78 | 937.96 * | 488.55 | 747.94 | 1109.77 | 0.00 |
| Zn | 6798.32 | 6610.94 | 6832.31 | 6801.54 | 4454.86 | 4473.60 | 7213.15 | 0.13 |
| Se | 62.98 | 49.63 * | 56.12 * | 49.61 * | 43.85 * | 28.49 * | 72.82 | 0.00 |
| Samples | Number of Cells in Interphase | Number of Cells in Mitosis | Average Root Growth | Inhibition Rate (CTI) % | Mitotic Index(MI) |
|---|---|---|---|---|---|
| 1C− | 4493 | 507 | 2.04 ± 0.56 | 0 | 10.48 ± 2.48 |
| 2C+ | 4296 | 704 | 1.21 ± 0.57 * | 40.53 | 14.10 ± 2.54 * |
| BOD P | 4391 | 609 | 1.77 ± 0.71 * | 18.42 | 12.18 ± 1.80 |
| BOD PP | 4393 | 607 | 1.66 ± 0.96 * | 18.66 | 12.38 ± 2.02 |
| BOD SS | 4381 | 619 | 1.91 ± 0.77 | 6.30 | 12.14 ± 1.98 |
| PIR P | 4301 | 699 * | 1.63 ± 0.71 * | 20.06 | 13.98 ± 2.35 * |
| PIR PP | 4309 | 691 * | 1.54 ± 0.78 * | 24.29 | 13.82 ± 2.38 * |
| PIR SS | 4358 | 642 | 1.50 ± 0.70 * | 26.33 | 12.84 ± 1.33 |
| PIN P | 4388 | 612 | 1.54 ± 0.59 * | 24.14 | 12.24 ± 2.01 |
| PIN PP | 4305 | 695 * | 2.18 ± 0.61 | −7.12 | 13.90 ± 2.32 * |
| PIN SS | 4402 | 598 | 2.16 ± 0.66 | −6.08 | 11.96 ± 1.70 |
| LAJ P | 4384 | 616 | 2.11 ± 0.66 | −3.74 | 12.32 ± 2.04 |
| LAJ PP | 4378 | 622 | 2.02 ± 0.67 | 1.04 | 12.44 ± 3.08 |
| LAJ SS | 4380 | 620 | 1.76 ± 0.48 * | 13.61 | 12.40 ± 2.24 |
| MAR P | 4364 | 636 * | 1.77 ± 0.81 | 12.90 | 12.72 ± 1.80 * |
| MAR PP | 4322 | 678 * | 2.13 ± 0.99 | −4.52 | 13.56 ± 2.36 * |
| MAR SS | 4397 | 603 | 1.44 ± 0.65 * | 29.47 | 12.06 ± 1.18 |
| Samples | Bridging | Loss | Breaking | Bridging and Loss | Bridging and Breaking | Loss and Breaking | C-Metaphase | Sticking | Total CA | Micronuclei |
|---|---|---|---|---|---|---|---|---|---|---|
| 1C− | 7 ± 0.48 | 1 ± 0.18 | 2 ± 0.36 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 4 ± 0.64 | 0 ± 0.0 | 12 ± 3.48 | 4 ± 0.48 |
| 2C+ | 24 ± 2.08 | 32 ± 2.66 | 10 ± 1.60 | 4 ± 0.79 | 0 ± 0.0 | 1 ± 0.20 | 70 ± 2.60 | 21 ± 1.88 | 162 ± 35.56 * | 31 ± 2.52 * |
| BOD P | 8 ± 0.36 | 5 ± 0.36 | 3 ± 0.52 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 20 ± 1.48 | 0 ± 0.0 | 36 ± 8.89 * | 5 ± 0.50 |
| BOD PP | 17 ± 1.36 | 20 ± 0.42 | 7 ± 1.04 | 0 ± 0.0 | 0 ± 0.0 | 3 ± 0.0 | 54 ± 1.36 | 0 ± 0.0 | 101 ± 24.47 * | 7 ± 0.56 |
| BOD SS | 10 ± 1.76 | 0 ± 0.0 | 1 ± 0.48 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 22 ± 1.10 | 0 ± 0.0 | 33 ± 9.56 | 8 ± 0.48 |
| PIR P | 15 ± 1.9 | 21 ± 1.92 | 18 ± 2.52 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 57 ± 2.90 | 8 ± 2.77 | 119 ± 27.36 * | 10 ± 0.80 |
| PIR PP | 9 ± 0.90 | 0 ± 0.0 | 7 ± 0.84 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 25 ± 1.40 | 5 ± 0.86 | 54 ± 12.22 * | 4 ± 0.48 |
| PIR SS | 10 ± 0.70 | 10 ± 1.20 | 6 ± 0.98 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 27 ± 0.0 | 5 ± 0.0 | 58 ± 13.16 * | 3 ± 0.42 |
| PIN P | 15 ± 0.70 | 4 ± 0.64 | 4 ± 0.64 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 27 ± 1.30 | 4 ± 0.72 | 54 ± 13.33 * | 4 ± 0.48 |
| PIN PP | 11 ± 1.32 | 2 ± 0.32 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 21 ± 0.92 | 6 ± 0.96 | 40 ± 10.07 * | 9 ± 0.54 |
| PIN SS | 17 ± 1.70 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 6 ± 1.08 | 0 ± 0.0 | 27 ± 1.64 | 4 ± 0.72 | 57 ± 14.0 * | 6 ± 0.7 |
| LAJ P | 19 ± 1.30 | 5 ± 0.96 | 7 ± 0.98 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 45 ± 1.10 | 0 ± 0.0 | 76 ± 19.85 * | 8 ± 0.48 |
| LAJ PP | 11 ± 0.90 | 11 ± 1.12 | 5 ± 0.70 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 30 ± 1.68 | 12 ± 1.68 | 69 ± 15.18 * | 9 ± 0.57 |
| LAJ SS | 23 ± 1.76 | 0 ± 0.0 | 3 ± 0.48 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 20 ± 1.1 | 14 ± 1.48 | 60 ± 14.14 * | 7 ± 0.4 |
| MAR P | 22 ± 2.0 | 7 ± 0.44 | 5 ± 0.70 | 0 ± 0.0 | 3 ± 0.54 | 0 ± 0.0 | 9 ± 1.08 | 0 ± 0.0 | 46 ± 10.56 | 4 ± 0.56 |
| MAR PP | 15 ± 1.4 | 13 ± 1.82 | 11 ± 1.12 | 0 ± 0.0 | 0 ± 0.0 | 0 ± 0.0 | 19 ± 1.50 | 6 ± 0.96 | 64 ± 12.30 * | 7 ± 0.56 |
| MAR SS | 14 ± 1.68 | 9 ± 0.90 | 8 ± 0.98 | 3 ± 0.59 | 0 ± 0.0 | 5 ± 0.90 | 31 ± 1.88 | 7 ± 0.54 | 77 ± 16.40 * | 7 ± 0.42 |
| Samples | Chromosomal Aberrations | p Value | Average Root Growth | p Value |
|---|---|---|---|---|
| BOD P vs. PIR P | Yes | <0.001 | No | >0.999 |
| BOD PP vs. PIR PP | Yes | 0.049 | No | >0.999 |
| BOD PP vs. PIN PP | Yes | <0.001 | No | 0.240 |
| BOD SS vs. MAR SS | Yes | 0.023 | No | 0.193 |
| PIR P vs. LAJ P | Yes | <0.001 | No | 0.125 |
| PIR P vs. MAR P | Yes | <0.001 | No | >0.999 |
| PIR PP vs. PIN PP | Yes | <0.001 | Yes | 0.016 |
| MAR PP vs. PIR PP | No | >0.999 | Yes | 0.028 |
| MAR SS vs. PIN SS | No | 0.806 | Yes | 0.005 |
| PIR SS vs. PIN SS | No | >0.99 | Yes | 0.019 |
| PIN P vs. LAJ P | No | 0.148 | Yes | 0.013 |
| Samples | No Metabolization | With Metabolization | ||||||
|---|---|---|---|---|---|---|---|---|
| TA100 a | MR | TA 98 b | MR | TA 100 c | MR | TA98 d | MR | |
| BOD | 61 ± 2.0 | 3.21 | 69 ± 2.7 | 3.83 | 85.5 ± 2.9 | 2.80 | 88 ± 2.5 | 2.51 |
| PIR | 65.5 ± 4.5 | 3.45 | 38 ± 4.0 | 2.11 | - | - | - | - |
| PIN | 64 ± 2.0 | 3.37 | 40 ± 2.0 | 2.22 | - | - | - | - |
| LAJ | 71 ± 3.0 | 3.74 | 69 ± 3.6 | 3.83 | 72 ± 5.5 | 2.36 | 80.05 ± 2.7 | 2.29 |
| MAR | 76 ± 4.0 | 4.00 | 39.5 ± 0.5 | 2.19 | 65 ± 3.1 | 2.13 | 78 ± 3.0 | 2.23 |
| C− | 19 ± 3.0 | - | 18 ± 2.0 | - | 30.5 ± 3.3 | - | 35 ± 2.5 | - |
| C+ | 84 ± 5.1 | - | 71 ± 3.3 | - | 201.5 ± 5.1 | - | 98 ± 3.5 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, H.T.; Magalhães, T.S.; Pires, S.A.; Santos, A.P.R.; Rodrigues, J.L.; Faria, M.C.d.S. Artisanal Gem Mining in Brazil: Evaluation of Oxidative Stress and Genotoxicity Biomarkers. Int. J. Environ. Res. Public Health 2024, 21, 871. https://doi.org/10.3390/ijerph21070871
da Silva HT, Magalhães TS, Pires SA, Santos APR, Rodrigues JL, Faria MCdS. Artisanal Gem Mining in Brazil: Evaluation of Oxidative Stress and Genotoxicity Biomarkers. International Journal of Environmental Research and Public Health. 2024; 21(7):871. https://doi.org/10.3390/ijerph21070871
Chicago/Turabian Styleda Silva, Heberson Teixeira, Thainá Sprícido Magalhães, Sumaia Araújo Pires, Ana Paula Rufino Santos, Jairo Lisboa Rodrigues, and Márcia Cristina da Silva Faria. 2024. "Artisanal Gem Mining in Brazil: Evaluation of Oxidative Stress and Genotoxicity Biomarkers" International Journal of Environmental Research and Public Health 21, no. 7: 871. https://doi.org/10.3390/ijerph21070871
APA Styleda Silva, H. T., Magalhães, T. S., Pires, S. A., Santos, A. P. R., Rodrigues, J. L., & Faria, M. C. d. S. (2024). Artisanal Gem Mining in Brazil: Evaluation of Oxidative Stress and Genotoxicity Biomarkers. International Journal of Environmental Research and Public Health, 21(7), 871. https://doi.org/10.3390/ijerph21070871