The Influence of Different Classes of Amino Acids on Calcium Phosphates Seeded Growth


 , ,
, ,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Precipitation Experiments
2.2.1. Preparation of Stock Solutions
2.2.2. Preparation of Metastable Solutions
2.2.3. Precipitation Systems
2.3. Precipitate Characterization
3. Results and Discussion
3.1. Influence of Amino Acids on the Rate of Precipitation
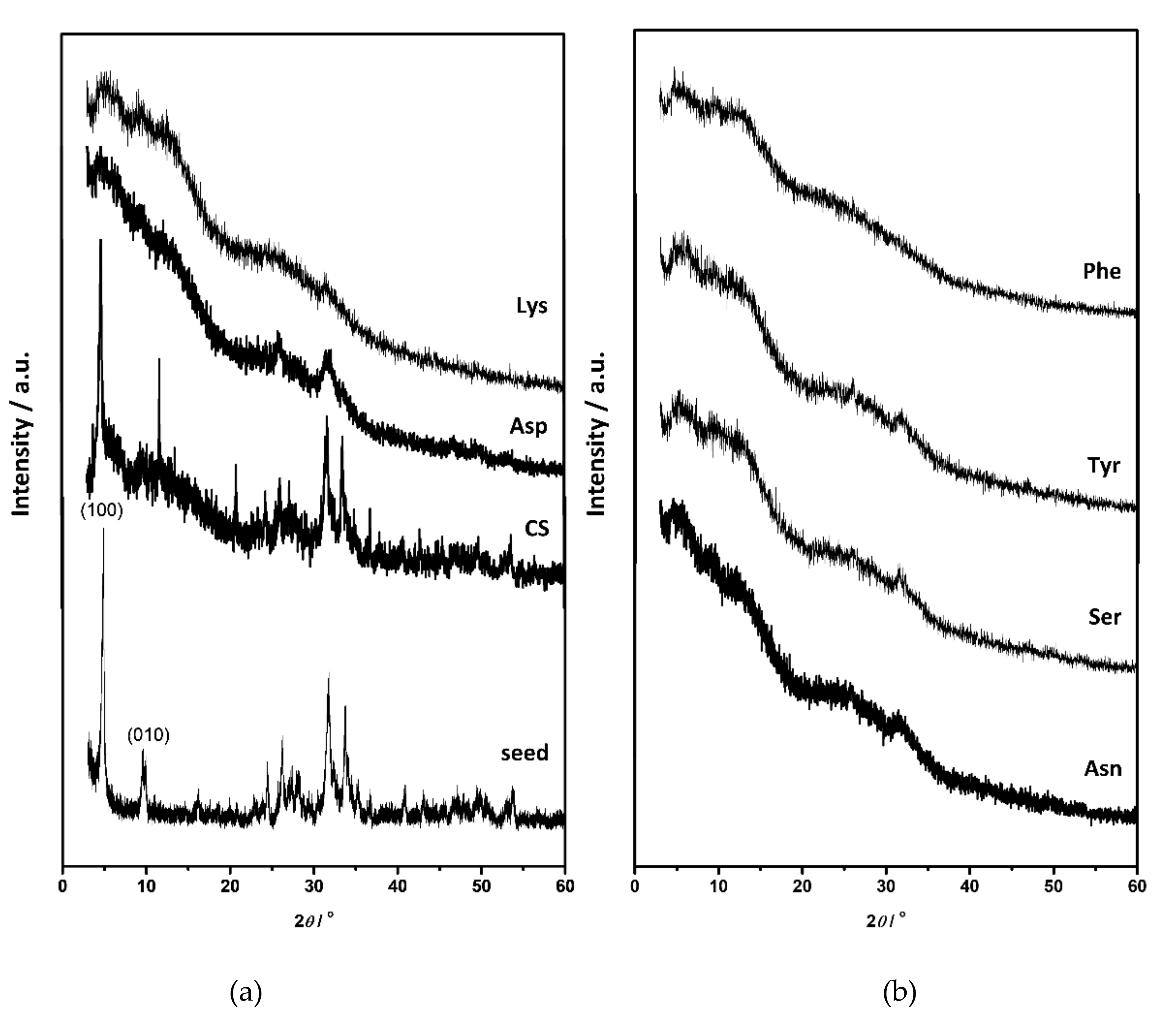
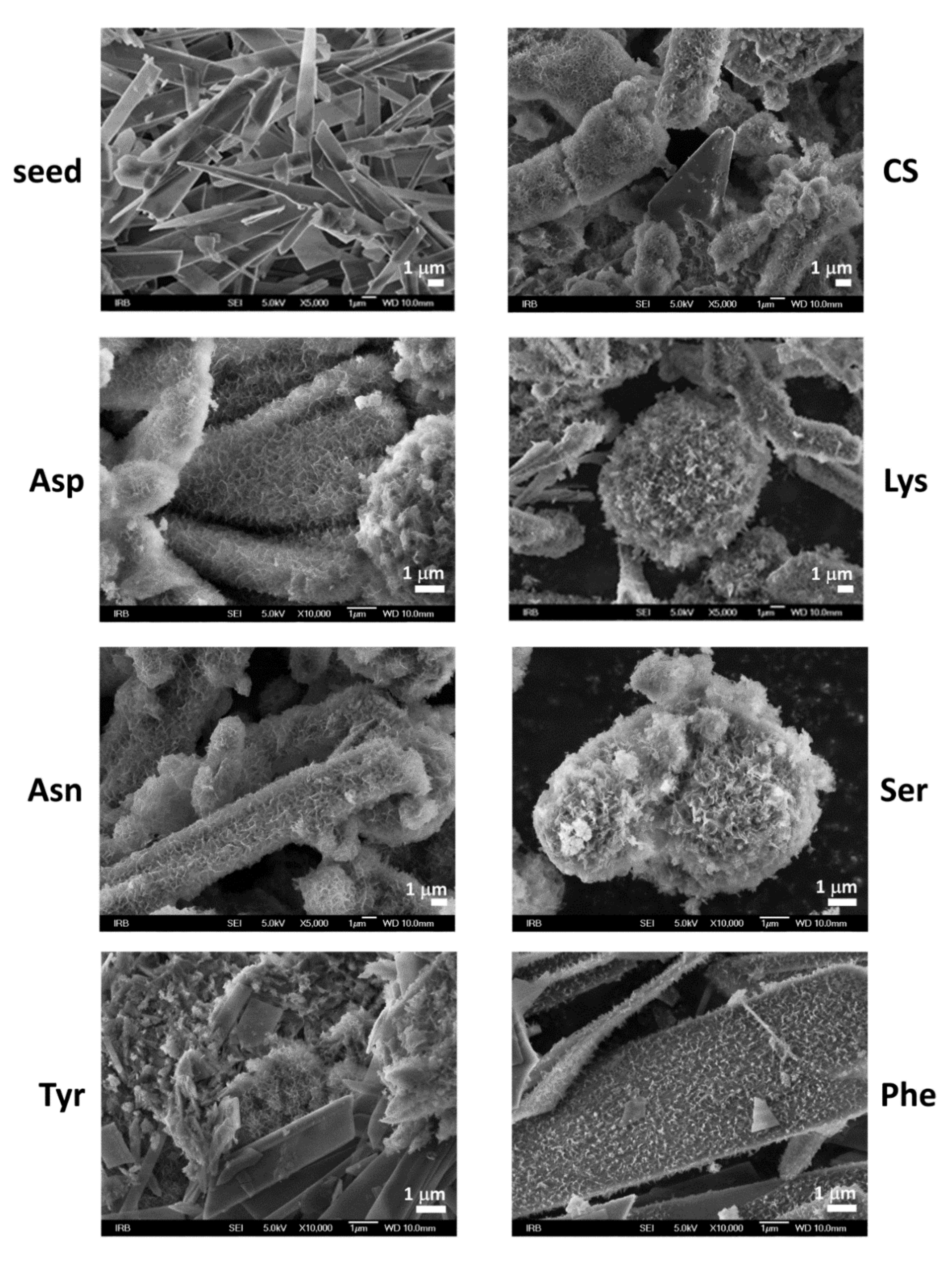
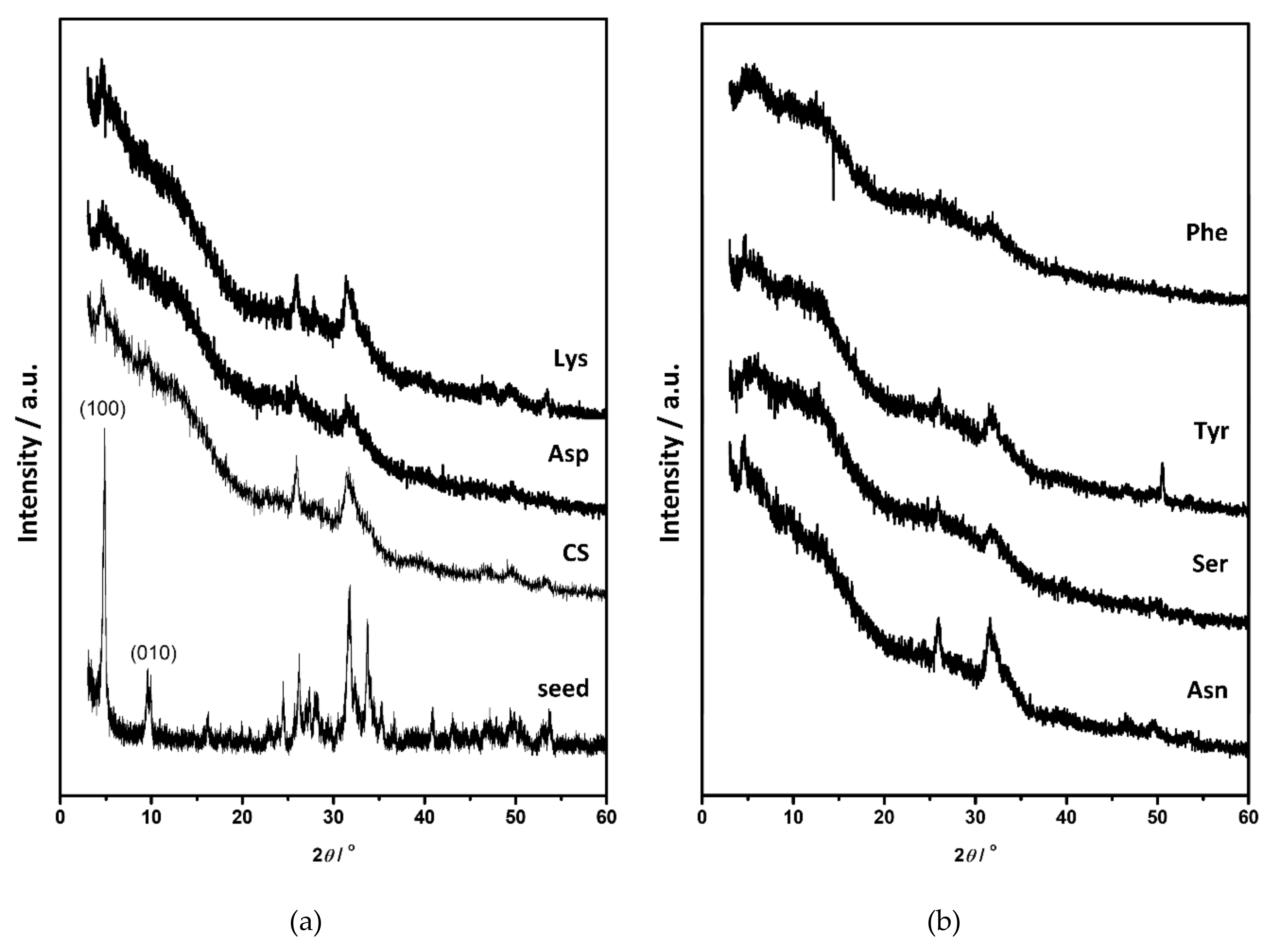
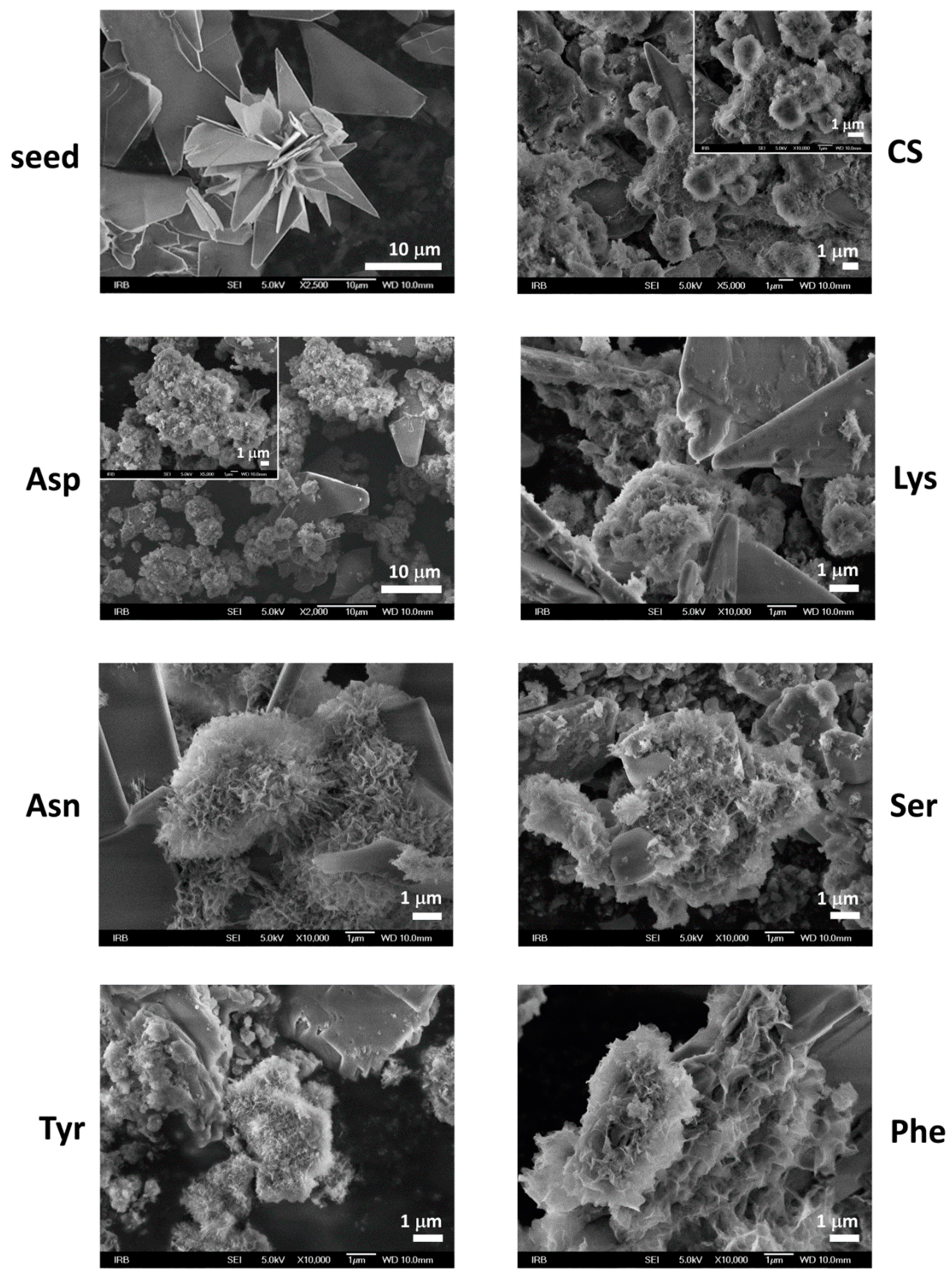
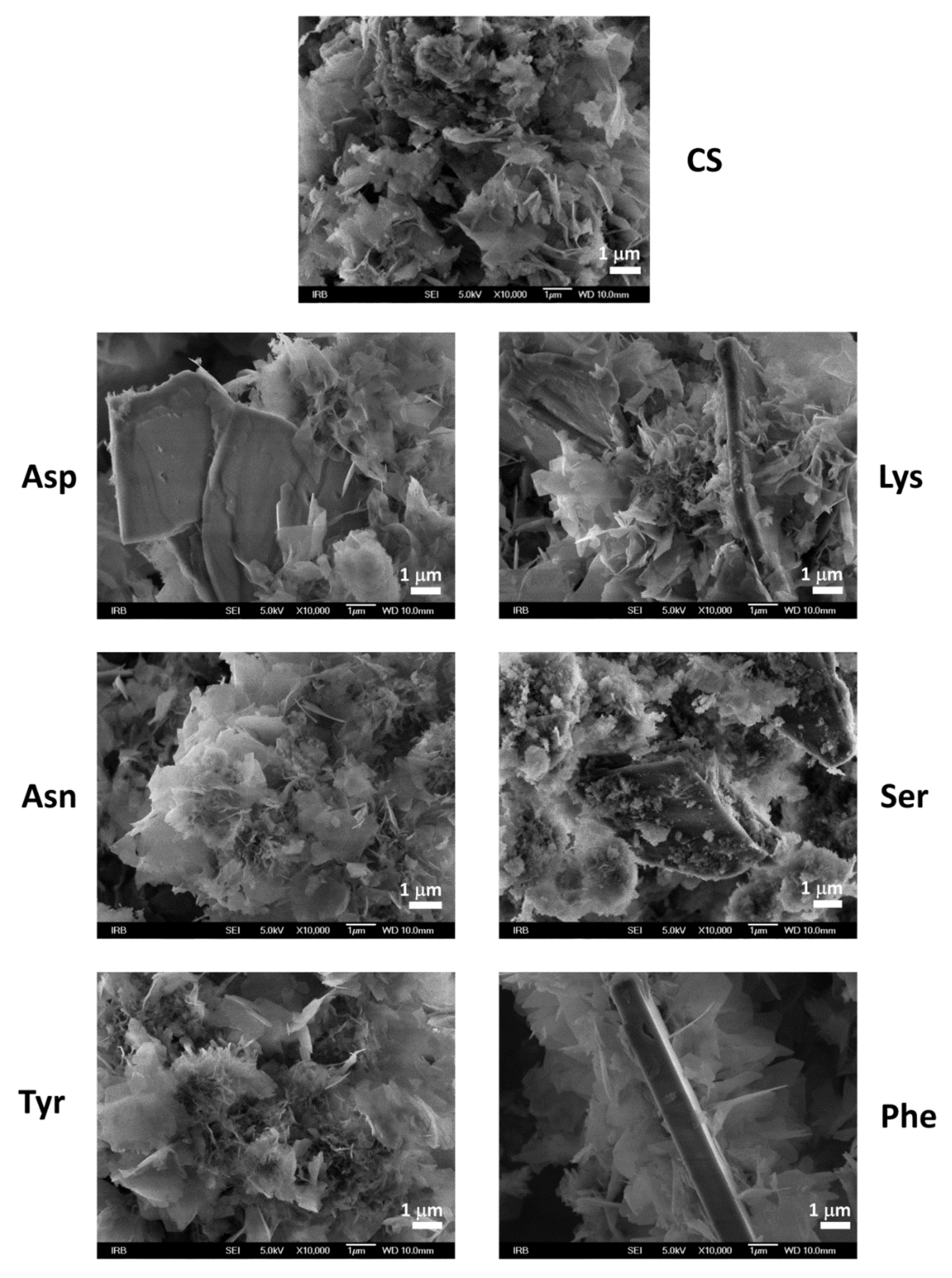
3.2. Influence on Composition and Morphology
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press: New York, NY, USA, 1989. [Google Scholar]
- Mann, S. Biomineralization: Principles and Concepts in Bioinorganic Materials Chemistry; Oxford University Press: New York, NY, USA, 2001; p. 198. [Google Scholar]
- LeGeros, R.Z. Calcium Phosphate-Based Osteoinductive Materials. Chem. Rev. 2008, 108, 4742–4753. [Google Scholar] [CrossRef]
- Crichton, R.R. Biological Inorganic Chemistry; Elsevier: Amsterdam, The Netherlands, 2008; p. 369. [Google Scholar]
- Weiner, S.; Dove, P.M. An Overview of Biomineralization Processes and the Problem of the Vital Effect. Rev. Mineral. Geochem. 2003, 54, 1–29. [Google Scholar] [CrossRef]
- Addadi, L.; Joester, D.; Nudelman, F.; Weiner, S. Mollusk shell formation: A source of new concepts for understanding biomineralization processes. Chem. Eur. J. 2006, 12, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, M.B.; Sandhage, K.H.; Naik, R.R. Protein- and peptide-directed syntheses of inorganic materials. Chem. Rev. 2008, 108, 4935–4978. [Google Scholar] [CrossRef] [PubMed]
- Falini, G.; Fermani, S. The strategic role of adsorption phenomena in biomineralization. Cryst. Res. Technol. 2013, 48, 864–876. [Google Scholar] [CrossRef]
- Dorozhkin, S.V. Calcium Orthophosphates: Applications in Nature, Biology, and Medicine; Pan Stanford Publishing Pte. Ltd.: Singapore, 2012; ISBN 9789814316620. [Google Scholar]
- Dorozhkin, S.V.; Epple, M. Biological and medical significance of calcium phosphates. Angew. Chem. Int. Ed. 2002, 41, 3130–3146. [Google Scholar] [CrossRef]
- Tung, M.S. Calcium Phosphates: Structure, Composition, Solubility, and Stability. In Calcium Phosphates in Biological and Industrial Systems; Amjad, Z., Ed.; Springer: New York, NY, USA, 1998; pp. 1–19. [Google Scholar]
- Zhang, Z.L.; Chen, X.R.; Bian, S.; Huang, J.; Zhang, T.L.; Wang, K. Identification of dicalcium phosphate dihydrate deposited during osteoblast mineralization in vitro. J. Inorg. Biochem. 2014, 131, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.; Brown, P.W. Reactions of octacalcium phosphate to form hydroxyapatite. J. Cryst. Growth 1996, 165, 106–115. [Google Scholar] [CrossRef]
- Zhang, J.; Nancollas, G.H. Kinetics and mechanisms of octacalcium phosphate dissolution at 37 °C. J. Phys. Chem. 1992, 96, 5478–5483. [Google Scholar] [CrossRef]
- Arellano-Jiménez, M.J.; García-García, R.; Reyes-Gasga, J. Synthesis and hydrolysis of octacalcium phosphate and its characterization by electron microscopy and X-ray diffraction. J. Phys. Chem. Solids 2009, 70, 390–395. [Google Scholar] [CrossRef]
- Brown, P.W.; Fulmer, M. Kinetics of Hydroxyapatite Formation at Low Temperature. J. Am. Ceram. Soc. 1991, 74, 934–940. [Google Scholar] [CrossRef]
- Bodier-Houllé, P.; Steuer, P.; Meyer, J.M.; Bigeard, L.; Cuisinier, F.J.G. High-resolution electron-microscopic study of the relationship between human enamel and dentin crystals at the dentinoenamel junction. Cell Tissue Res. 2000, 301, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Xin, R.; Leng, Y.; Wang, N. In situ TEM examinations of octacalcium phosphate to hydroxyapatite transformation. J. Cryst. Growth 2006, 289, 339–344. [Google Scholar] [CrossRef]
- Chu, X.; Jiang, W.; Zhang, Z.; Yan, Y.; Pan, H.; Xu, X.; Tang, R. Unique Roles of Acidic Amino Acids in Phase Transformation of Calcium Phosphates. J. Phys. Chem. B 2011, 115, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Saruwatari, K.; Kogure, T.; Yamamoto, Y.; Nishimura, T.; Kato, T.; Nagasawa, H. An Acidic Matrix Protein, Pif, Is a Key Macromolecule for Nacre Formation. Science 2009, 325, 1388–1390. [Google Scholar] [CrossRef]
- Jahromi, M.T.; Yao, G.; Cerruti, M. The importance of amino acid interactions in the crystallization of hydroxyapatite. J. R. Soc. Interface 2012, 10, 20120906. [Google Scholar] [CrossRef] [PubMed]
- Tavafoghi, M.; Cerruti, M. The role of amino acids in hydroxyapatite mineralization. J. R. Soc. Interface 2016, 13, 20160462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubini, K.; Boanini, E.; Bigi, A. Role of aspartic and polyaspartic acid on the synthesis and hydrolysis of brushite. J. Funct. Biomater. 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-McQuire, R.; Chane-Ching, J.Y.; Vignaud, E.; Lebugle, A.; Mann, S. Synthesis and characterization of amino acid-functionalized hydroxyapatite nanorods. J. Mater. Chem. 2004, 14, 2277–2281. [Google Scholar] [CrossRef]
- Boanini, E.; Torricelli, P.; Gazanno, M.; Giardino, R.; Bigi, A. Nanocomposites of hydroxyapatite with aspartic acid and glutamic acid and their interaction with osteoblast-like cells. Biomaterials 2006, 27, 4428–4433. [Google Scholar] [CrossRef]
- Palazzo, B.; Walsh, D.; Iafisco, M.; Foresti, E.; Bertinetti, L.; Martra, G.; Bianchi, C.L.; Cappelletti, G.; Roveri, N. Amino acid synergetic effect on structure, morphology and surface properties of biomimetic apatite nanocrystals. Acta Biomater. 2009, 5, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Jack, K.S.; Vizcarra, T.G.; Trau, M. Characterization and Surface Properties of Amino-Acid-Modified Carbonate-Containing Hydroxyapatite Particles. Langmuir 2007, 23, 12233–12242. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, B.; Wang, L.; Qin, Y.; Henneman, Z.J.; Nancollas, G.H. Influence of magnesium ions and amino acids on the nucleation and growth of hydroxyapatite. Cryst. Eng. Comm. 2011, 13, 1153–1158. [Google Scholar] [CrossRef]
- Koutsoupolos, S.; Dalas, E. Hydroxyapatite Crystallization in the Presence of Amino Acids with Uncharged Polar Side Groups: Glycine, Cysteine, Cystine, and Glutamine. Langmuir 2001, 17, 1074–1079. [Google Scholar] [CrossRef]
- Tsai, T.W.T.; Chen, W.Y.; Tseng, Y.H.; Chan, J.C.C. Phase transformation of calcium phosphates in the presence of glutamic acid. Can. J. Chem. 2011, 89, 885–891. [Google Scholar] [CrossRef]
- Sikirić, M.; Babić-Ivančić, V.; Milat, O.; Sarig, S.; Füredi-Milhofer, H. Factors Influencing Additive Interactions with Calcium Hydrogenphosphate Dihydrate Crystals. Langmuir 2000, 16, 9261–9266. [Google Scholar] [CrossRef]
- Gilman, H.; Hukins, D.W.L. Seeded growth of hydroxyapatite in the presence of dissolved albumin. J. Inorg. Biochem. 1994, 55, 21–30. [Google Scholar] [CrossRef]
- Weiner, S.; Wagner, H.D. The Material Bone: Structure-Mechanical Function Relations. Ann. Rev. Mater. Sci. 1998, 28, 271–298. [Google Scholar] [CrossRef]
- Abou Neel, E.; Aljabo, A.; Strange, A.; Ibrahim, S.; Coathup, M.; Young, A.; Bozec, L.; Mudera, V. Demineralization & remineralization dynamics in teeth and bone. Inter. J. Nanomed. 2016, 11, 4743–4763. [Google Scholar]
- Elliott, J.C. Structure and chemistry of the apatites and other calcium orthophosphates. Stud. Org. Chem. 1994, 18, 94008066. [Google Scholar] [CrossRef]
- Furedi-Milhofer, H.; Moradian-Oldak, J.; Weiner, S.; Veis, A.; Mintz, K.P.; Addadf, L. Interactions of matrix proteins from mineralized tissues with octacalcium phosphate. Connect. Tissue Res. 1994, 30, 251–264. [Google Scholar] [CrossRef]
- Rey, C. Calcium Phosphates for Medical Applications. In Calcium Phosphates in Biological and Industrial Systems; Springer: New York, NY, USA, 1998; pp. 217–251. [Google Scholar]
- Hanein, D.; Geiger, B.; Addadi, L. Fibronectin Adsorption to Surfaces of Hydrated Crystals. An Analysis of the Importance of Bound Water in Protein-Substrate Interactions. Langmuir 1993, 9, 1058–1065. [Google Scholar] [CrossRef]
- Abbona, F.; Christensson, F.; Angela, M.F.; Madsen, H.E.L. Crystal habit and growth conditions of brushite, CaHPO4 2H2O. J. Cryst. Growth 1993, 131, 331–346. [Google Scholar] [CrossRef]
- Markovic, M. Octacalcium Phosphate; Karger: Basel, Switzerland, 2001; Volume 18, pp. 77–93. [Google Scholar]
- Monma, H.; Moriyoshi, Y. Zeolitic dehydration-rehydration of adipate-intercalated octacalcium phosphate. J. Mater. Sci. 1990, 1, 21–25. [Google Scholar] [CrossRef]
- Nancollas, G.H.; Wu, W. Biomineralization mechanisms: A kinetics and interfacial energy approach. J. Cryst. Growth 2000, 211, 137–142. [Google Scholar] [CrossRef]
- Brown, W.E.; Mathew, M.; Chow, L.C. Adsorption on and Surface Chemistry of Hydroxyapatite; Springer: New York, NY, USA, 1984; pp. 13–28. [Google Scholar]
- Bigi, A.; Bracci, B.; Panzavolta, S.; Iliescu, M.; Plouet-Richard, M.; Werckmann, J.; Cam, D. Morphological and Structural Modifications of Octacalcium Phosphate Induced by Poly-L-Aspartate. Cryst. Growth Des. 2004, 4, 141–146. [Google Scholar] [CrossRef]
- LeGeros, R.Z. Preparation of octacalcium phosphate (OCP): A direct fast method. Calcif. Tissue Int. 1985, 37, 194–197. [Google Scholar] [CrossRef]
- Buljan Meić, I.; Kontrec, J.; Domazet Jurašin, D.; Njegić Džakula, B.; Štajner, L.; Lyons, D.M.; Dutour Sikirić, M.; Kralj, D. Comparative Study of Calcium Carbonates and Calcium Phosphates Precipitation in Model Systems Mimicking the Inorganic Environment for Biomineralization. Cryst. Growth Des. 2017, 17, 1103–1117. [Google Scholar] [CrossRef]
- Wang, L.; Nancollas, G.H. Pathways to biomineralization and biodemineralization of calcium phosphates: The thermodynamic and kinetic controls. Dalton Trans. 2009, 2665–2672. [Google Scholar] [CrossRef]
- Selmani, A.; Coha, I.; Magdić, K.; Čolović, B.; Jokanović, V.; Šegota, S.; Gajović, S.; Gajović, A.; Jurašin, D.; Dutour Sikirić, M. Multiscale study of the influence of cationic surfactants on amorphous calcium phosphate precipitation. Cryst. Eng. Comm. 2015, 17, 8529–8548. [Google Scholar] [CrossRef] [Green Version]
- Ofir, P.B.-Y.; Govrin-Lippman, R.; Garti, N.; Füredi-Milhofer, H. The Influence of Polyelectrolytes on the Formation and Phase Transformation of Amorphous Calcium Phosphate. Cryst. Growth Des. 2004, 4, 177–183. [Google Scholar] [CrossRef]
- Li, S.; Wang, L. Phosphorylated osteopontin peptides inhibit crystallization by resisting the aggregation of calcium phosphate nanoparticles. Cryst. Eng. Comm. 2012, 14, 8037–8043. [Google Scholar] [CrossRef]
- Jahromi, T.M.; Cerruti, M. Amino acid/ion aggregate formation and their role in hydroxyapatite precipitation. Cryst. Growth Des. 2015, 15, 1096–1104. [Google Scholar] [CrossRef]
- Suzuki, O. Octacalcium phosphate: Osteoconductivity and crystal chemistry. Acta Biomater. 2010, 6, 3379–3387. [Google Scholar] [CrossRef] [PubMed]
- Koutsopoulos, S. Synthesis and characterization of hydroxyapatite crystals: A review study on the analytical methods. J. Biomed. Mater. Res. 2002, 62, 600–612. [Google Scholar] [CrossRef]
- Fowler, B.O.; Marković, M.; Brown, W.E. Octacalcium Phosphate. 3. Infrared and Raman Vibrational Spectra. Chem. Mater. 1993, 5, 1417–1423. [Google Scholar] [CrossRef]
- Nancollas, G.H.; Wefel, J.S. Seeded Growth of Calcium Phosphates: Effect of Different Calcium Phosphate Seed Material. J. Dent. Res. 1976, 55, 617–624. [Google Scholar] [CrossRef]
- Bigi, A.; Boanini, E.; Falini, G.; Panzavolta, S.; Roveri, N. Effect of sodium polyacrylate on the hydrolysis of octacalcium phosphate. J. Inorg. Biochem. 2000, 78, 227–233. [Google Scholar] [CrossRef]
- Bigi, A.; Boanini, E.; Bracci, B.; Falini, G.; Rubini, K. Interaction of acidic poly-amino acids with octacalcium phosphate. J. Inorg. Biochem. 2003, 95, 291–296. [Google Scholar] [CrossRef]
- Dorozhkin, S. V Amorphous calcium (ortho)phosphates. Acta Biomater. 2010, 6, 4457–4475. [Google Scholar] [CrossRef]
- Karampas, I.A.; Kontoyannis, C.G. Characterization of calcium phosphates mixtures. Vib. Spectroscop. 2013, 64, 126–133. [Google Scholar] [CrossRef]
- Xu, J.; Butler, I.S.; Gilson, D.F.R. FT-Raman and high-pressure infrared spectroscopic studies of dicalcium phosphate dihydrate (CaHPO4·2H2O) and anhydrous dicalcium phosphate (CaHPO4). Spectochim. Acta Part A 1999, 55, 2801–2809. [Google Scholar] [CrossRef]
- Obadia, L.; Rouillon, T.; Bujoli, B.; Daculsi, G.; Bouler, J.M. Calcium-deficient apatite synthesized by ammonia hydrolysis of dicalcium phosphate dihydrate: Influence of temperature, time, and pressure. J. Biomed. Mater. Res. Part B 2007, 80, 32–42. [Google Scholar] [CrossRef]
- Tas, A.C.; Bhaduri, S.B. Chemical Processing of CaHPO4·2H2O: Its Conversion to Hydroxyapatite. J. Am. Ceram. Soc. 2004, 87, 2195–2200. [Google Scholar] [CrossRef] [Green Version]
- Štulajterová, R.; Medvecký, Ľ. Effect of calcium ions on transformation brushite to hydroxyapatite in aqueous solutions. Colloids Surf. A 2008, 316, 104–109. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | γ/ppm | tind/min | |
|---|---|---|---|
| OCP Seed | DCPD Seed | ||
| Asp | 0 | 46.90 ± 3.34 | 44.68 ± 2.39 |
| 5 | 31.47 ± 3.24 | 42.29 ± 2.90 | |
| 10 | 34.65 ± 2.62 | 47.38 ± 2.98 | |
| Lys | 5 | 65.17 ± 4.51 | 47.12 ± 2.15 |
| 10 | 34.36 ± 3.15 | 31.33 ± 2.95 | |
| Asn | 5 | 56.31 ± 4.23 | 39.77 ± 0.96 |
| 10 | 58.88 ± 2.23 | 32.18 ± 3.13 | |
| Ser | 10 | 90.46 ± 7.20 | 41.80 ± 4.41 |
| Tyr | 5 | 65.15 ± 4.63 | 46.01 ± 0.74 |
| 10 | 53.68 ± 4.64 | 48.68 ± 4.41 | |
| Phe | 5 | 23.91± 1.73 | 18.03 ± 1.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihelj Josipović, T.; Kovačević, M.; Mateša, S.; Kostešić, M.; Matijaković, N.; Radatović, B.; Lyons, D.M.; Kralj, D.; Dutour Sikirić, M. The Influence of Different Classes of Amino Acids on Calcium Phosphates Seeded Growth. Materials 2020, 13, 4798. https://doi.org/10.3390/ma13214798
Mihelj Josipović T, Kovačević M, Mateša S, Kostešić M, Matijaković N, Radatović B, Lyons DM, Kralj D, Dutour Sikirić M. The Influence of Different Classes of Amino Acids on Calcium Phosphates Seeded Growth. Materials. 2020; 13(21):4798. https://doi.org/10.3390/ma13214798
Chicago/Turabian StyleMihelj Josipović, Tea, Monika Kovačević, Sarah Mateša, Marina Kostešić, Nives Matijaković, Borna Radatović, Daniel M. Lyons, Damir Kralj, and Maja Dutour Sikirić. 2020. "The Influence of Different Classes of Amino Acids on Calcium Phosphates Seeded Growth" Materials 13, no. 21: 4798. https://doi.org/10.3390/ma13214798
APA StyleMihelj Josipović, T., Kovačević, M., Mateša, S., Kostešić, M., Matijaković, N., Radatović, B., Lyons, D. M., Kralj, D., & Dutour Sikirić, M. (2020). The Influence of Different Classes of Amino Acids on Calcium Phosphates Seeded Growth. Materials, 13(21), 4798. https://doi.org/10.3390/ma13214798