
Stimulation of Metabolic Activity and Cell Differentiation in Osteoblastic and Human Mesenchymal Stem Cells by a Nanohydroxyapatite Paste Bone Graft Substitute

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of nHA Paste-Conditioned Culture Medium Extract
2.2. Cell Culture
2.3. Cell Viability Assays
2.4. Determination of the Ca2+ Ions Released from the nHA Paste
2.5. Study of the Effect of Ions Released from the Paste on Cell Metabolism and Their Potential Mechanism
2.6. CaSR Agonist, 3-(Trifluoromethyl) Cinnamic Acid
2.7. CaSR Antagonist, NPS2143 Hydrochloride
2.8. Measurement of Gene Expression
3. Results
3.1. Effect of nHA Paste-Conditioned Medium on Cell Metabolic Activity
3.2. Cell Metabolic Response of Human Osteoblastic Cells, hBM-MSCs, and Murine Myoblastic Cells to nHA paste-Conditioned Medium (PCM)
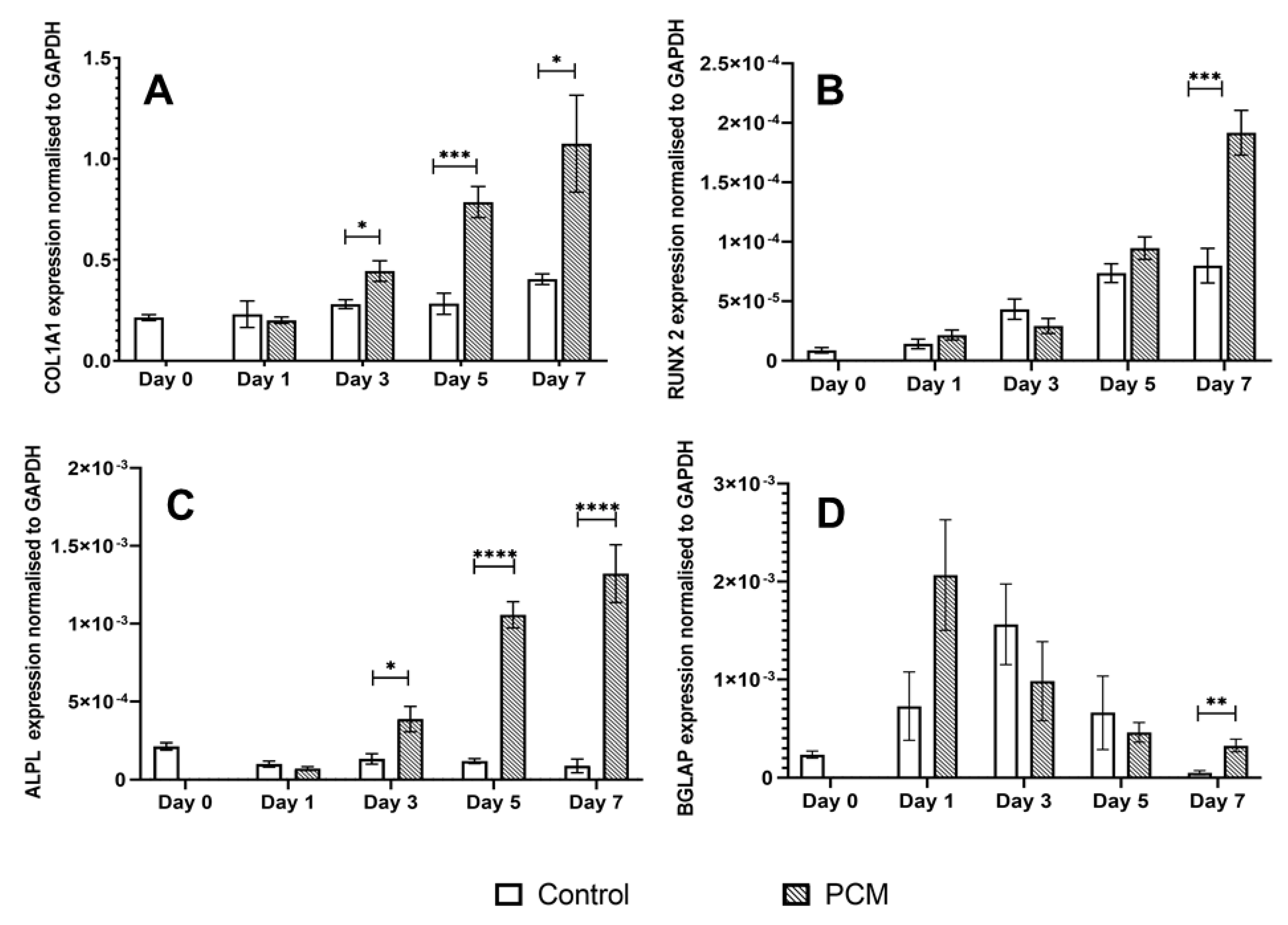
3.3. Osteogenic Potential of the PCM
3.4. Determination of Ca2+ and Na+ Ion Concentrations in the PCM Samples
3.5. Effect of Low Ion Concentrations on Cellular Metabolic Activity
3.6. Effect of PCM on the Calcium-Sensing Receptor (CaSR) of Osteoblastic Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carter, D.H.; Hatton, P.V.; Aaron, J.E. The Ultrastructure of Slam-Frozen Bone Mineral. Histochem. J. 1997, 29, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Bala, Y.; Seeman, E. Bone’s material constituents and their contribution to bone strength in health, disease and treatment. Calcif. Tissue Int. 2015, 97, 308–326. [Google Scholar] [CrossRef] [PubMed]
- LeGeros, R.Z. Calcium phosphate-based osteoinductive materials. Chem. Rev. 2008, 108, 4742–4753. [Google Scholar] [CrossRef] [PubMed]
- Szpalski, C.; Wetterau, M.; Barr, J.; Warren, S.M. Bone tissue engineering: Current strategies and techniques—Part I: Scaffolds. Tissue Eng. Part B Rev. 2012, 18, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Tadic, D. A thorough physicochemical characterisation of 14 calcium phosphate-based bone substitution materials in comparison to natural bone. Biomaterials 2004, 25, 987–994. [Google Scholar] [CrossRef]
- Zakaria, S.M.; Zein, S.H.; Othman, M.R.; Yang, F.; Jansen, J.A. Nanophase hydroxyapatite as a biomaterial in advanced hard tissue engineering: A review. Tissue Eng. Part B Rev. 2013, 19, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Harrison, C.J.; Hatton, P.V.; Gentile, P.; Miller, C.A. Nano-scale strontium-substituted hydroxyapatite pastes and gels for bone tissue regeneration. Nanomaterials 2021, 11, 161. [Google Scholar] [CrossRef]
- Sadat-Shojai, M.; Khorasani, M.-T.; Dinpanah-Khoshdargi, E.; Jamshidi, A. Synthesis methods for nanosized hydroxyapatite with diverse structures. Acta Biomater. 2013, 9, 7591–7621. [Google Scholar] [CrossRef]
- Molino, G.; Palmieri, M.C.; Montalbano, G.; Fiorilli, S.; Vitale-Brovarone, C. Biomimetic and mesoporous nano-hydroxyapatite for bone tissue application: A short review. Biomed. Mater. 2020, 15, 022001. [Google Scholar] [CrossRef]
- Lowe, B.; Hardy, J.G.; Walsh, L.J. Optimising nanohydroxyapatite nanocomposites for bone tissue engineering. ACS Omega 2021, 5, 1–9. [Google Scholar] [CrossRef]
- Fathi, M.H.; Hanifi, A.; Mortazavi, V. Preparation and bioactivity evaluation of bone-like hydroxyapatite nanopowder. J. Mater. Process. Technol. 2008, 202, 536–542. [Google Scholar] [CrossRef]
- Hruschka, V.; Tangl, S.; Ryabenkova, Y.; Heimel, P.; Barnewitz, D.; Möbus, G.; Keibl, C.; Ferguson, J.; Quadros, P.; Miller, C.A.; et al. Comparison of nanoparticular hydroxyapatite pastes of different particle content and size in a novel scapula defect model. Sci. Rep. 2017, 7, 43425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, P.; Wilcock, C.J.; Miller, C.A.; Moorehead, R.; Hatton, P.V. Process Optimisation to Control the Physico-Chemical Characteristics of Biomimetic Nanoscale Hydroxyapatites Prepared Using Wet Chemical Precipitation. Materials 2015, 8, 2297–2310. [Google Scholar] [CrossRef]
- Cai, Y.; Liu, Y.; Yan, W.; Hu, Q.; Tao, J.; Zhang, M.; Shi, Z.; Tang, R. Role of hydroxyapatite, nanoparticle size in bone cell proliferation. J. Mater. Chem. 2007, 17, 3730–3738. [Google Scholar] [CrossRef]
- Dorozhkin, S.V. Nanosized and nanocrystalline calcium orthophosphates. Acta Biomater. 2010, 6, 715–734. [Google Scholar] [CrossRef] [PubMed]
- Huber, F.X.; Belyaev, O.; Hillmeier, J.; Kock, H.J.; Huber, C.; Meeder, P.J.; Berger, I. First histological observations on the incorporation of a novel nanocrystalline hydroxyapatite paste OSTM on human cancellous bone. BMC Musculoskelet. Disord. 2006, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Canuto, R.A.; Pol, R.; Maratinasso, G.; Muzio, G.; Gallesio, G.; Mozzati, M. Hydroxyapatite paste Ostim®, without elevation of full-thickness flaps, improved alveolar healing stimulating BMP- and VEGF-mediated signal pathways: An experimental study in humans. Clin. Oral Implants Res. 2013, 24, 42–48. [Google Scholar] [CrossRef]
- Conteduca, F.; di Sette, P.; Iorio, R.; Caperna, L.; Argento, G.; Mazza, D.; Ferretti, A. Nanohydroxyapatite promotes the healing process in open-wedge high tibial osteotomy: A CT study. Knee 2016, 23, 763–769. [Google Scholar] [CrossRef]
- Gao, H.; Shi, W.; Freund, L.B. Mechanics of receptor-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2005, 102, 9469–9474. [Google Scholar] [CrossRef] [Green Version]
- Decuzzi, P.; Ferrari, M. The receptor-mediated endocytosis of nonspherical particles. Biophys. J. 2008, 94, 3790–3797. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.T.; Nie, G.J.; Meng, H.; Xia, T.; Nel, A.; Zhao, Y.L. Physicochemical properties determine nanomaterial cellular uptake, transport, and fate. Acc. Chem. Res. 2013, 46, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Motskin, M.; Wright, D.M.; Muller, K.; Kyle, N.; Gard, T.G.; Porter, A.E.; Skepper, J.N. Hydroxyapatite nano and microparticles: Correlation of particle properties with cytotoxicity and biostability. Biomaterials 2009, 30, 3307–3317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, J.; Lykotrafitis, G.; Bao, G.; Suresh, S. Size-Dependent Endocytosis of Nanoparticles. Adv. Mater. 2009, 21, 419–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Huang, X.; Cai, Y.; Tang, R.; Yang, D. Size effect of hydroxyapatite nanoparticles on proliferation and apoptosis of osteoblast-like cells. Acta Biomater. 2009, 5, 338–345. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, M.; Lu, J.; Ma, J.; Wei, J.; Wei, S. Cell responses to two kinds of nanohydroxyapatite with different sizes and crystallinities. Int. J. Nanomed. 2012, 7, 1239–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Ng, S.; Heng, B.C.; Guo, J.; Ma, L.; Tan, T.Y.T.; Ng, K.W.; Loo, S.C.J. Cytotoxicity of hydroxyapatite nanoparticles is shape and cell dependent. Arch. Toxicol. 2013, 87, 1037–1052. [Google Scholar] [CrossRef]
- Dey, S.; Das, M.; Balla, V.K. Effect of hydroxyapatite particle size, morphology and crystallinity on proliferation of colon cancer HCT116 cells. Mater. Sci. Eng. C 2014, 39, 336–339. [Google Scholar] [CrossRef]
- Chen, L.; Mccrate, J.M.; Lee, J.C.-M.; Li, H. The role of surface charge on the uptake and biocompatibility of hydroxyapatite nanoparticles with osteoblast cells. Nanotechnology 2011, 22, 105708. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, V.; Kozlova, D.; Knuschke, T.; Buer, J.; Westendorf, A.M.; Epple, M. Mechanism of the uptake of of cationic and anionic calcium phosphate nanoparticles by cells. Acta Biomater. 2013, 9, 7527–7535. [Google Scholar] [CrossRef]
- Kopp, M.; Rotan, O.; Papadopoullos, C.; Schulze, N.; Meyer, H.; Epple, M. Delivery of the autofluorescent protein R-phycoerythrin by calcium phosphate nanoparticles into four different eukaryotic cell lines (HeLa, HEK293T, MG63, MC3T3): Highly efficient, but leading to endolysosomal proteolysis in HeLa and MC3T3 cells. PLoS ONE 2017, 12, e0178260. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Liu, C.; Sun, J. Effects of four types of hydroxyapatite nanoparticles with different nanocrystal morphologies and sizes on apoptosis in rat osteoblasts. J. Appl. Toxicol. 2011, 32, 429–435. [Google Scholar] [CrossRef]
- Epple, E. Review of potential risks associated with nanoscopic calcium phosphate. Acta Biomater. 2018, 77, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Remya, N.S.; Syama, S.; Gayathri, V.; Varma, H.K.; Mohanan, P.V. An in vitro study on the interaction of hydroxyapatite nanoparticles and bone marrow mesenchymal stem cells for assessing the toxicological behaviour. Colloids Surf. B Biointerfaces 2014, 117C, 389–397. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, G.; Liu, H.; Niu, X.F.; Han, J.Y.; Zheng, L.H.; Fan, Y.B. Nano-hydroxyapatite particles induce apoptosis on MC3T3-E1 cells and tissue cells in SD rats. Nanoscale 2012, 4, 2894–2899. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Jackson, S.; Haycock, J.W.; MacNeil, S. Culture of skin cells in 3D rather than 2D improves their ability to survive exposure to cytotoxic agents. J. Biotechnol. 2006, 122, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lilly, G.D.; Doty, R.C.; Podsiadlo, P.; Kotov, N.A. In vitro toxicity testing of nanoparticles in 3D cell culture. Small 2009, 5, 1213–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavasi, R.-M.; Coelho CCPlatania, V.; Quadros, P.A.; Chatzinikolaidou, M. In vitro biocompatability assessment of nano-hydroxyapatite. Nanomaterials 2021, 11, 1152. [Google Scholar] [CrossRef]
- Hannan, F.M.; Kallay, E.; Chang, W.; Brandi, M.L.; Thakker, R.V. The calcium-sensing receptor in physiology and in calcitropic and non-calcitropic diseases. Nat. Rev. Endocrinol. 2019, 15, 33–51. [Google Scholar] [CrossRef]
- Brown, E.M. Role of the calcium-sensing receptor in extracellular calcium homeostasis. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 333–343. [Google Scholar] [CrossRef]
- Goltzman, D.; Hendy, G.N. The calcium-sensing receptor in bone-mechanistic and therapeutic insights. Nature 2015, 11, 298–307. [Google Scholar] [CrossRef]
- Dvorak-Ewell Chen, T.-H.; Liang, N.; Garvey, C.; Liu, B.; Tu, C.; Chang, W.; Bikle, D.D.; Shoback, D.M. Osteoblast extracellular Ca2+-sensing receptor regulates bone development, mineralisation and turnover. Am. Soc. Bone Miner. Res. 2011, 26, 2935–2947. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Chattopadhyay, N.; Kifor, O.; Ye, C.P.; Vassilev, P.M.; Sanders, J.L.; Brown, E.M. Expression of extracellular calcium-sensing receptor in human osteoblastic MG-63 cell line. Am. J. Physiol. Cell Physiol. 2001, 280, C382–C393. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Kifor, O.; Chattopadhyay, N.; Brown, E.M. Expression of extracellular calcium (Ca2+)-sensing receptor in the clonal osteoblast-like cell lines, UMR-106 and SAOS-2. Biochem. Biophys. Res. Commun. 1998, 243, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.O.; Kwak, J.; Kim, H.W.; Oh, K.-J.; Kim, Y.T.; Jung, S.M.; Cha, S.H. The extracellular calcium sensing receptor is expressed in mouse mesangial cells and modulates cell proliferation. Exp. Mol. Med. 2005, 37, 457–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, F.; Zhang, G.; Liu, L.; Jiang, L.; Wang, R.; Zheng, Y.; Wang, G.; Xie, M.; Duan, Y. Involvement of cationic channels in proliferation and migration of human mesenchymal stem cells. Tissue Cell. 2012, 44, 358–364. [Google Scholar] [CrossRef]
- González-Vázquez, A.; Planell, J.A.; Engel, E. Extracellular calcium and CaSR drive osteoinduction in mesenchymal stromal cells. Acta Biomater. 2014, 10, 2824–2833. [Google Scholar] [CrossRef]
- Marie, P.J. The calcium-sensing receptor in bone cells: A potential therapeutic target in osteoporosis. Bone 2010, 46, 571–576. [Google Scholar] [CrossRef]
- Al-Dujaili, S.A.; Koh, A.J.; Dang, M.; Chang, W.; Ma, P.X.; McCauley, L.K. Calcium sensing receptor function supports osteoblast survival and acts as a co-factor in PTH anabolic actions in bone. J. Cell Biochem. 2016, 117, 1556–1567. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, A.R.B.; Hvidtfeld, T.M.; Bräuner-Osborne, H. Biased agonism of the calcium-sensing receptor. Cell Calcium 2012, 51, 107–116. [Google Scholar] [CrossRef]
- Davey, A.E.; Leach, K.; Valant, C.; Conigrave, A.D.; Sexton, P.M.; Christopoulos, A. Positive and negative allosteric modulators promote biased signaling at the calcium-sensing receptor. Endocrinology 2012, 153, 1232–1241. [Google Scholar] [CrossRef] [Green Version]
- Thiel, O.R.; Bernard, C.; Tormos, W.; Brewin, A.; Hirotani, S.; Murakami, K.; Saito, K. Practical synthesis of the calcimimetic agent, cinacalcet. Tetrahedron Lett. 2008, 49, 13–15. [Google Scholar] [CrossRef]
- Nemeth, E.F. The search for calcium receptor antagonists (calcilytics). J. Mol. Endocrinol. 2002, 29, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, H.; Cailly, T.; Rojas, B.I.E.; Thomsen, A.; Bräuner-Osborne, H.; Sejer Pedersen, D. Synthesis of the calcilytic ligand NPS 2143. Beilstein J. Org. Chem. 2013, 9, 1383–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschi, R.T.; James, W.M.; Zerlauth, G. 1 alpha, 25-dihydroxyvitamin D3 specific regulation of growth, morphology and fibronectin in a human osteosarcoma cell line. J Cell Physiol. 1985, 123, 401–409. [Google Scholar] [CrossRef]
- Rodan, S.B.; Imai, Y.; Theide, M.A.; Wesolowski, G.; Thompson, D.; Bar-Shavit, Z.; Shull, S.; Mann KRodan, G.A. Characterisation of a human osteosarcoma cell line (Saos-2) with osteogenic properties. Cancer Res. 1987, 47, 4961–4966. [Google Scholar] [PubMed]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In search of an osteoblastic cell model for in vitro research. Eur. Cells Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ryabenkova, Y.; Pinnock, A.; Quadros, P.A.; Goodchild, R.L.; Mobus, G.; Crawford, A.; Hatton, P.V.; Miller, C.A. The relationship between particle morphology and rheological properties in injectable nano-hydroxyapatite bone graft substitutes. Mater. Sci. Eng. C 2017, 75, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Cao, T.; Saw, T.Y.; Heng, B.C.; Liu, H.; Yap, A.U.J.; Ng, M.L. Comparison of different test models for the assessment of cytotoxicity of composite resins. J. Appl. Toxicol. 2005, 25, 101–108. [Google Scholar] [CrossRef]
- Sodium phosphate. Cold Spring Harb. Protoc. 2006. [CrossRef]
- Yong, K.W.; Pingguan-Murphy, B.; Xu, F.; Abas, W.A.B.W.; Choi, J.R.; Omar, S.Z.; Azmi, M.A.N.; Chua, K.H.; Safwani, W.K.Z.M. Phenotypic and functional characterisation of long-term cryopreserved human adipose-derived stem cells. Sci. Rep. 2015, 5, 9596. [Google Scholar] [CrossRef] [Green Version]
- Mandatori, D.; Penolazzi, L.; Pelusi, L.; Lambertini, E.; Michelucci, F.; Porreca, A.; Cerritelli, P.; Pipino, C.; Di Inorio, A.; Bruni, D.; et al. Three-Dimensional Co-Culture System of Human Osteoblasts and Osteoclast Precursors from Osteoporotic Patients as an Innovative Model to Study the Role of Nutrients: Focus on Vitamin K2. Nutrients 2021, 13, 2823. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.H.; Moon, R.J. Pathophysiology of hypercalcaemia. In Encyclopedia of Endocrine Disorders, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2019; Available online: https://www.sciencedirect.com/science/article/pii/B970128012383652467 (accessed on 7 January 2022).
- Gustavsson, J.; Ginebra, M.P.; Engel, E.; Planell, J. Ion reactivity of calcium-deficient hydroxyapatite in standard cell culture media. Acta Biomater. 2011, 7, 4242–4252. [Google Scholar] [CrossRef] [PubMed]
- Maeno, S.; Niki, Y.; Matsumoto, H.; Morioka, H.; Yatabe, T.; Funayama, A.; Toyama, Y.; Tagushi, T.; Tanaka, J. The effect of calcium ion concentration on osteoblast viability, proliferation and differentiation in monolayer and 3D culture. Biomaterials 2005, 26, 4847–4855. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, J.; Ginebra, M.P.; Planell, J.; Engel, E. Osteoblast-like cellular response to dynamic changes in the ionic extracellular environment produced by calcium-deficient hydroxyapatite. J. Mater. Sci. Mater. Med. 2012, 23, 2509–2520. [Google Scholar] [CrossRef] [PubMed]
- Barradas, A.M.C.; Fernandes, H.A.M.; Groen, N.; Chai, Y.C.; Schrooten, J.; van de Peppel, J.; van Leeuwen, J.P.; van Blitterswijk, C.A.; de Boer, J. A calcium-induced signalling cascade leading to osteogenic differentiation of human bone marrow-derived mesenchymal stromal cells. Biomaterials 2012, 33, 3205–3215. [Google Scholar] [CrossRef]
- Hu, F.; Pan, L.; Zhang, K.; Xing, F.; Wang, X.; Lee, I.; Zhang, Z.; Xu, J. Elevation of Extracellular Ca2+ Induces Store-Operated Calcium Entry via Calcium-Sensing Receptors: A Pathway Contributes to the Proliferation of Osteoblasts. PLoS ONE 2014, 9, e107217. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, M.M.; Siddiqua, A.; Ward, D.T.; Carter, D.H.; Dallas, S.L.; Nemeth, E.F.; Riccardi, D. Physiological changes in extracellular calcium concentration directly control osteoblast function in the absence of calciotropic hormones. Proc. Natl. Acad. Sci. USA 2004, 101, 5140–5145. [Google Scholar] [CrossRef] [Green Version]
- Pilloni, A.; Pompa, G.; Sccucci, M.; di Carlo, G.; Rimondi, L.; Brama, M.; Zeeza, B.; Wannenes, F.; Migliaccio, S. Analysis of human alveolar osteoblast behaviour on a nano-hydroxyapatite substrate: An in vitro study. BMC Oral Health 2014, 14, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, S.-H.; Park, K.; Habib, M.M.; Beck, G.R., Jr. Nano-hydroxyapatite stimulation of gene expression requires Fgf receptor, phosphate transporter, and Erk1/2 signalling. ACS Appl. Mater. Interfaces 2017, 9, 39185–39196. [Google Scholar] [CrossRef]
- Ha, S.-W.; Jang, H.L.; Nam, K.T.; Beck, G.R., Jr. Nano-hydroxyapaptite modulates osteoblast lineage commitment by stimulation of DNA methylation and regulation of gene expression. Biomaterials. 2015, 65, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Sarem, M.; Heizmann, M.; Barbero, A.; Martin, I.; Shastri, V.P. Hyperstimulation of CaSR in human MSCs by biomimetic apatite inhibits endochondral ossification via temporal down-regulation of PTH1R. Proc. Natl. Acad. Sci. USA 2018, 115, E6135–E6144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, E.F.; Delmar, E.G.; Heaton, W.L.; Miller, M.A.; Lambert, L.D.; Conklin, R.L.; Gowen, M.; Gleason, J.G.; Bhatnagar, P.K.; Fox, J. Calcilytic Compounds: Potent and Selective Ca2+ Receptor antagonists that stimulate secretion of parathyroid hormone. J. Pharmacol. Exp. Ther. 2001, 299, 323–331. [Google Scholar] [PubMed]
- Centeno, P.P.; Herberger, A.; Mun, H.C.; Tu, C.; Nemeth, E.F.; Chang, W.; Conigrave, A.D.; Ward, D.T. Phosphate acts directly on the calcium-sensing receptor to stimulate parathyroid hormone secretion. Nat. Commun. 2019, 10, 4693. [Google Scholar] [CrossRef]
- Suto, M.; Nemoto, E.; Kanaya, S.; Suzuki, R.; Tsuchiya, M.; Shimauchi, H. Nanohydroxyapatite increases BMP-2 expression via a p38 MAP kinase dependent pathway in periodontal ligament cells. Arch. Oral Biol. 2013, 58, 1021–1028. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Na+ (mM) | Ca2+ (mM) |
|---|---|---|
| CM (theoretical values) | 143.5 | 1.80 |
| CM | 154.30 | 1.85 ± 0.04 |
| PCM | 133.40 | 0.45 ± 0.04 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herranz-Diez, C.; Crawford, A.; Goodchild, R.L.; Hatton, P.V.; Miller, C.A. Stimulation of Metabolic Activity and Cell Differentiation in Osteoblastic and Human Mesenchymal Stem Cells by a Nanohydroxyapatite Paste Bone Graft Substitute. Materials 2022, 15, 1570. https://doi.org/10.3390/ma15041570
Herranz-Diez C, Crawford A, Goodchild RL, Hatton PV, Miller CA. Stimulation of Metabolic Activity and Cell Differentiation in Osteoblastic and Human Mesenchymal Stem Cells by a Nanohydroxyapatite Paste Bone Graft Substitute. Materials. 2022; 15(4):1570. https://doi.org/10.3390/ma15041570
Chicago/Turabian StyleHerranz-Diez, Carolina, Aileen Crawford, Rebecca L. Goodchild, Paul V. Hatton, and Cheryl A. Miller. 2022. "Stimulation of Metabolic Activity and Cell Differentiation in Osteoblastic and Human Mesenchymal Stem Cells by a Nanohydroxyapatite Paste Bone Graft Substitute" Materials 15, no. 4: 1570. https://doi.org/10.3390/ma15041570
APA StyleHerranz-Diez, C., Crawford, A., Goodchild, R. L., Hatton, P. V., & Miller, C. A. (2022). Stimulation of Metabolic Activity and Cell Differentiation in Osteoblastic and Human Mesenchymal Stem Cells by a Nanohydroxyapatite Paste Bone Graft Substitute. Materials, 15(4), 1570. https://doi.org/10.3390/ma15041570