Using Fluorescence Quenching Titration to Determine the Orientation of a Model Transmembrane Protein in Mimic Membranes

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Vesicle Preparation
2.2. DLS Measurements
2.3. TEM Imaging
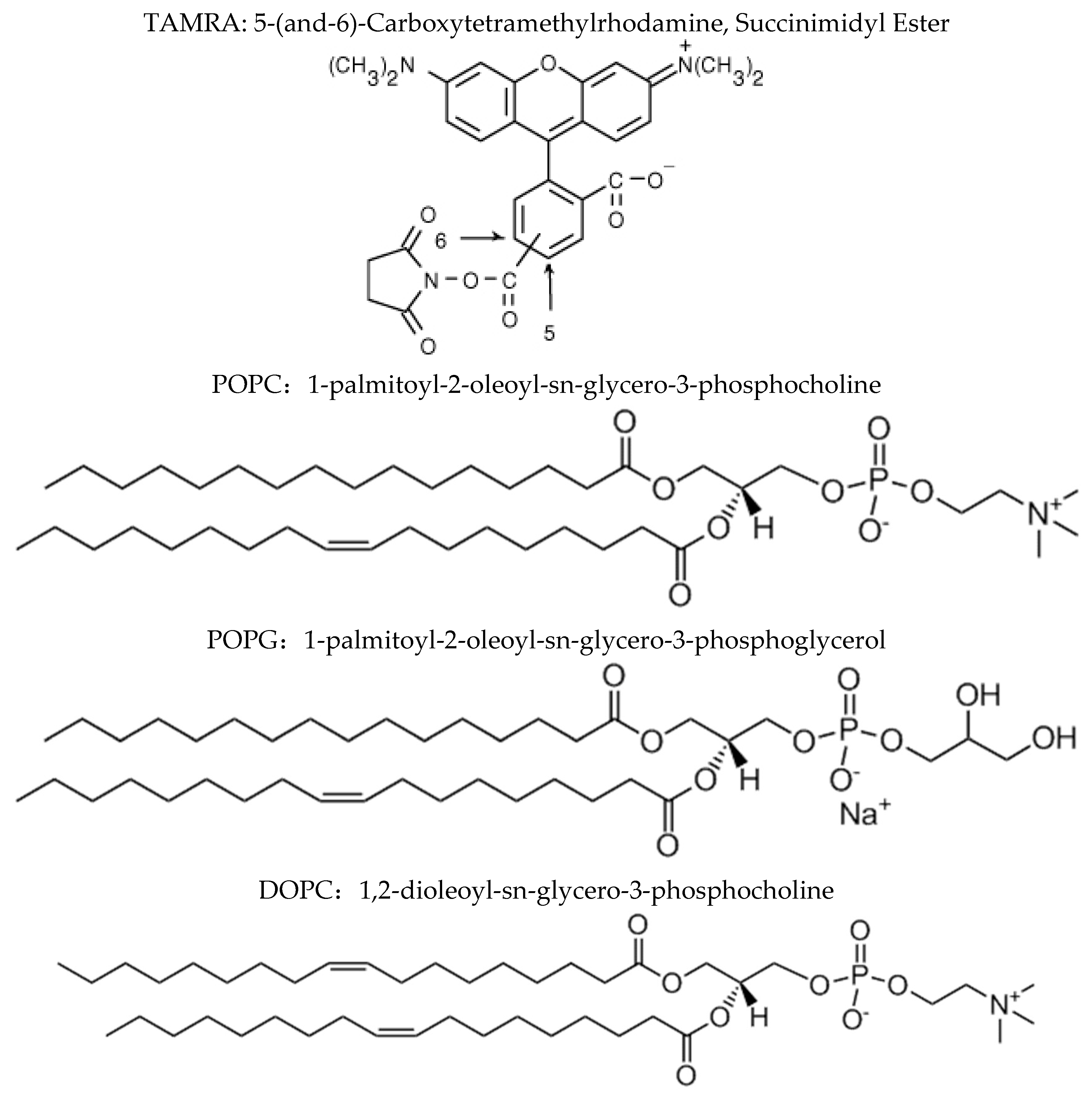
2.4. Preparation of bR-TAMRA and Reconstitution into Vesicles
2.5. Fluorescence Quenching Titration Experiments
3. Results and Discussion
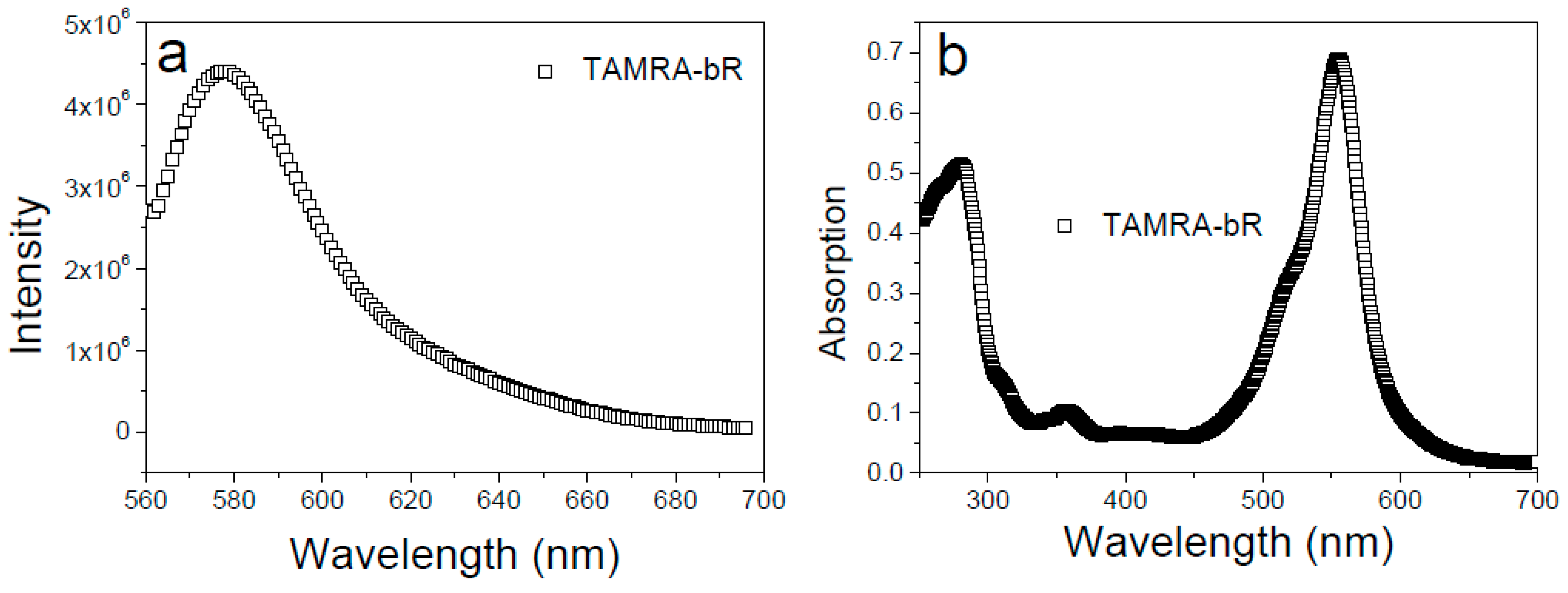
3.1. Labeling of bR with TAMRA and Reconstitution into Vesicles
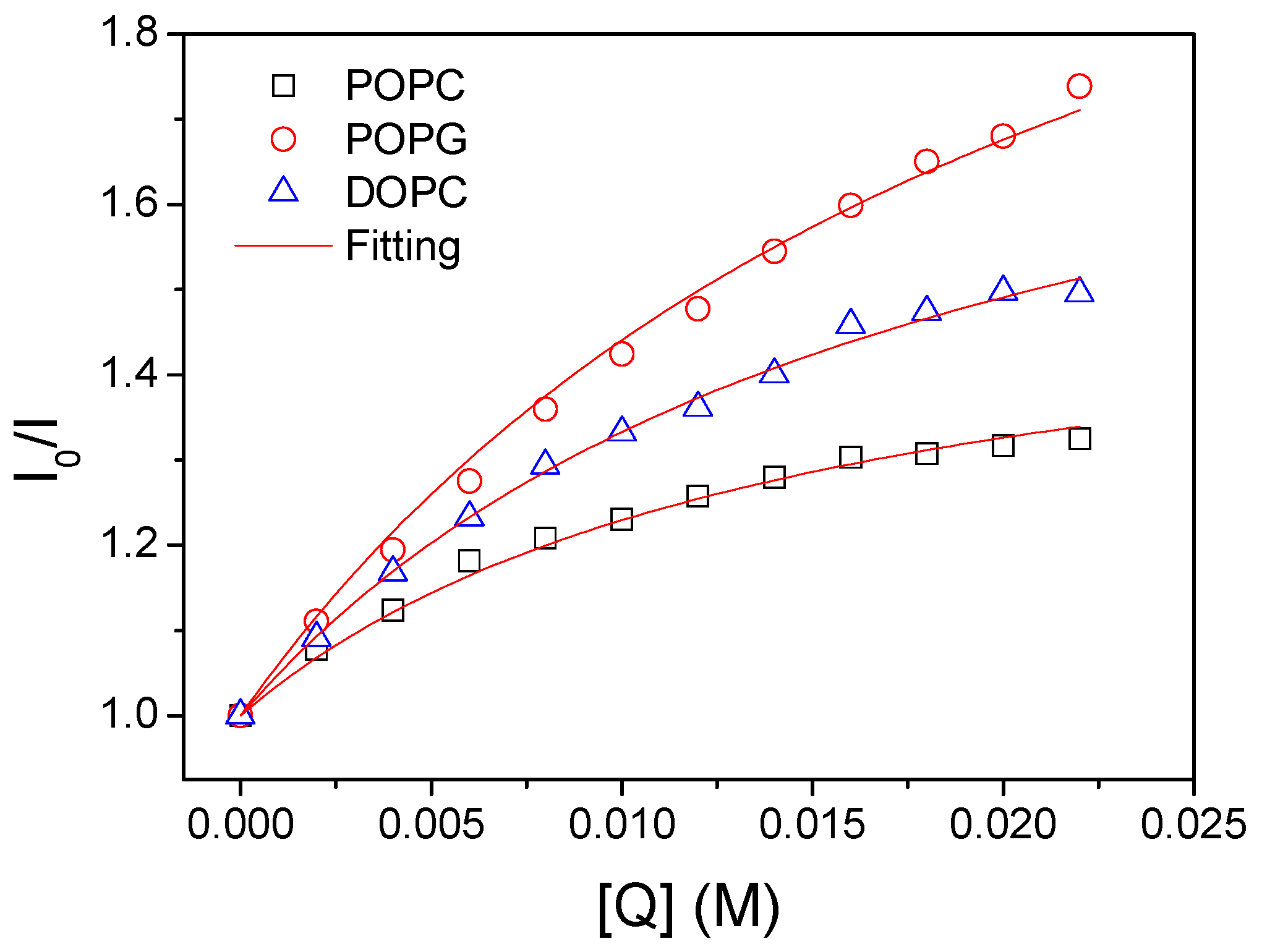
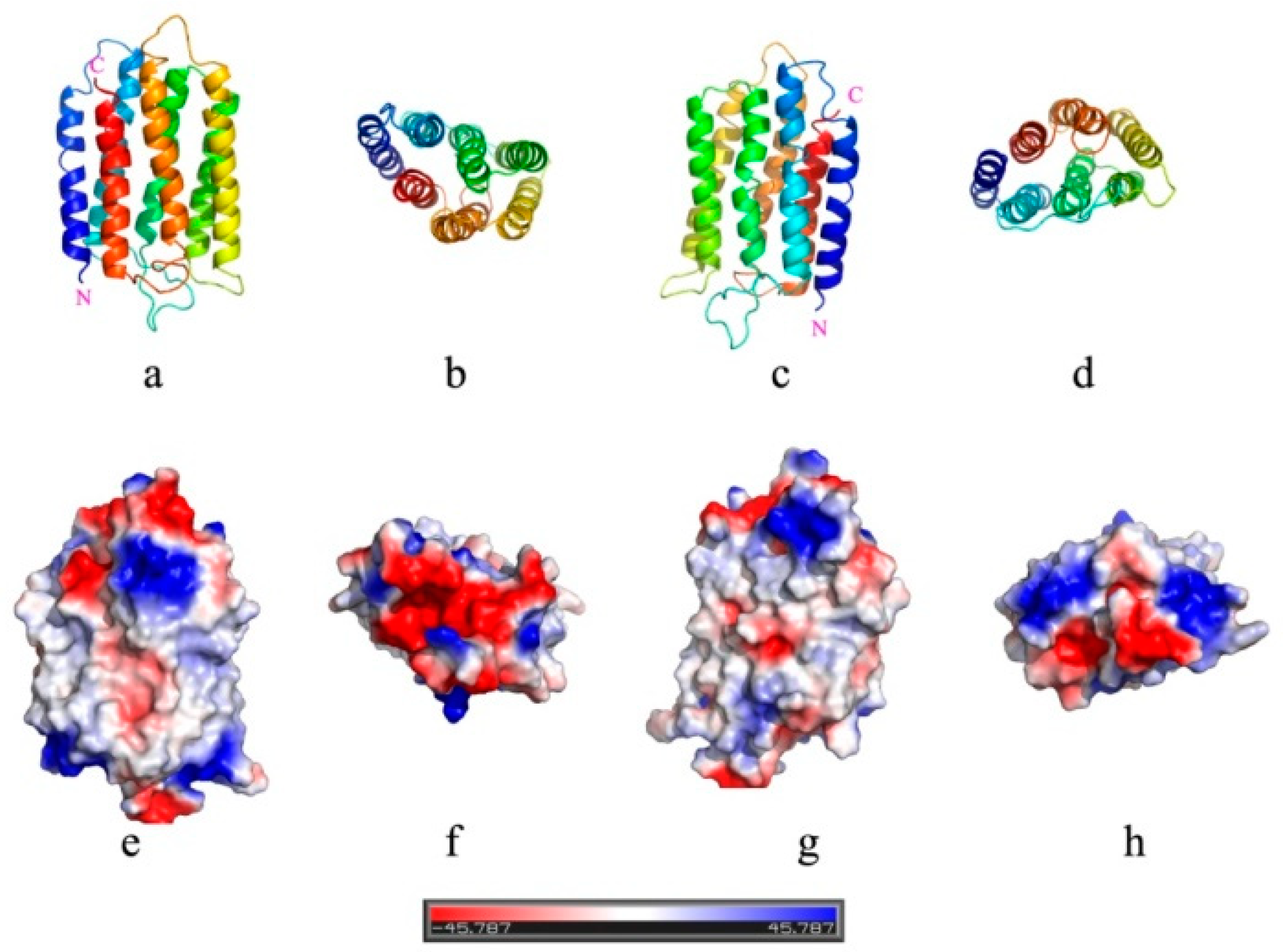
3.2. Lipid Composition Modulates Orientation of bR in Mimic Membranes
3.3. Effects of Other Environmental Factors
3.4. Folding States of bR Being Inserted into Vesicles
3.5. Caveats and Drawbacks
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dill, K.A. Theory for the folding and stability of globular proteins. Biochemistry 1985, 24, 1501–1509. [Google Scholar] [CrossRef]
- Bowie, J.U. Solving the membrane protein folding problem. Nature 2005, 438, 581. [Google Scholar] [CrossRef]
- White, S.H.; Wimley, W.C. Membrane protein folding and stability: Physical principles. Annu. Rev. Biophys. Biomol. Struct. 1999, 28, 319–365. [Google Scholar] [CrossRef]
- Jefferson, R.E.; Min, D.; Corin, K.; Wang, J.Y.; Bowie, J.U. Applications of Single-Molecule Methods to Membrane Protein Folding Studies. J. Mol. Biol. 2018, 430, 424–437. [Google Scholar] [CrossRef]
- Krainer, G.; Hartmann, A.; Anandamurugan, A.; Gracia, P.; Keller, S.; Schlierf, M. Ultrafast Protein Folding in Membrane-Mimetic Environments. J. Mol. Biol. 2018, 430, 554–564. [Google Scholar] [CrossRef]
- Henderson, R.; Unwin, P.N. Three-dimensional model of purple membrane obtained by electron microscopy. Nature 1975, 257, 28–32. [Google Scholar] [CrossRef]
- Stefanovic, S.; Hegde, R.S. Identification of a targeting factor for posttranslational membrane protein insertion into the ER. Cell 2007, 128, 1147–1159. [Google Scholar] [CrossRef]
- Hessa, T.; Kim, H.; Bihlmaier, K.; Lundin, C. Recognition of transmembrane helices by the endoplasmic reticulum translocon. Nature 2005, 433, 377. [Google Scholar] [CrossRef]
- Hessa, T.; Meindl-Beinker, N.M.; Bernsel, A.; Kim, H.; Sato, Y.; Lerch-Bader, M.; Nilsson, I.; White, S.H.; Von Heijne, G. Molecular code for transmembrane-helix recognition by the Sec61 translocon. Nature 2007, 450, 1026. [Google Scholar] [CrossRef]
- Hardie, K.R.; Lory, S.; Pugsley, A. Insertion of an outer membrane protein in Escherichia coli requires a chaperone-like protein. EMBO J. 1996, 15, 978. [Google Scholar] [CrossRef]
- Bulieris, P.V.; Behrens, S.; Holst, O.; Kleinschmidt, J.H. Folding and insertion of the outer membrane protein OmpA is assisted by the chaperone Skp and by lipopolysaccharide. J. Biol. Chem. 2003, 278, 9092–9099. [Google Scholar] [CrossRef]
- Booth, P.J.; Curran, A.R. Membrane protein folding. Curr. Opin. Struct. Biol. 1999, 9, 115–121. [Google Scholar] [CrossRef]
- Booth, P.J.; Riley, M.L.; Flitsch, S.L.; Templer, R.H.; Farooq, A.; Curran, A.R.; Chadborn, N.; Wright, P. Evidence that bilayer bending rigidity affects membrane protein folding. Biochemistry 1997, 36, 197–203. [Google Scholar] [CrossRef]
- Shen, Y.; Safinya, C.R.; Liang, K.S.; Ruppert, A.; Rothschild, K.J. Stabilization of the membrane protein bacteriorhodopsin to 140 C in two-dimensional films. Nature 1993, 366, 48–50. [Google Scholar] [CrossRef]
- Curran, A.R.; Templer, R.H.; Booth, P.J. Modulation of folding and assembly of the membrane protein bacteriorhodopsin by intermolecular forces within the lipid bilayer. Biochemistry 1999, 38, 9328–9336. [Google Scholar] [CrossRef]
- Booth, P.J.; Flitsch, S.L.; Stern, L.J.; Greenhalgh, D.A.; Kim, P.S.; Khorana, H.G. Intermediates in the folding of the membrane protein bacteriorhodopsin. Nat. Struct. Mol. Biol. 1995, 2, 139–143. [Google Scholar] [CrossRef]
- Booth, P.J. Unravelling the folding of bacteriorhodopsin. BBA-Bioenergetics 2000, 1460, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Bogdanov, M.; Dowhan, W. Lipid-assisted protein folding. J. Biol. Chem. 1999, 274, 36827–36830. [Google Scholar] [CrossRef]
- Blom, T.; Ikonen, E. Lipid-protein interactions: Finding the perfect match. Curr. Opin. Lipidol. 2012, 23, 581–583. [Google Scholar] [CrossRef]
- Bogdanov, M.; Xie, J.; Dowhan, W. Lipid-protein interactions drive membrane protein topogenesis in accordance with the positive inside rule. J. Biol. Chem. 2009, 284, 9637–9641. [Google Scholar] [CrossRef]
- Lee, D.; Walter, K.F.; Bruckner, A.K.; Hilty, C.; Becker, S.; Griesinger, C. Bilayer in small bicelles revealed by lipid-protein interactions using NMR spectroscopy. J. Am. Chem. Soc. 2008, 130, 13822–13823. [Google Scholar] [CrossRef]
- Yue, T.; Sun, M.; Zhang, S.; Ren, H.; Ge, B.; Huang, F. How transmembrane peptides insert and orientate in biomembranes: A combined experimental and simulation study. Phys. Chem. Chem. Phys. 2016, 18, 17483–17494. [Google Scholar] [CrossRef]
- Lyttle, M.H.; Carter, T.G.; Dick, D.J.; Cook, R.M. A tetramethyl rhodamine (Tamra) phosphoramidite facilitates solid-phase-supported synthesis of 5’-Tamra DNA. J. Org. Chem. 2000, 65, 9033–9038. [Google Scholar] [CrossRef]
- Lee, G.H.; Lee, E.J.; Hah, S.S. TAMRA- and Cy5-labeled probe for efficient kinetic characterization of caspase-3. Anal. Biochem. 2014, 446, 22–24. [Google Scholar] [CrossRef]
- Oesterhelt, D.; Stoeckenius, W. Isolation of the cell membrane of Halobacterium halobium and its fractionation into red and purple membrane. Method. Enzymol. 1974, 31, 667–678. [Google Scholar]
- Gaudriault, G.; Vincent, J.-P. Selective labeling of α-or ϵ-amino groups in peptides by the Bolton-Hunter reagent. Peptides 1992, 13, 1187–1192. [Google Scholar] [CrossRef]
- Xu, T.; Park, S.K.; Venable, J.D.; Wohlschlegel, J.A.; Diedrich, J.K.; Cociorva, D.; Lu, B.; Liao, L.; Hewel, J.; Han, X.; et al. ProLuCID: An improved SEQUEST-like algorithm with enhanced sensitivity and specificity. J. Proteomics 2015, 129, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Miteva, M.A.; Tufféry, P.; Villoutreix, B.O. PCE: Web tools to compute protein continuum electrostatics. Nucleic Acids Res. 2005, 33, W372–W375. [Google Scholar] [CrossRef]
- Grigorieff, N.; Ceska, T.A.; Downing, K.H.; Baldwin, J.M.; Henderson, R. Electron-crystallographic refinement of the structure of bacteriorhodopsin. J. Mol. Biol. 1996, 259, 393–421. [Google Scholar] [CrossRef]
- Subramaniam, S. The structure of bacteriorhodopsin: An emerging consensus. Curr. Opin. Struct. Biol. 1999, 9, 462–468. [Google Scholar] [CrossRef]
- Xia, Y.; Jang, H.-S.; Shen, Z.; Bothun, G.D.; Li, Y.; Nieh, M.-P. Effects of Membrane Defects and Polymer Hydrophobicity on Networking Kinetics of Vesicles. Langmuir 2017, 33, 5745–5751. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Wang, X.; Li, J.; Ren, H.; Huang, F. Folding of newly translated membrane protein CCR5 is assisted by the chaperonin GroEL-GroES. Sci. Rep. 2015, 5, 17037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popot, J.L.; Gerchman, S.E.; Engelman, D.M. Refolding of bacteriorhodopsin in lipid bilayers. A thermodynamically controlled two-stage process. J. Mol. Biol. 1987, 198, 655–676. [Google Scholar] [CrossRef]
- Huang, K.S.; Bayley, H.; Liao, M.J.; London, E.; Khorana, H.G. Refolding of an integral membrane protein. Denaturation, renaturation and reconstitution of intact bacteriorhodopsin and two proteolytic fragments. J. Biol. Chem. 1981, 256, 3802–3809. [Google Scholar] [PubMed]
- Wang, J.; Heyes, C.D.; El-Sayed, M.A. Refolding of Thermally Denatured Bacteriorhodopsin in Purple Membrane. J. Phys. Chem. B 2001, 106, 723–729. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ionic Strength | x |
|---|---|
| POPC 10 mM | 0.64 ± 0.01 |
| POPC 50 mM | 0.66 ± 0.03 |
| POPG 10 mM | 0.41 ± 0.01 |
| POPG 50 mM | 0.49 ± 0.01 |
| pH | x |
|---|---|
| POPC pH 6.2 | 0.64 ± 0.03 |
| POPC pH 8.0 | 0.57 ± 0.02 |
| POPG pH 6.2 | 0.48 ± 0.01 |
| POPG pH 8.0 | 0.44 ± 0.01 |
| Lipid Vesicle Size | x |
|---|---|
| POPC 50 nm | 0.64 ± 0.01 |
| POPC 100 nm | 0.64 ± 0.01 |
| POPG 50 nm | 0.43 ± 0.01 |
| POPG 100 nm | 0.40 ± 0.01 |
| DOPC 50 nm | 0.57 ± 0.01 |
| DOPC 100 nm | 0.53 ± 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Ge, B.; Zhang, S.; Li, J.; Sun, C.; Yue, T.; Huang, F. Using Fluorescence Quenching Titration to Determine the Orientation of a Model Transmembrane Protein in Mimic Membranes. Materials 2019, 12, 349. https://doi.org/10.3390/ma12030349
Huang H, Ge B, Zhang S, Li J, Sun C, Yue T, Huang F. Using Fluorescence Quenching Titration to Determine the Orientation of a Model Transmembrane Protein in Mimic Membranes. Materials. 2019; 12(3):349. https://doi.org/10.3390/ma12030349
Chicago/Turabian StyleHuang, Haihong, Baosheng Ge, Shuai Zhang, Jiqiang Li, Chenghao Sun, Tongtao Yue, and Fang Huang. 2019. "Using Fluorescence Quenching Titration to Determine the Orientation of a Model Transmembrane Protein in Mimic Membranes" Materials 12, no. 3: 349. https://doi.org/10.3390/ma12030349