
Mineral Contents in Aboveground Biomass of Sedges (Carex L., Cyperaceae)

 ,
,
Abstract
:1. Introduction
2. Data Sources and Methodology
2.1. The Soil Properties Analysis Method
2.2. Elemental Analysis Method
2.3. Statistical Analysis
3. Results and Discussion
3.1. Results
3.1.1. Physicochemical Properties of the Soil
3.1.2. Mineral Composition of the Studied Plants
3.1.3. Macromineral Concentration in Studied Sedges
3.1.4. Micromineral Concentration in Studied Sedges
3.2. Discussion
4. Conclusions
- According to the hypothesis, studied sedges are a good source of macroelements and microelements.
- Species characterized by the highest numbers of constituents at optimal level are namely: C. diandra, C. lepidocarpa and C. pseudocyperus.
- None of the examined species contained the optimal amount of all analysed elements.
- Generally, the studied sedges contained excess Fe and Mn and lacked P, Cu and Zn.
- The elemental composition of studied Carex is diversified but is generally similar to the composition of the meadow grasses.
- Aboveground parts of Carex are suitable for fodder purposes or in order to supplement the deficits in particular components.
- Sedges can increase the fodder value and can be used in meadow management.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goetghebeur, P. Cyperaceae. In The Families and Genera of Vascular Plants. Monocotyledons, 4th ed.; Springer: Hamburg, Germany, 1998. [Google Scholar]
- Egorova, T.V. The Sedges Carex L. of Russia and Adjacent States; State Chemical-Pharmaceutical Academy St Petersburg: St Petersburg, Russia, 1999. [Google Scholar]
- Janyszek, M.; Jagodzinski, A.M.; Janyszek, S.; Wronska-Pilarek, D. Morphological variability of Carex spicata Huds utricles among plant communities. Flora 2008, 203, 386–395. [Google Scholar] [CrossRef]
- Govaerts, R.; Simpson, D.A.; Bruhl, J. World Checklist of Cyperaceae; Royal Botanic Gardens Kew: London, UK, 2009. [Google Scholar]
- Janyszek, M.; Jagodzinski, A.M. Variability of perigynium morphology of Central European members of Carex sect Phaestoglochin Cyperaceae from variable plant communities. Plant Syst. Evol. 2009, 278, 87–99. [Google Scholar] [CrossRef]
- Catling, P.M.; McElroy, A.R.; Spicer, K.W. Potential forage value of some eastern Canadian sedges Cyperaceae. Carex. J. Rang. Manag. 1994, 47, 226–230. [Google Scholar] [CrossRef]
- Larter, N.C.; Nagy, J.A.; Hik, D.S. Does sesonal variation in forage quality influence the potential for resource competition between muskoxen and Peary Caribou on Banks Island? Ragnifer 2002, 22, 143–153. [Google Scholar]
- Clark, P.E. Date and plant community effects on elk sedge forage quality. J. Rang. Manag. 2003, 56, 21–26. [Google Scholar] [CrossRef]
- Meehan, M.A.; DeKeyser, E.S.; Sedivec, K.K.; Norland, J.E. Nutrition composition of Sprengel’s sedge Carex sprengelii. Can. J. Plant Sci. 2012, 92, 867–871. [Google Scholar] [CrossRef]
- Alldrege, M.W.; Peek, J.M.; Wall, W.A. Nutritional quality of forages used by elk in Northern Idaho. J. Rang. Manag. 2002, 55, 253–259. [Google Scholar] [CrossRef]
- Choo, Y.S.; Lee, C.B.; Albert, R. Effects of nitrogen nutrition on the pattern of ions and organic solutes in five sedges (Carex spp.). Flora 2002, 197, 56–66. [Google Scholar] [CrossRef]
- Ringqvist, L.; Oborn, I. Cooper and zinc adsorption onto poorly humified Sphagnum and Carex peat. Water Res. 2002, 36, 2233–2242. [Google Scholar] [CrossRef]
- Ngai, J.T.; Jeffers, R.L. Nutrient limitation of plant growth and forage quality in Arctic coastal marshes. J. Ecol. 2004, 92, 1001–1010. [Google Scholar] [CrossRef]
- Krasicka-Korczyńska, E.; Dembek, R.; Korczyński, M.; Stosik, T. Natural and fodder values of community with Carex nigra in the Bydgoszcz Canal Valley. Acta Sci. Pol. Agric. 2014, 134, 77–91. [Google Scholar]
- Veselkin, D.V.; Konoplenko, M.A.; Betekhtina, A.A. Means for soil nutrient uptake in sedges with different ecological strategies. Russ. J. Ecol. 2014, 45, 547–554. [Google Scholar] [CrossRef]
- Nekhoroshkov, P.S.; Kravtsova, A.V.; Kamnev, A.N.; Duliu, O.; Bunkova, O.M.; Frontasyeva, M.V.; Yermakov, I.P. Assessment of major and trace elements in aquatic macrophytes soils and bottom sediments collected along different water objects in the lack Sea Coastal zone by using Neutron Activation Analysis. Mod. Chem. Appl. 2017, 5, 1. [Google Scholar]
- Li, F.; Gao, H.; Zhu, L.; Xie, Y.; Yang, G.; Hu, C.; Deng, Z. Foliar nitrogen and phosphorus stoichiometry of three wetland plants distributed along an elevation gradient in Dongting Lake, China. Sci. Rep. 2017, 7, 1–9. [Google Scholar]
- Parzych, A.E.; Sobisz, Z.; Jonczak, J. Comparing Carex species of mid-forest spring ecosystem in terms of ability to accumulate macro- and microelements. J. Ecol. Eng. 2017, 18, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Nie, L.; Xu, Y.; Li, M.; Lv, Y. Traffic-emitted metal status and uptake by Carex meyeriana Kunth and Thelypteris palustris var. pubescens Fernald growing in roadside turfy swamp in the Changbai Mountain area, China. Environ. Sci. Pollut. Res. Int. 2018, 25, 18498–18509. [Google Scholar]
- Hinzke, T.; Tanneberger, F.; Aggenbach, C.; Dahlke, S.; Knorr, K.H.; Kotowski, W.; Kozub, Ł.; Lange, J.; Li, G.; Pronin, E.; et al. Can nutrient uptake by Carex counteract eutrophication in fen peatlands? Sci. Total Environ. 2021, 785, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodriguez, C.C.; Izquiredo-Ramirez, C.R.; Lanfranco-Colina, L.J.; Ciraco-Mosqueira, J.E.; Cervantes-Peralta, M.E.; Sanchez-Pena, M.A. Phytoextraction of heavy metals from the soil of aurora-patricia mining environmental liability by herbaceous species Carex mandoniana, Equisetum bogotense, and Muehlenbeckia tamnifolia, La Encañada-Peru 2020. Conference paper. In Proceedings of the Brasilian Technology Symposium, Virtual Event, 8–10 November 2021. [Google Scholar]
- Zhao, L.; Ma, X.; Liu, H.Q.; Xiong, L.H.; Guo, X.L.; Li, L.P.; Dong, L.Q.; Zhang, K. Responses of leaf carbon, nitrogen, and phosphorus stoichiometry of Carex muliensis to water table drawdown in an alpine marsh on the Ruoergai Plateau, China. J. Appl. Ecol. 2021, 32, 2426–2432. [Google Scholar]
- Bennett, J.P.; Chiriboga, E.; Coleman, V.; Waller, D.M. Heavy metals in wild rice from northern Wisconsin. Sci. Total Ennviron. 2000, 246, 261–269. [Google Scholar] [CrossRef]
- Ozcan, M. Mineral contents of some plants used as condiments in Turkey. Food Chem. 2003, 84, 437–440. [Google Scholar] [CrossRef]
- Juknevicius, S.; Sabiene, N. The content of mineral elements in some grasses and legumes. Ekologija 2007, 53, 44–52. [Google Scholar]
- Soetan, K.O.; Olaiya, C.O.; Oyewole, O.E. The importance of mineral elements for humans domestic animals and plants, A review. Afr. J. Food Sci. 2010, 45, 200–222. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants III; Academic Press: London, UK, 2011. [Google Scholar]
- Babu, R.H.; Savithramma, N. Studies on mineral analysis of grasses of Poaceae. Int. J. Pharma Sci. 2014, 4, 526–531. [Google Scholar]
- Grzegorczyk, S.; Alberski, J.; Olszewska, M.; Grabowski, K.; Bałuch-Małecka, A. Content of calcium and phosphorus and the Ca, P ratio in selected species in leguminous and herbaceous plants. J. Elem. 2017, 22, 663–669. [Google Scholar]
- Wyłupek, T. Rolnicza charakterystyka zespołu Caricetum acutae Tx1937 z doliny Poru. Woda-Środ-Obsz Wiej. 2001, 1, 171–183. (In Polish) [Google Scholar]
- Bengtsson, H.; Oborn, L.; Jonsson, S.; Nilsson, I.; Andersson, A. Field balances of some mineral nutrients and trace elements in organic and conventional dairy farming—A case study at Ojebyn Sweden. Eur. J. Agron. 2003, 20, 101–116. [Google Scholar] [CrossRef]
- Warman, P.R.; Termeer, W.C. Evaluation of sewage sludge septic waste and sludge compost applications to corn and forage, yields and N P and K content of crops and soils. Bioresour. Technol. 2005, 96, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S. The mineral nutrition of wild plants. Ann. Rev. Ecol. Syst. 1980, 11, 233–260. [Google Scholar] [CrossRef]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soil and Plants, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Malzahn, E. Igły sosny zwyczajnej jako bioindykator zagrożeń środowiska leśnego Puszczy Białowieskiej. Biul. Monit. Przyr. 2002, 1, 24–31. (In Polish) [Google Scholar]
- Ostrowska, A.; Porębska, G. Skład Chemiczny Roślin Jego Interpretacja i Wykorzystanie w Ochronie Środowiska; Inst Ochr Środ.: Warszawa, Poland, 2002. (In Polish) [Google Scholar]
- Parzych, A. Azot fosfor i węgiel w roślinności leśnej Słowińskiego Parku Narodowego w latach 2002–2005. Ochr. Środ. Zas. Nat. 2010, 43, 45–64. (In Polish) [Google Scholar]
- Demeyer, A.; Voundi Nkana, J.C.; Verloo, M.G. Characteristics of wood ash and influence on soil properties and nutrient uptake, an overview. Biores. Technol. 2001, 77, 287–295. [Google Scholar] [CrossRef]
- Mandre, M.; Korsjukov, R.; Ots, K. Effect of wood ash on the biomass distribution and physiological state of Norway spruce seedlings on sandy soils. Plant Soil 2004, 265, 301–314. [Google Scholar] [CrossRef]
- Saarsalmi, A.; Malkonen, E.; Kukkola, M. Effects of wood ash fertilization on soil chemical properties and stand nutrient status and growth of some coniferous stands in Finland. Scand. J. For. Res. 2004, 19, 217–233. [Google Scholar] [CrossRef]
- Mandre, M. Influence of wood ash on soil chemical composition and biochemical parameters of young Scots pine. Proc. Estonian Acad. Sci. Biol. Ecol. 2006, 55, 91–107. [Google Scholar] [CrossRef]
- Doongar, R.C.; Ghosh, A. Bioaccumulation of nutrient elements from fly ash-amended soil in Jatropha curcas L., a biofuel crop. Environ. Monit. Assess. 2013, 185, 6705–6712. [Google Scholar]
- Brannvall, E.; Wolters, M.; Sjoblom, R.; Kumpiene, J. Elements availability in soil fertilized with pelletized fly ash and biosolids. J. Environ. Manag. 2015, 159, 27–36. [Google Scholar] [CrossRef] [PubMed]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, up-Date 2015 International Soil Classification System for Naming Soil and Creating Leg-Ends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Mocek, A.; Drzymała, S. Geneza, Analiza i Klasyfikacja Gleb; Wyd. UP Poznań: Poznań, Poland, 2010. (In Polish) [Google Scholar]
- Bojko, O.; Kabała, C. Organic carbon pools in mountain soils—Sources of variability and predicted changes in relation to climate and land use changes. Catena 2017, 149, 209–220. [Google Scholar] [CrossRef]
- Broadley, M.R.; Bowen, H.C.; Cotterill, H.L.; Hammond, J.P.; Meacham, M.C.; Mead, A.; White, P.J. Phylogenetic variation in the shoot mineral concentration of angiosperms. J. Exp. Bot. 2004, 33, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Domański, P. Poradnik dla Użytkowników łąk i Pastwisk; Prodruk: Poznań, Poland, 1998. (In Polish) [Google Scholar]
- Falkowski, M.; Kukułka, I.; Kozłowski, S. Właściwości Chemiczne Roślin Łąkowych; Wydawnictwo AR w Poznaniu: Poznań, Poland, 2000. (In Polish) [Google Scholar]
- NRC (National Research Council). Nutrient Requirements of Beef Cattle, 2000 ed.; National Academy Press: Washington, DC, USA, 2000; Available online: http//www.nap.edu/books/0309069343/html (accessed on 12 September 2021).
- NRC (National Research Council). Nutrient Requirement of Dairy Cattle, 2001 ed.; Academic Press: Washington, DC, USA, 2001; Available online: http,//www.nap.edu/catalog/9825.html (accessed on 12 September 2021).
- Smólczyński, S.; Orzechowski, M.; Kalisz, B.; Sowiński, P. Content of selected elements and exchangeable cations in soils formed from glacio-lacustrine sediments. J. Elem. 2020, 25, 347–361. [Google Scholar]
- Zharikova, E.A. Geochemical characterization of soils of the eastern coast of the Northern Sakhalin Lowland. Eurasian Soil Sci. 2017, 50, 34–41. [Google Scholar] [CrossRef]
- Smólczyński, S.; Orzechowski, M.; Kalisz, B.; Sowiński, P. Content of selected elements and exchangeable cations in soils developed from glacio-lacustrine sediments of Sępopolska Plain (NE Poland). J. Elem. 2020, 20, 417–434. [Google Scholar]
- Glina, B.; Kołodziejczyk-Prażmo, K.; Bogacz, A.; Tomaszewska, K.; Kaczmarek, Z.; Gajewski, P. Application of Shumann and Joosten classification in fen peatland degradation stage assessment—A case study from southern Poland. Ecol. Quest. 2017, 27, 89–96. [Google Scholar]
- Glina, B.; Gajewski, P.; Mendyk, Ł.; Zawieja, B.; Kaczmarek, Z. Recent changes in soil properties and carbon stocks in fen peatlands adjacent to open-pit lignite mines. Land Degrad. Dev. 2019, 30, 2371–2380. [Google Scholar] [CrossRef] [Green Version]
- Wiesmeier, M.; Urbansk, L.; Hobley, E.; Lang, B.; von Lützow, M.; Marin-Spiotta, E.; Kögel-Knabner, I. Soil organic carbon storage as a key function of soils—A review of drivers and indicators at various scales. Geoderma 2019, 333, 149–162. [Google Scholar] [CrossRef]
- Liu, H.; Janssen, M.; Lennartz, B. Changes in flow and transport patterns in fen peat following soil degradation. Eur. J. Soil Sci. 2016, 67, 763–772. [Google Scholar] [CrossRef]
- Jarnuszewski, G.; Meller, E. Total content of macroelements and trace elements in Holocene calcareous gyttja from the post-bog area of North-western Poland. Soil Water Res. 2019, 14, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Smólczyński, S.; Orzechowski, M.; Kalisz, B. Distribution of elements in soil catenas developed in ice-dammed lake and morainic landscapes in NE Poland. J. Elem. 2015, 25, 347–361. [Google Scholar]
- Pirhofer-Walzl, K.; Søegaard, K.; Høgh-Jensen, H.; Eriksen, J.; Sanderson, M.A.; Rasmusen, J.; Rasmussen, J. Forage herbs improve mineral composition of grassland herbage. Grass Forage Sci. 2011, 66, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Elgersma, A.; Soegaard, K.; Jensen, S.K.; Sehested, J. Herbage mineral contents in grass and legume species. Conference paper, Sustainable meat and milk production from grasslands. In Proceedings of the 27th General Meeting of the European Grassland Federation, Cork, Ireland, 17–21 June 2018; pp. 169–171. [Google Scholar]
- Whitehead, D.C. Nutrient Elements in Grasslands, Soil-Plant-Animal Relationship; CABI Publishing: Wallingford, UK, 2000. [Google Scholar]
- Goliński, P.; Spychalski, W. Effect of Trifolium pratense cultivars on yield and chemical composition of grass-legume swards Workshop on dynamics N-flows and forage utilization in legume-based systems. Grado 2005, 2, 45–49. [Google Scholar]
- Bucinova, K.; Jamnicka, G.; Havrantova, J.; Kukla, J.; Kuklova, M. Content of Selected Elements in Aboveground of Parts of Carex Pilosa Scop in Sporocarps of Some Microfungi at the Ecological Experimental Site in the Kremnicke Vrchy Mts Bratislava; Slovak Library of Forestry and Wood Sciences: Bratislava, Slovakia, 2008. [Google Scholar]
- Czerwiński, Z.; Pracz, J. Content of mineral components in the over-ground parts of herb layer plants in the Sphagno girgensohnii-Piceetum community. Pol. Ecol. Stud. 1995, 21, 195–205. [Google Scholar]
- Marinas, A.; Garcia-Gonzalez, R. Preliminary data on nutritional value of abundand species in supraforestal pyrenean pastures. Pirineos 2006, 161, 85–109. [Google Scholar] [CrossRef]
- Jankowska-Hufejt, H.; Wróbel, B.; Barszczewski, J. Ocena wartości pokarmowej pasz z użytków zielonych na tle zasobności gleb i bilansu składników N P K w wybranych gospodarstwach ekologicznych. J. Res. Appl. Agric. Eng. 2000, 543, 95–102. (In Polish) [Google Scholar]
- NRC (National Research Council). Nutrient Requirements of Beef Cattle, 1996 ed.; National Academy Press: Washington, DC, USA, 1996; Available online: http://www.nap.edu/books/0309069343/html (accessed on 15 September 2021).
- Ayan, I.; Mut, H.; Onal-Asci, O.; Basaran, U.; Acar, Z. Effects of manure application on the chemical composition of rangeland hay. J. Anim. Vet. Adv. 2010, 9, 1852–1857. [Google Scholar]
- Kumar, K.; Soni, A. Elemental ratio and their importance in feed and fodder. Int. J. Pure App. Biosci. 2014, 2, 154–160. [Google Scholar]
- Metson, A.J.; Gibson, E.J.; Hunt, J.L.; Sauders, W.M.H. Seasonal variations in chemical composition of pasture. N. Zealand J. Agric. Res. 1979, 22, 309–318. [Google Scholar] [CrossRef]
- Grzegorczyk, S.; Alberski, J. Zawartość niektórych mikroelementów w wybranych gatunkach ziół łąkowo-pastwiskowych. Zesz. PTPN. 2000, 471, 705–710. (In Polish) [Google Scholar]
- Warda, M.; Ćwintal, B. Zawartość boru miedzi żelaza manganu i cynku w wybranych gatunkach roślin pastwiskowych. Zesz. PTPN. 2000, 471, 841–846. (In Polish) [Google Scholar]
- Loneragan, J.F. The Availability and Absorption of Trace Elements in Soil-Plant Systems and Their Relation to Movement and Concentration of Trace Elements in Plants, in Trace Elements in Soil-Plant Animal Systems; Nicholas, D.J.D., Egan, A.R., Eds.; Academic Press: New York, NY, USA, 1975. [Google Scholar]
- Piekarczyk, M.; Kotwica, K.; Jaskólski, D. The elemental composition of ash from straw and hay in the content of their agricultural utilization. Acta Sci. Pol. 2011, 10, 97–104. [Google Scholar]
- Waliszewska, B.; Janyszek, M.; Grzelak, M.; Gaweł, E. Characteristic of thermochemical parameters of aboveground parts of selected sedges (Carex L. Cyperaceae). IOSR-JAVS 2013, 5, 1–6. [Google Scholar] [CrossRef]
- Grzelak, M.; Gaweł, E.; Majchrzak, L. Diversity of floristic, habitat and energy value of selected species of genus Carex L. Acta Sci. Pol. 2019, 18, 181–187. [Google Scholar]
- Waliszewska, B.; Grzelak, M.; Gaweł, E.; Spek-Dźwigała, A.; Sieradzka, A.; Czekała, W. Chemical characteristic of selected grass species from Polish meadows and their potential utilization for energy generation purposes. Energies 2021, 14, 1669. [Google Scholar] [CrossRef]
- Murawski, M.; Grzelak, M.; Waliszewska, B.; Knioła, A.; Czekała, W. Energy value and yelding from extensively used meadows. Fragm. Agron. 2015, 32, 71–78. [Google Scholar]
- Szczepanik, M.; Szyszlak-Bargłowicz, J.; Zając, G.; Koniuszy, H.; Hawrot-Paw, M.; Wolak, A. The Use of Multivariate Data Analysis (HCA and PCA) to Characterize Ashes from Biomass Combustion. Energies 2021, 14, 6887. [Google Scholar] [CrossRef]
- Zając, G.; Szyszlak-Bargłowicz, J.; Gołębiowski, W.; Szczepanik, M. Chemical Characteristics of Biomass Ashes. Energies 2018, 11, 2885. [Google Scholar] [CrossRef] [Green Version]
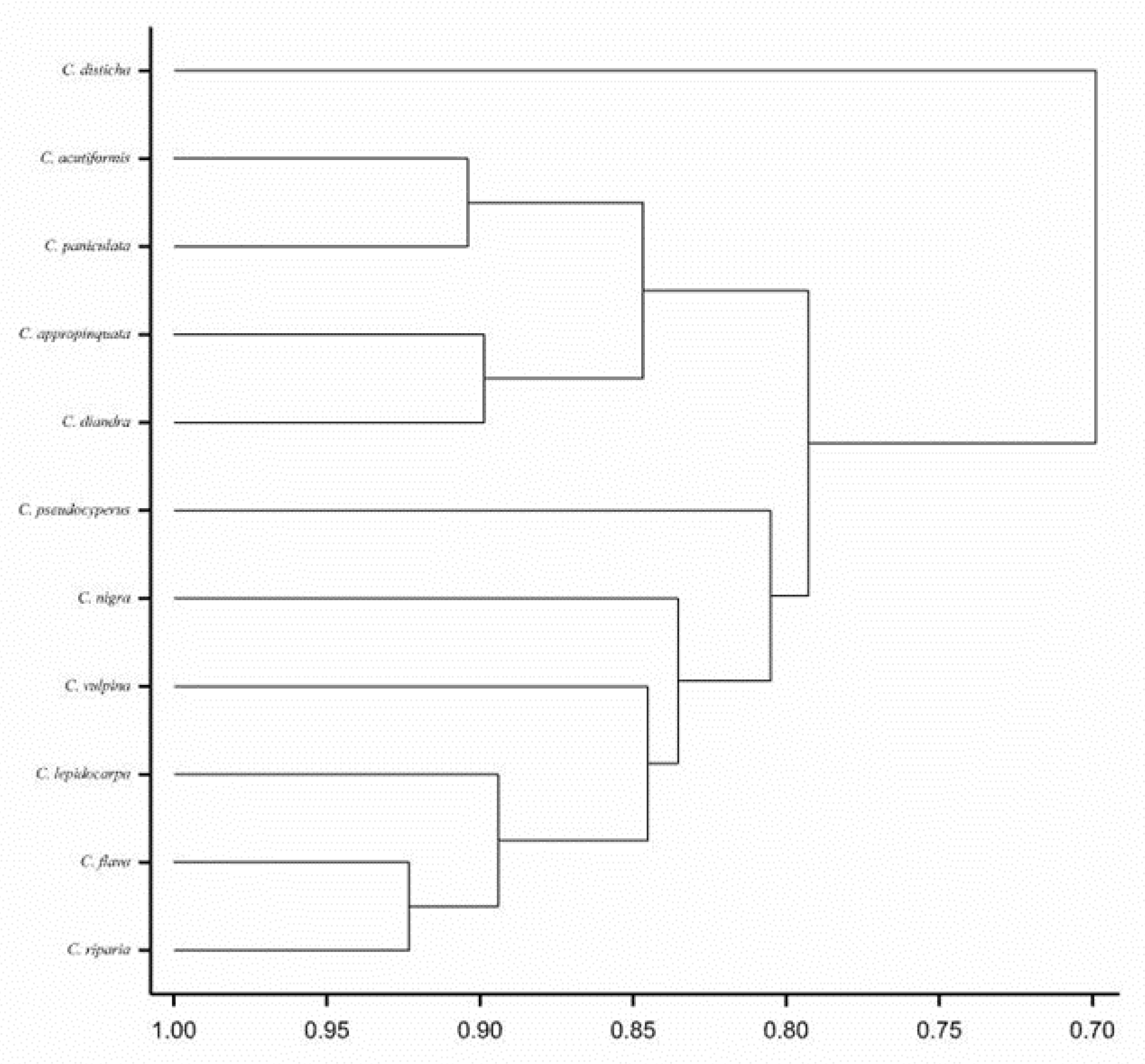

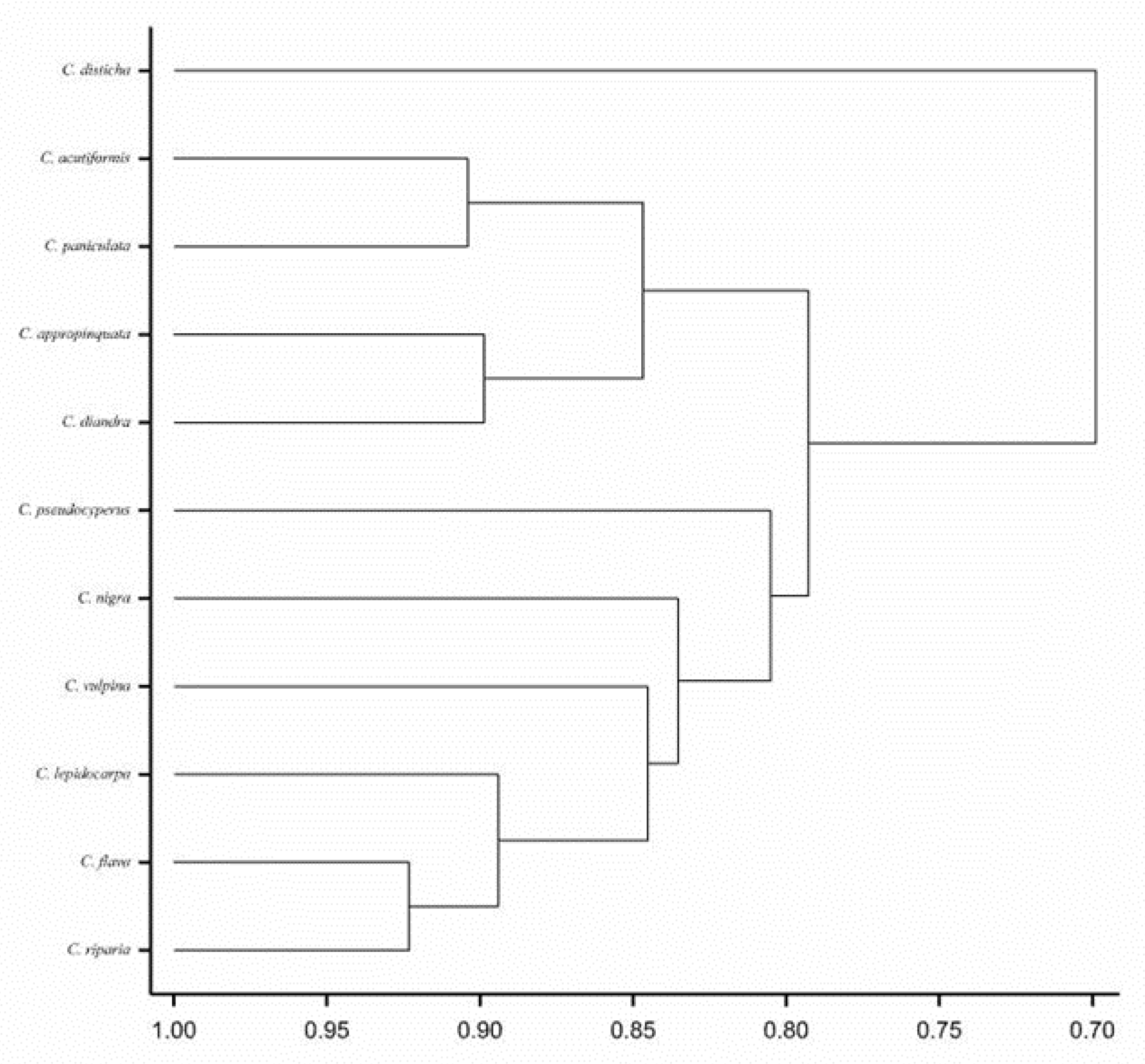{kind=link}
| No. of Site | Species | Genetic Horizon Acc, to FAO-WRB | Soil Material/Texture | Depth (cm) | pHe in 1MKCl | C tot, (g·kg−1) | N tot, (g·kg−1) | C–N | OMC (g·kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | C. acutiformis | Ha1 | sapric peat | 0–16 | 5.44 g | 223.3 d | 17.10 cd | 13.1 c | 430.1 e |
| Ha2 | sapric peat | 16–31 | 6.61c d | 203.6 e | 18.30 c | 11.1 d | 408.3 e | ||
| 2 | C. appropinquata | Ha1 | murshic | 0–27 | 6.72 c | 274.1 cd | 25.50 b | 10.7 de | 621.3 cd |
| Ha2 | murshic | 27–42 | 5.84 e | 324. 5b | 28.60 ab | 11.3 d | 614.3 d | ||
| 3 | C. diandra | He1 | hemic peat | 0–20 | 5.41 g | 294.4 c | 27.40 b | 10.7 de | 642 c |
| He2 | hemic peat | 20–160 | 5.63 f | 446.6 a | 29.90 a | 14.9 b | 802.3 a | ||
| 4 | C. disticha | A | sand | 0–32 | 5.68 ef | 11.1 j | 0.90 i | 12.3 c | 27.40 k |
| Bw | silt | 32–72 | 5.84 e | 5.6 k | 0.50 i | 11.2 d | 15.30 l | ||
| 5 | C. flava | Ha1 | sapric peat | 0–15 | 5.94 e | 372.8 b | 32.10 a | 11.6 cd | 712.4 b |
| Ha2 | sapric peat | 15–33 | 6.72 c | 193.2 e | 17.90 c | 10.8 de | 302.6 fg | ||
| 5 | C. lepidocarpa | Ha1 | sapric peat | 0–15 | 5.94 e | 372.8 b | 32.10 a | 11.6 cd | 712.4 b |
| Ha2 | sapric peat | 15–33 | 6.72 c | 193.2 e | 17.90 c | 10.8 de | 302.6 fg | ||
| 6 | C. nigra | Ha1 | murshic | 0–31 | 7.51 a | 127.3 fg | 11.70 e | 10.9 de | 230.7 g |
| Ha2 | sapric peat | 31–36 | 7.04 b | 150.2 f | 11.50 e | 13.1 c | 326.2 f | ||
| 2 | C. paniculata | Ha1 | murshic | 0–27 | 6.72 c | 274.1 cd | 25.50 b | 10.7 de | 621.3 cd |
| Ha2 | murshic | 27–42 | 5.84 e | 324.5 b | 28.60 ab | 11.3 d | 614.3 d | ||
| 7 | C. pseudocyperus | Ha1 | murshic | 0–26 | 6.18 d | 174.5 ef | 16.30 d | 10.7 de | 323.4 f |
| Ha2 | sapric peat | 26–55 | 7.35 ab | 289.3 c | 17.7 cd | 16.3 a | 619.3 d | ||
| 7 | C. riparia | Ha1 | murshic | 0–26 | 6.18 d | 174.5 ef | 16.30 d | 10.7 de | 323.4 f |
| Ha2 | sapric peat | 26–55 | 7.35 ab | 289.3 c | 17.7 cd | 16.3 a | 619.3 d | ||
| 8 | C. vulpina | Ah1 | silt | 0–13 | 7.63 a | 46.2 h | 5.10 g | 9.1 f | 88.9 i |
| Ah2 | silt | 13–32 | 6.92 c | 97.7 g | 9.50 f | 10.3 e | 187.6 h | ||
| C | silty loam | 32–56 | 7.44 a | 33.2 i | 2.61 h | 12.7 c | 65.1 j |
| No. of Site | Species | Genetic Horizon Acc, to FAO-WRB | Soil Material | Depth (cm) | Macroelements Content (g·kg−1) | ||||
|---|---|---|---|---|---|---|---|---|---|
| P | K | Ca | Mg | Na | |||||
| 1 | C. acutiformis | Ha1 | sapric peat | 0–16 | 0.41 ab | 0.31 b | 14.41 d | 0.62 bc | 0.61 cd |
| Ha2 | sapric peat | 16–31 | 0.32 b | 0.26 bc | 11.20 d | 0.70 b | 0.44 e | ||
| 2 | C. appropinquata | Ha1 | murshic | 0–27 | 0.39 b | 0.08 e | 2.10 f | 0.70 b | 0.80 c |
| Ha2 | murshic | 27–42 | 0.28 bc | 0.19 cd | 1.56 g | 0.44 cd | 0.54 d | ||
| 3 | C. diandra | He1 | hemic peat | 0–20 | 0.32 b | 0.32 b | 23.90 b | 0.62 bc | 0.71 cd |
| He2 | hemic peat | 20–160 | 0.25 bc | 0.13 d | 32.10 a | 0.40 cd | 1.12 b | ||
| 4 | C. disticha | A | sand | 0–32 | 0.05 d | 0.22 c | 0.56 h | 0.44 cd | 0.22 f |
| Bw | silt | 32–72 | 0.06 d | 0.24 c | 0.33 h | 0.28 d | 0.13 g | ||
| 5 | C. flava | Ha1 | sapric peat | 0–15 | 0.51 a | 0.28 bc | 30.10 a | 0.62 bc | 1.02 b |
| Ha2 | sapric peat | 15–33 | 0.56 a | 0.15 d | 5.89 e | 1.01 a | 0.66 cd | ||
| 5 | C. lepidocarpa | Ha1 | sapric peat | 0–15 | 0.51 a | 0.28 bc | 30.10 a | 0.62 bc | 1.02 b |
| Ha2 | sapric peat | 15–33 | 0.56 a | 0.15 d | 5.89 e | 1.01 a | 0.66 cd | ||
| 6 | C. nigra | Ha1 | murshic | 0–31 | 0.35 b | 0.22 c | 33.21 a | 0.54 c | 0.52 de |
| Ha2 | sapric peat | 31–36 | 0.24 bc | 0.38 b | 12.20 d | 0.65 bc | 0.32 e | ||
| 2 | C. paniculata | Ha1 | murshic | 0–27 | 0.39 b | 0.08 e | 2.10 f | 0.70 b | 0.80 c |
| Ha2 | murshic | 27–42 | 0.28 bc | 0.19 cd | 1.56 g | 0.44 cd | 0.54 d | ||
| 7 | C. pseudocyperus | Ha1 | murshic | 0–26 | 0.36 b | 0.14 d | 4.01 ef | 0.51 c | 0.31 e |
| Ha2 | sapric peat | 26–55 | 0.28 bc | 0.09 de | 1.56 g | 0.61 bc | 0.60 cd | ||
| 7 | C. riparia | Ha1 | murshic | 0–26 | 0.36 b | 0.14 d | 4.01 ef | 0.51 c | 0.31 e |
| Ha2 | sapric peat | 26–55 | 0.28 bc | 0.09 de | 1.56 g | 0.61 bc | 0.60 cd | ||
| 8 | C. vulpina | Ah1 | silt | 0–13 | 0.08 d | 0.19 cd | 23.1 b | 1.22 a | 3.67 a |
| Ah2 | silt | 13–32 | 0.18 c | 0.76 s | 5.18 e | 0.66 bc | 0.44 e | ||
| C | silty loam | 32–56 | 0.20 c | 0.29 bc | 20.1 c | 1.02 a | 1.77 b | ||
| No of Site | Species | Genetic Horizon Acc, to FAO-WRB | Soil Material | Depth (cm) | Microelements Content (mg·kg−1) | |||
|---|---|---|---|---|---|---|---|---|
| Cu | Zn | Cr | Ni | |||||
| 1 | C. acutiformis | Ha1 | sapric peat | 0–16 | 12.01 b | 62.33 a | 12.88 e | 11.2 cd |
| Ha2 | sapric peat | 16–31 | 10.1 c | 41.27 c | 15.41 d | 12.4 c | ||
| 2 | C. appropinquata | Ha1 | murshic | 0–27 | 11.2 b | 13.4 f | 9.97 f | 12.31 c |
| Ha2 | murshic | 27–42 | 12.8 b | 12.04 g | 9.5 f | 15.41 b | ||
| 3 | C. diandra | He1 | hemic peat | 0–20 | 11.2 b | 5.63 i | 7.14 h | 10.21 d |
| He2 | hemic peat | 20–160 | 10.21 c | 8.01 h | 6.32 h | 9.41 e | ||
| 4 | C. disticha | A | sand | 0–32 | 5.87 de | 31.77 d | 6.98 h | 6.03 f |
| Bw | silt | 32–72 | 6.21 d | 22.9 e | 9.32 f | 5.04 g | ||
| 5 | C. flava | Ha1 | sapric peat | 0–15 | 9.05 c | 14.21 f | 8.01g h | 18.5 ab |
| Ha2 | sapric peat | 15–33 | 8.87 c | 56.32 ab | 23.14 b | 15.21 b | ||
| 5 | C. lepidocarpa | Ha1 | sapric peat | 0–15 | 9.05 c | 14.21 f | 8.01g h | 18.5 ab |
| Ha2 | sapric peat | 15–33 | 8.87 c | 56.32 ab | 23.14 b | 15.21 b | ||
| 6 | C. nigra | Ha1 | murshic | 0–31 | 17.2 a | 39.82 c | 19.63 bc | 12.7 c |
| Ha2 | sapric peat | 31–36 | 4.01 e | 3.65 ij | 12.01 e | 6.57 f | ||
| 2 | C. paniculata | Ha1 | murshic | 0–27 | 11.2 b | 13.4 f | 9.97 f | 12.31 c |
| Ha2 | murshic | 27–42 | 12.8 b | 12.04 g | 9.5 f | 15.41 b | ||
| 7 | C. pseudocyperus | Ha1 | murshic | 0–26 | 6.23 d | 23.17 e | 27.85 a | 8.07 ef |
| Ha2 | sapric peat | 26–55 | 3.24 e | 2.97 j | 8.96 fg | 9.32 e | ||
| 7 | C. riparia | Ha1 | murshic | 0–26 | 6.23 d | 23.17 e | 27.85 a | 8.07 ef |
| Ha2 | sapric peat | 26–55 | 3.24 e | 2.97 j | 8.96 fg | 9.32 e | ||
| 8 | C. vulpina | Ah1 | silt | 0–13 | 4.12 e | 12.03 g | 15.18 d | 20.1 a |
| Ah2 | silt | 13–32 | 5.21 de | 22.33 e | 17.42 c | 11.4 cd | ||
| C | silty loam | 32–56 | 3.50 e | 21.14 e | 8.31 g | 10.6 d | ||
| Species | C (g kg−1 DM) | N (g kg−1 DM) | C–N | |||
|---|---|---|---|---|---|---|
| Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | |
| C. acutiformis | 434–438 | 436.2 (0.4) G | 12.8–14.7 | 13.81 (0.27) EF | 29.7–34.3 | 31.68 (0.66) D |
| C. appropinquata | 429–433 | 430.9 (0.4) H | 15.0–16.0 | 15.71 (0.10) D | 26.8–28.7 | 27.44 (0.18) E |
| C. diandra | 447–449 | 447.7 (0.3) E | 20.0–21.3 | 20.66 (0.19) B | 21.0–22.4 | 21.68 (0.19) IJ |
| C. disticha | 464–468 | 465.7 (0.4) A | 18.0–18.8 | 18.48 (0.09) C | 24.7–25.9 | 25.20 (0.13) FG |
| C. flava | 444–452 | 447.5 (1.0) E | 11.8–13.9 | 12.59 (0.26) GH | 32.6–37.8 | 35.64 (0.65) BC |
| C. lepidocarpa | 453–458 | 455.3 (0.7) C | 12.5–13.8 | 13.29 (0.15) FG | 33.1–36.6 | 34.30 (0.37) C |
| C. nigra | 459–463 | 461.0 (0.5) B | 12.0–13.0 | 12.53 (0.13) GH | 35.4–38.5 | 36.83 (0.41) B |
| C. paniculata | 453–456 | 455.4 (0.3) C | 19.1–20.5 | 19.93 (0.15) B | 22.2–23.9 | 22.86 (0.18) HI |
| C. pseudocyperus | 435–437 | 435.8 (0.3) G | 9.9–10.6 | 10.13 (0.08) I | 41.0–44.0 | 43.03 (0.35) A |
| C. riparia | 441–444 | 442.6 (0.3) F | 11.0–12.8 | 12.04 (0.19) H | 34.5–40.2 | 36.83 (0.60) B |
| C. vulpina | 440–442 | 440.7 (0.3) F | 12.0–12.9 | 12.43 (0.10) H | 34.1–36.7 | 35.48 (0.28) BC |
| ANOVA P > F | <0.0001 | <0.0001 | <0.0001 | |||
| Species | P (g kg−1 DM) | K (g kg−1 DM) | Ca (g kg−1 DM) | Mg (g kg−1 DM) | Na (g kg−1 DM) | Si (g kg−1 DM) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | |
| Optimal value | 3.0–4.0 | 16.0–20.0 | 6.0–9.0 | 2.0–3.0 | 1.2–2.5 | ca 9.0 | ||||||
| C. acutiformis | 1.54–1.68 | 1.598 (0.019) J | 17.1–18.7 | 18.04 (0.20) H | 7.0–8.1 | 7.75 (0.12) B | 1.00–1.27 | 1.099 (0.038) L | 1.13–1.34 | 1.255 (0.022) I | 7.1–8.2 | 7.87 (0.13) ABC |
| C. appropinquata | 1.87–2.00 | 1.942 (0.016) GH | 15.0–16.7 | 15.75 (0.26) J | 7.4–8.1 | 7.70 (0.11) BC | 2.43–2.63 | 2.560 (0.023) B | 2.29–2.69 | 2.447 (0.046) D | 6.5–7.0 | 6.77 (0.07) EF |
| C. diandra | 1.87–1.93 | 1.900 (0.006) GHI | 19.6–20.5 | 20.10 (0.10) G | 7.1–7.6 | 7.34 (0.06) CD | 2.21–2.67 | 2.489 (0.073) B | 2.20–2.41 | 2.326 (0.027) DE | 7.2–7.7 | 7.46 (0.06) CD |
| C. disticha | 3.12–3.27 | 3.208 (0.015) D | 10.0–11.0 | 10.58(0.14) K | 4.4–4.8 | 4.58 (0.05) I | 3.12–3.24 | 3.171 (0.014) A | 4.33–4.49 | 4.422 (0.018) A | 6.0–7.2 | 6.60 (0.16) EF |
| C. flava | 2.28–2.37 | 2.317 (0.010) F | 23.5–24.9 | 24.22 (0.17) E | 6.4–6.6 | 6.54 (0.02) F | 2.18–2.29 | 2.219 (0.011) C | 1.29–1.35 | 1.313 (0.007) I | 6.5–6.6 | 6.56 (0.02) EF |
| C. lepidocarpa | 3.69–3.90 | 3.821 (0.026) B | 16.3–17.5 | 17.11 (0.11) I | 3.2–3.3 | 3.27 (0.02) J | 2.08–2.24 | 2.151 (0.019) CD | 2.20–2.31 | 2.262 (0.011) EF | 6.5–7.0 | 6.62 (0.06) EF |
| C. nigra | 1.76–1.87 | 1.801 (0.012) HI | 35.2–36.9 | 35.95 (0.16) A | 6.2–6.6 | 6.40 (0.05) FG | 2.49–2.64 | 2.589 (0.015) B | 1.00–1.17 | 1.090 (0.017) J | 6.1–6.6 | 6.48 (0.05) EF |
| C. paniculata | 2.59–2.74 | 2.676 (0.019) E | 22.9–23.3 | 23.12 (0.04) F | 6.0–6.2 | 6.12 (0.03) G | 1.19–1.43 | 1.332 (0.025) JK | 1.24–1.34 | 1.299 (0.011) I | 8.2–8.3 | 8.26 (0.01) A |
| C. pseudocyperus | 1.67–1.84 | 1.751 (0.021) IJ | 17.0–18.1 | 17.39 (0.16) HI | 7.7–8.2 | 7.97 (0.05) B | 2.09–2.56 | 2.259 (0.052) C | 2.03–2.42 | 2.212 (0.049) EF | 6.8–7.1 | 6.95 (0.03) DE |
| C. riparia | 3.00–4.11 | 3.628 (0.116) C | 17.0–18.0 | 17.56 (0.13) HI | 5.1–5.9 | 5.56 (0.12) H | 1.34–1.57 | 1.467 (0.024) HI | 1.59–1.76 | 1.691 (0.015) GH | 6.3–6.9 | 6.59 (0.07) EF |
| C. vulpina | 2.00–2.15 | 2.076 (0.018) G | 28.0–29.1 | 28.48 (0.16) B | 5.3–5.6 | 5.50 (0.03) H | 1.70–1.78 | 1.742 (0.011) EF | 1.57–1.70 | 1.630 (0.015) H | 6.5–7.1 | 6.85 (0.09) E |
| ANOVA P > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||||||
| Optimal Value Species | Cu (mg kg−1 DM) | Zn (mg kg−1 DM) | Mn (mg kg−1 DM) | Fe (mg kg−1 DM) | Cr (mg kg−1 DM) | Ni (mg kg−1 DM) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | Min–Max | Mean (SE) | |||||||
| ca 10.0 | ca 50.0 | ca 50.0 | ca 30.0 | 0.3–5.0 | 0.1–5.0 | |||||||||||||
| C. acutiformis | 5.0–6.0 | 5.36 (0.15) | FG | 26.1–27.3 | 26.84 (0.14) | H | 77.8–78.9 | 78.22 (0.15) | O | 97.6–98.9 | 98.21 (0.16) | M | 1.67–1.88 | 1.773 (0.031) | B | 1.19–1.24 | 1.213 (0.007) | E |
| C. appropinquata | 6.6–6.9 | 6.74 (0.04) | E | 24.7–26.1 | 25.22 (0.18) | IJ | 86.3–87.2 | 86.87 (0.09) | N | 205.3–208.0 | 206.67 (0.37) | F | 0.65–0.69 | 0.667 (0.005) | LM | 0.24–0.28 | 0.252 (0.005) | I |
| C. diandra | 8.4–8.8 | 8.62 (0.05) | D | 38.1–39.1 | 38.51 (0.16) | C | 88.8–90.2 | 89.30 (0.18) | N | 76.0–80.0 | 77.37 (0.47) | N | 0.49–0.53 | 0.505 (0.005) | N | 0.33–0.39 | 0.355 (0.007) | HI |
| C. disticha | 6.3–7.0 | 6.54 (0.10) | E | 25.2–27.1 | 26.49 (0.21) | H | 419.7–424.2 | 421.38 (0.69) | A | 131.0–132.1 | 131.44 (0.15) | K | 0.56–0.66 | 0.609 (0.012) | M | 0.05–0.05 | 0.050 (0.000) | J |
| C. flava | 6.3–6.5 | 6.45 (0.02) | E | 28.9–29.3 | 29.04 (0.05) | FG | 95.0–99.0 | 97.19 (0.47) | M | 95.0–96.8 | 95.83 (0.19) | M | 0.68–0.78 | 0.714 (0.010) | JKL | 2.67–2.88 | 2.776 (0.025) | B |
| C. lepidocarpa | 8.6–8.9 | 8.77 (0.04) | D | 32.8–33.6 | 33.08 (0.09) | E | 100.2–103.9 | 102.14 (0.50) | L | 143.0–147.0 | 145.13 (0.52) | J | 0.76–0.81 | 0.786 (0.005) | JK | 0.40–0.49 | 0.444 (0.010) | H |
| C. nigra | 10.0–10.6 | 10.25 (0.08) | B | 25.2–26.0 | 25.72 (0.10) | HI | 181.2–185.0 | 183.12 (0.52) | C | 339.6–347.0 | 343.89 (1.07) | C | 0.77–0.83 | 0.796 (0.006) | J | 0.83–0.87 | 0.847 (0.005) | F |
| C. paniculata | 2.2–4.3 | 3.99 (0.23) | H | 27.5–29.1 | 28.25 (0.22) | G | 72.9–75.1 | 74.19 (0.25) | P | 151.2–156.5 | 153.65 (0.69) | I | 1.49–1.55 | 1.526 (0.006) | D | 0.20–0.25 | 0.226 (0.006) | IJ |
| C.pseudocyperus | 10.0–11.0 | 10.37 (0.14) | B | 34.0–35.1 | 34.29 (0.13) | D | 170.0–177.9 | 174.37 (1.12) | E | 422.0–437.1 | 429.32 (2.12) | B | 1.98–2.31 | 2.090 (0.038) | A | 1.49–1.69 | 1.558 (0.021) | D |
| C. riparia | 5.0–5.4 | 5.16 (0.06) | G | 27.1–29.0 | 28.13 (0.23) | G | 95.9–100.1 | 98.24 (0.59) | M | 118.9–121.0 | 119.61 (0.27) | L | 1.67–1.88 | 1.742 (0.022) | B | 1.76–2.01 | 1.890 (0.030) | C |
| C. vulpina | 12.0–13.0 | 12.44 (0.13) | A | 41.8–42.9 | 42.24 (0.15) | B | 164.0–167.5 | 166.05 (0.51) | F | 193.7–198.0 | 195.52 (0.57) | G | 0.68–0.77 | 0.707 (0.008) | KL | 3.25–3.56 | 3.376 (0.036) | A |
| ANOVA P > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||||||||||||
| Optimal Value Species | K–Na | K–Ca + Mg | Ca–P |
|---|---|---|---|
| C. acutiformis | 14:1 | 2.0 | 4.8:1 |
| C. appropinquata | 6:1 | 1.5 | 3.9:1 |
| C. diandra | 9:1 | 2.0 | 3.8:1 |
| C. disticha | 2:1 | 1.4 | 1.4:1 |
| C. flava | 19:1 | 3.1 | 2.8:1 |
| C. lepidocarpa | 7:1 | 3.1 | 0.86:1 |
| C. nigra | 32:1 | 4.2 | 3.6:1 |
| C. paniculata | 18:1 | 3.1 | 2.3:1 |
| C. pseudocyperus | 8:1 | 1.7 | 4.4:1 |
| C. riparia | 10:1 | 2.5 | 1.6:1 |
| C. vulpina | 18:1 | 3.9 | 2.6:1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janyszek-Sołtysiak, M.; Grzelak, M.; Gajewski, P.; Jagodziński, A.M.; Gaweł, E.; Wrońska-Pilarek, D. Mineral Contents in Aboveground Biomass of Sedges (Carex L., Cyperaceae). Energies 2021, 14, 8007. https://doi.org/10.3390/en14238007
Janyszek-Sołtysiak M, Grzelak M, Gajewski P, Jagodziński AM, Gaweł E, Wrońska-Pilarek D. Mineral Contents in Aboveground Biomass of Sedges (Carex L., Cyperaceae). Energies. 2021; 14(23):8007. https://doi.org/10.3390/en14238007
Chicago/Turabian StyleJanyszek-Sołtysiak, Magdalena, Mieczysław Grzelak, Piotr Gajewski, Andrzej M. Jagodziński, Eliza Gaweł, and Dorota Wrońska-Pilarek. 2021. "Mineral Contents in Aboveground Biomass of Sedges (Carex L., Cyperaceae)" Energies 14, no. 23: 8007. https://doi.org/10.3390/en14238007
APA StyleJanyszek-Sołtysiak, M., Grzelak, M., Gajewski, P., Jagodziński, A. M., Gaweł, E., & Wrońska-Pilarek, D. (2021). Mineral Contents in Aboveground Biomass of Sedges (Carex L., Cyperaceae). Energies, 14(23), 8007. https://doi.org/10.3390/en14238007