
Exploring In Vivo Pulmonary and Splenic Toxicity Profiles of Silicon Quantum Dots in Mice
 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis of SiQDs and Their Characteristics
2.2. Administration of SiQDs to Mice
2.3. Histology and Immunohistochemistry
2.4. Obtaining the Tissue Homogenates
2.5. Measurement of Antioxidant Enzyme Activities
2.6. Determination of Malondialdehyde Level (MDA)
2.7. Determination Reduced Glutathione (GSH) Level
2.8. Western Blot
2.9. Detection of 8-Hyroxy-2′-deoxyguanosine (8-OHdG)
2.10. Quantification of 5-Methyl Cytosine (5-mC)
2.11. Assessment of Histone Methylation
2.12. Statistical Analysis
3. Results
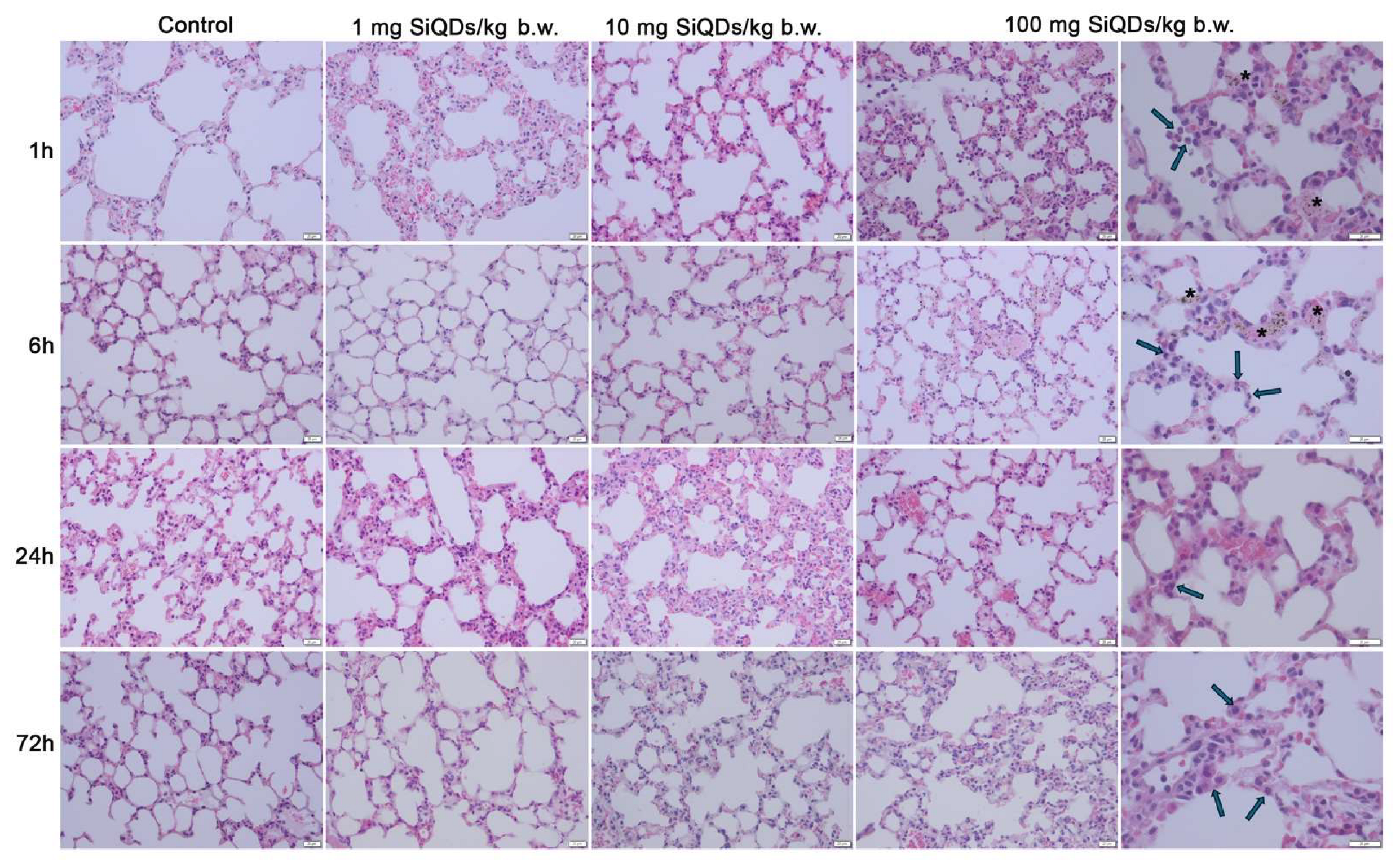
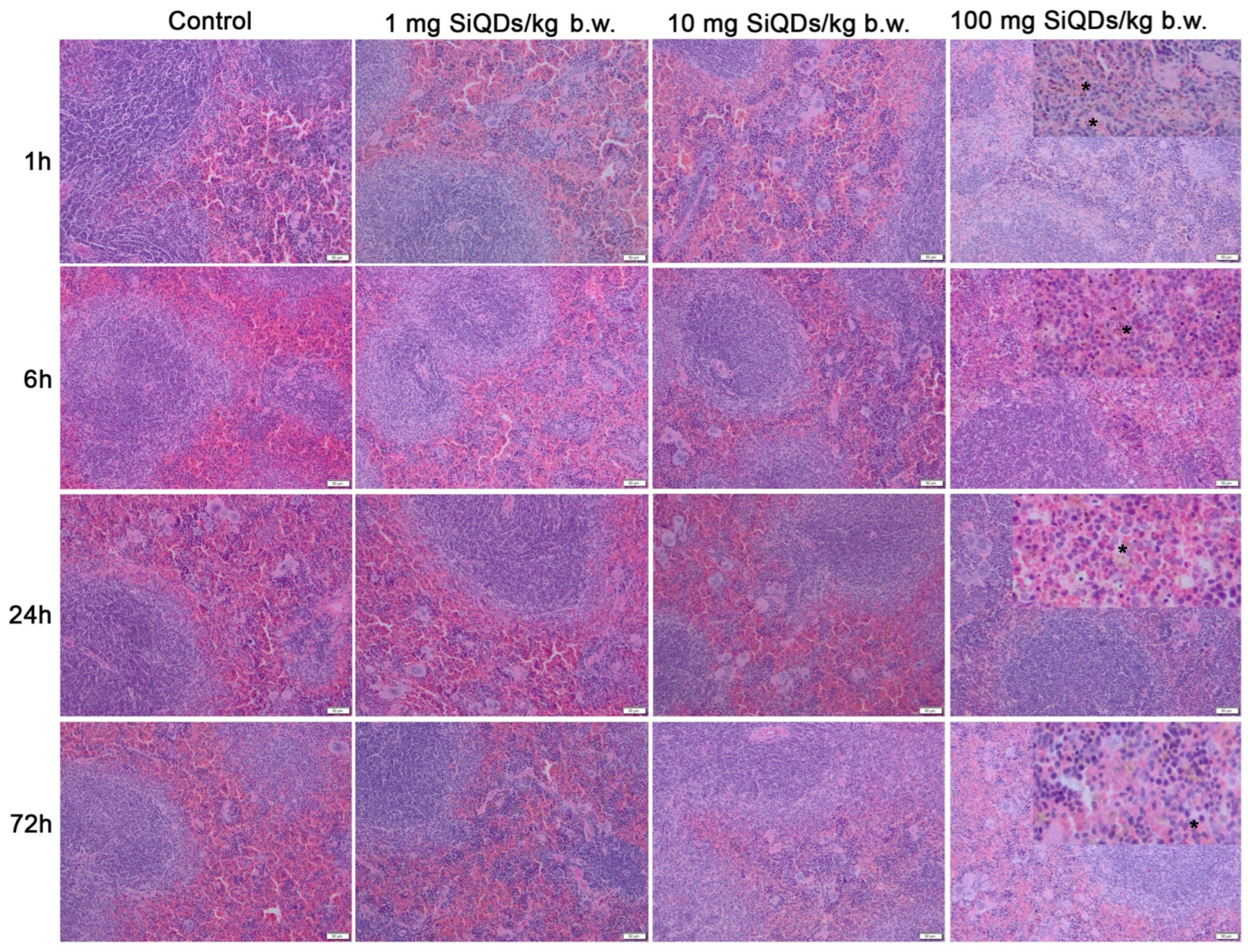
3.1. Histopathology of the Lungs and Spleen of Mice Exposed to SiQDs
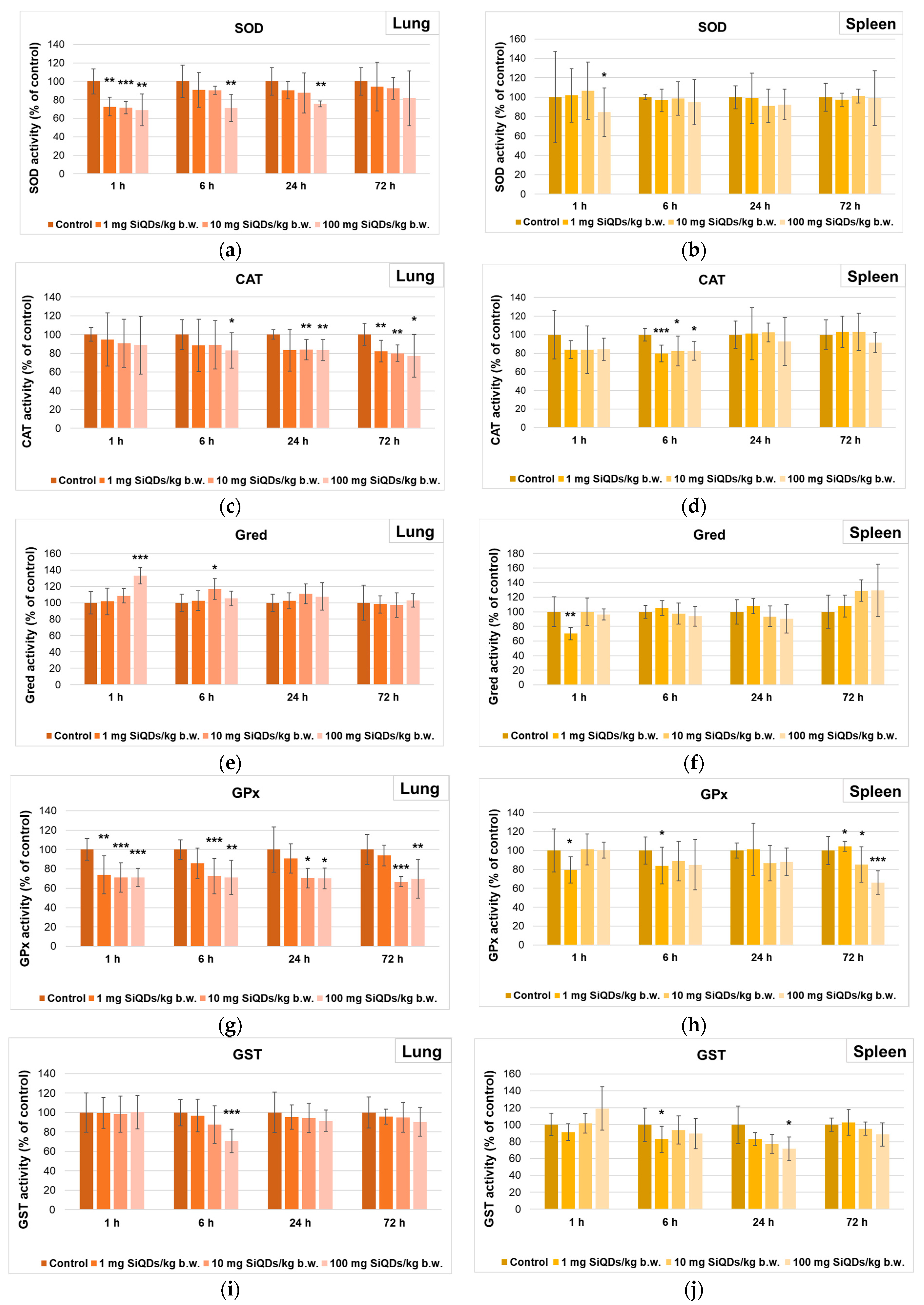
3.2. Analysis of Oxidative Stress Induced by SiQD Administration
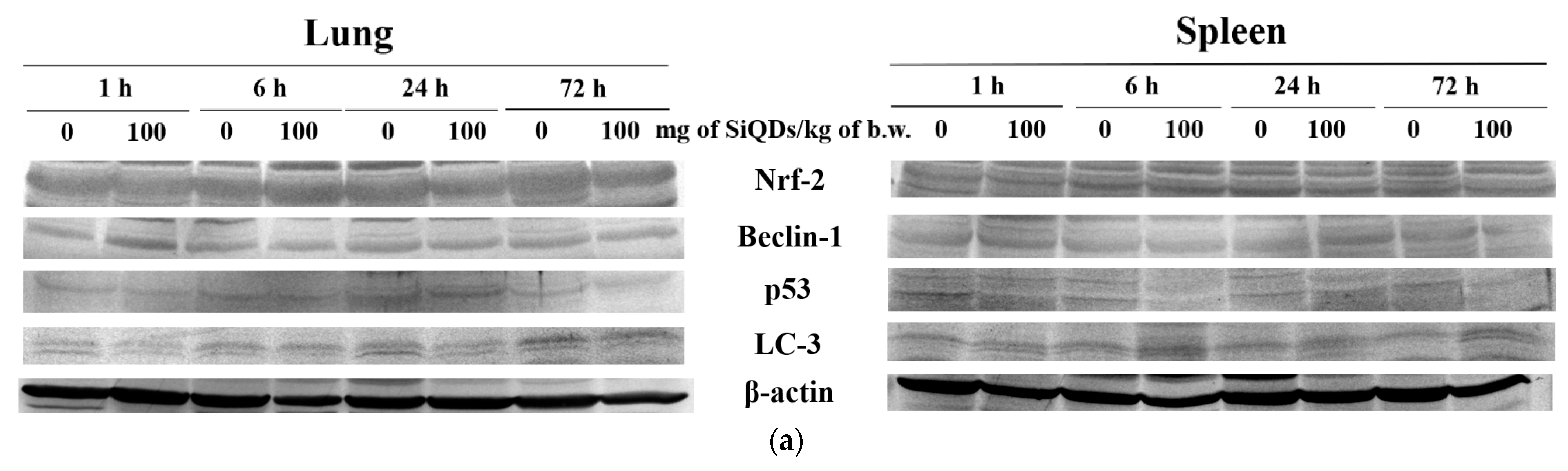
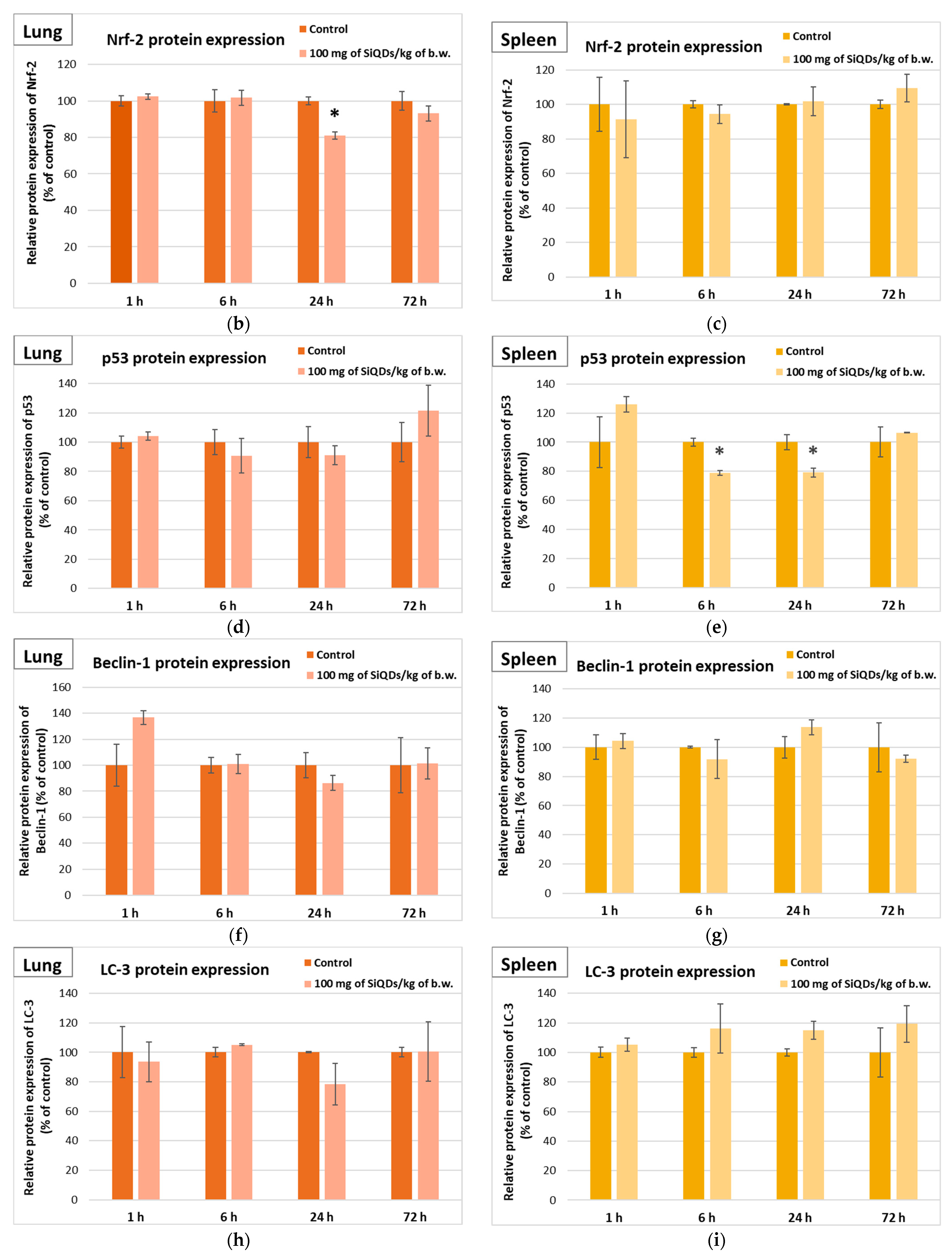
3.3. Analysis of Proteins’ Expression Involved in Antioxidant Defense System, Apoptosis, and Autophagy
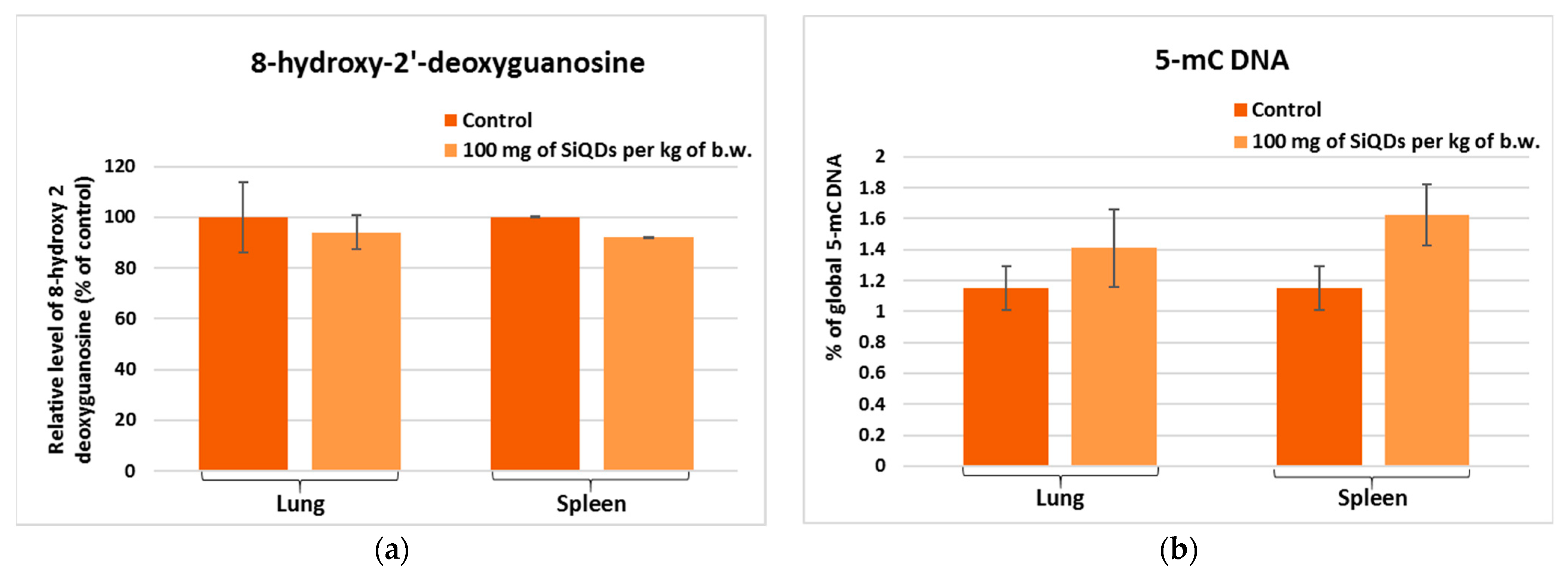
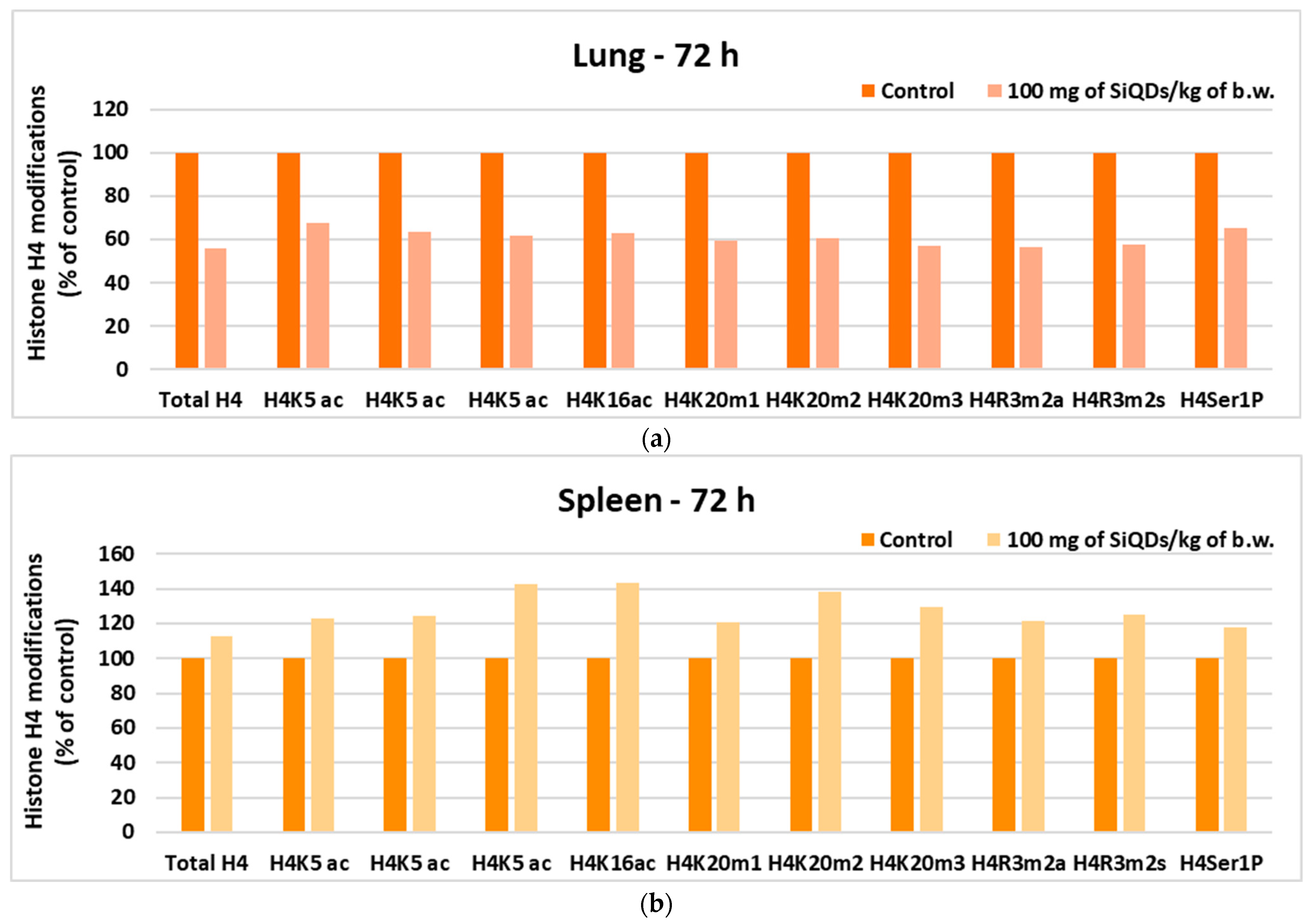
3.4. Genotoxicity Evaluation
3.4.1. Analysis of 8-OHdG Level
3.4.2. Analysis of Histone H4 Modification
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Al-Kayiem, H.; Lin, S.; Lukmon, A. Review on nanomaterials for thermal energy storage technologies. Nanosci. Nanotechnol.-Asia 2013, 3, 60–71. [Google Scholar] [CrossRef]
- Soldado, A.; Barrio, L.C.; Diaz-Gonzalez, M.; de la Escosura-Muniz, A.; Costa-Fernandez, J.M. Advances in quantum dots as diagnostic tools. Adv. Clin. Chem. 2022, 107, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Shen, S.Z.; Sun, H.; Sun, K.; Liu, F. Synthesis of L-glutathione-capped-ZnSe quantum dots for the sensitive and selective determination of copper ion in aqueous solutions. Sens. Actuators B Chem. 2014, 203, 35–43. [Google Scholar] [CrossRef]
- Nam, E.; Lee, C.; Kim, S.J.; Chung, H.K.; Chae, H. Stability and dispersion improvement of quantum-dot films by hydrosilylation between quantum-dot ligands and a siloxane matrix. Opt. Express 2019, 27, 20037–20046. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Rong, X.; Ma, H.; Tao, G. Dithizone functionalized CdSe/CdS quantum dots as turn-on fluorescent probe for ultrasensitive detection of lead ion. J. Hazard. Mater. 2013, 250, 45–52. [Google Scholar] [CrossRef]
- Vinceti, M.; Wei, E.T.; Malagoli, C.; Bergomi, M.; Vivoli, G. Adverse health effects of selenium in humans. Rev. Environ. Health. 2001, 16, 233–251. [Google Scholar] [CrossRef] [PubMed]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Jhonsi, M.A. Carbon quantum dots for bioimaging. State Art Nano-Bioimaging 2018, 20, 72723. [Google Scholar] [CrossRef]
- Sivasankarapillai, V.S.; Jose, J.; Shanavas, M.S.; Marathakam, A.; Uddin, M.S.; Mathew, B. Silicon Quantum dots: Promising theranostic probes for the future. Curr. Drug Targets 2019, 20, 1255–1263. [Google Scholar] [CrossRef]
- Sumanth, K.D.; Jai, K.B.; Mahesh, H.M. Quantum nanostructures (QDS): An overview. Synth. Inorg. Nanomater. 2018, 93, 59–88. [Google Scholar] [CrossRef]
- Svrcek, V.; Sasaki, T.; Shimizu, Y.; Koshizaki, N. Blue luminescent silicon nanocrystals prepared by nanosecond laser ablation and stabilized in electronically compatible spin on glasses. J. Appl. Phys. 2008, 103, 023101. [Google Scholar] [CrossRef]
- Stan, M.S.; Sima, C.; Dinischiotu, A. Silicon Quantum Dots: From synthesis to bioapplications. In Bioactivity of Engineered Nanoparticles; Yan, B., Zhou, H., Gardea-Torresdey, J., Eds.; Springer: Singapore, 2017; pp. 339–359. [Google Scholar] [CrossRef]
- Montalti, M.; Cantelli, A.; Battistelli, G. Nanodiamonds and silicon quantum dots: Ultrastable and biocompatible luminescent nanoprobes for long-term bioimaging. Chem. Soc. Rev. 2015, 44, 4853–4921. [Google Scholar] [CrossRef] [PubMed]
- Sahay, G.; Alakhova, D.Y.; Kabanov, A.V. Endocytosis of nanomedicines. JCR 2010, 145, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Nie, G.; Meng, H.; Xia, T.; Nel, A.; Zhao, Y. Physicochemical properties determine nanomaterial cellular uptake, transport, and fate. Acc. Chem. Res. 2012, 46, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Rauch, J.; Kolch, W.; Laurent, S.; Mahmoudi, M. Big signals from small particles: Regulation of cell signaling pathways by nanoparticles. Chem. Rev. 2013, 113, 3391–3406. [Google Scholar] [CrossRef]
- Li, M.; Gu, M.; Tian, X.; Xiao, B.; Lu, S.; Zhu, W.; Yu, L.; Shang, Z. Hydroxylated graphene quantum dots induce DNA damage and disrupt microtubule structure in human esophageal epithelial cells. Toxicol. Sci. 2018, 164, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.R.; Antonini, J.M.; Porter, D.W.; Chapman, R.S.; Scabilloni, J.F.; Young, S.-H.; Mercer, R.R. Lung toxicity and biodistribution of Cd/Se-ZnS quantum dots with different surface functional groups after pulmonary exposure in rats. Part. Fibre Toxicol. 2013, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Stan, M.S.; Sima, C.; Cinteză, L.O.; Dinischiotu, A. Silicon-based quantum dots induce inflammation in human lung cells and disrupt extracellular matrix homeostasis. FEBS J. 2015, 282, 2914–2929. [Google Scholar] [CrossRef] [PubMed]
- Stan, M.S.; Memet, I.; Sima, C.; Popescu, T.; Teodorescu, V.S.; Hermenean, A.; Dinischiotu, A. Si/SiO2 quantum dots cause cytotoxicity in lung cells through redox homeostasis imbalance. Chem. Biol. Interact. 2014, 220, 102–115. [Google Scholar] [CrossRef]
- Wang, Y.W.; Yang, K.; Tang, H.; Chen, D.; Bai, Y.L. Toxicity assessment of repeated intravenous injections of arginine-glycine-aspartic acid peptide conjugated CdSeTe/ZnS quantum dots in mice. Int. J. Nanomed. 2014, 17, 4809–4817. [Google Scholar] [CrossRef]
- Grigoriu, C.; Kuroki, Y.; Nicolae, I.; Zhu, X.; Hirai, M.; Suematsu, H.; Takata, M.; Yatsui, K. Photo and cathodoluminescence of Si/SiO2 nanoparticles produced by laser ablation, J. Optoelectron. Adv. Mater. 2005, 7, 2979–2984. [Google Scholar]
- Aebi, H. Catalase in vitro. Meth. Enzymol. 1974, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, F.; Aldinucci, D.; Mocali, A.; Caparrini, A. A sensitive spectrophotometric method for the determination of superoxide dismutase activity in tissue extracts. Anal. Biochem. 1986, 154, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E. Glutathione peroxidase. In Red Cell Metabolism; Beutler, E., Ed.; A Manual of Biochemical Methods; Grune and Stratton: Orlando, FL, USA, 1984; pp. 74–76. [Google Scholar]
- Goldberg, D.M.; Spooner, R.J. Assay of Glutathione Reductase. In Methods of Enzymatic Analysis, 3rd ed.; Bergmeyen, H.V., Ed.; Verlog Chemie, Deerfiled Beach: Hoboken, NJ, USA, 1983; Volume 3, pp. 258–265. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Omari, S.S.; Nasirzadeh, N. 8-Hydroxy-2′-deoxyguanosine (8-OHdG) as a biomarker of oxidative DNA damage induced by occupational exposure to nanomaterials: A systematic review. Nanotoxicology 2021, 15, 850–864. [Google Scholar] [CrossRef] [PubMed]
- Diz, A.P.; Truebano, M.; Skibinski, D.O.F. The consequences of sample pooling in proteomics: An empirical study. Electrophoresis 2009, 30, 2967–2975. [Google Scholar] [CrossRef] [PubMed]
- Laurin, E.; Thakur, K.; Mohr, P.G.; Hick, P.; Crane, M.S.J.; Gardner, I.A.; Moody, N.J.G.; Colling, A.; Ernst, I. To pool or not to pool? Guidelines for pooling samples for use in surveillance testing of infectious diseases in aquatic animals. J. Fish. Dis. 2019, 42, 1471–1491. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. The cell-cycle arrest and apoptotic functions of p53 in tumor initiation and progression. Cold Spring Harb. Perspect. Med. 2016, 6, a026104. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Tooze, S.A. Emerging roles of ATG proteins and membrane lipids in autophagosome formation. Cell Discov. 2020, 6, 32. [Google Scholar] [CrossRef]
- Tran, S.; Fairlie, W.D.; Lee, E.F. BECLIN1: Protein structure, function and regulation. Cells 2021, 10, 1522. [Google Scholar] [CrossRef]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and autophagy. Methods Mol. Biol. 2008, 445, 77–88. [Google Scholar] [CrossRef]
- Pilger, A.; Rudiger, H.W. 8-Hydroxy-2′-deoxyguanosine as a marker of oxidative DNA damage related to occupational and environmental exposures. Int. Arch. Occup. Environ. Health 2006, 80, 1–15. [Google Scholar] [CrossRef]
- Rim, K.T.; Song, S.W.; Kim, H.Y. Oxidative DNA damage from nanoparticle exposure and its application to workers’ health: A literature review. Saf. Health Work 2013, 4, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, M.C.; Ruiter, S.; Lommelaars, T.; Sippel, J.; Hodemaekers, H.M.; van den Brandhof, E.J.; Pennings, J.L.; Kamstra, J.H.; Jelinek, J.; Issa, J.P.; et al. Zebrafish embryos as a screen for DNA methylation modifications after compound exposure. Toxicol. Appl. Pharmacol. 2016, 291, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Guo, Y. Epigenetic clock: DNA methylation in aging. Stem. Cells Int. 2020, 8, 1047896. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Gupta, V.; Zern, B.J.; Pan, D.; Zakrewsky, M.; Muzykantov, V.; Mitragotri, S. Delivering nanoparticles to lungs while avoiding liver and spleen through adsorption on red blood cells. ACS Nano 2013, 7, 11129–11137. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Allan, S.M.; Al-Ahmady, Z.S. Re-directing nanomedicines to the spleen. A potential technology for peripheral immunomodulation. J. Cont. Rel. 2022, 350, 60–79. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox Family NADPH oxidases in mechano-transduction: Mechanisms and consequences. Antioxid. Redox Signal. 2014, 20, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wang, L.; Mettenbrink, E.M.; DeAngelis, P.L.; Wilhelm, S. Nanoparticle Toxicology. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 269–289. [Google Scholar] [CrossRef]
- Yim, M.B.; Chock, P.B.; Stadtman, E.R. Copper, zinc dismutase catalyzes hydroxyl radical production from hydrogen peroxide. Proc Natl. Acad. Sci. USA 1990, 87, 5006–5010. [Google Scholar] [CrossRef]
- Gebicka, L.; Krych-Madej, J. The role of catalases in the prevention/promotion of oxidative stress. J. Inorg. Biochem. 2019, 197, 110699. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochem. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Rad. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Jindal, A.B. Nanocarriers for spleen targeting: Anatomo-physiological considerations, formulation strategies and therapeutic potential. Drug Deliv. Translat. Res. 2016, 6, 473–485. [Google Scholar] [CrossRef]
- Das, K.; Meena, R.; Gaharwar, U.S.; Priyadarshini, E.; Rawat, K.; Paulraj, R.; Mohanta, Y.K.; Saravanan, M.; Bohidar, H.B. Bioaccumulation of CdSe quantum dots show biochemical and oxidative damage in wistar rats. Oxid. Med. Cell Longev. 2023, 6, 7707452. [Google Scholar] [CrossRef]
- Sun, H.; Yang, B.; Cui, E.; Liu, R. Spectroscopic investigations on the effect of N-acetyl-L-cysteine-capped CdTe quantum dots on CAT. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 132, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, M.; Hakimian, S.; Mustafa, T.A.; Aziz, F.M.; Salihi, A.; Ale-Ebrahim, M.; Mirpour, M.; Rasti, B.; Akhtari, K.; Shahpasand, K.; et al. The interaction of silica nanoparticles with catalase and human mesenchymal stem cells: Biophysical, theoretical and cellular studies. Int. J. Nanomed. 2019, 14, 5355–5368. [Google Scholar] [CrossRef] [PubMed]
- Stan, M.S.; Cinteza, L.O.; Petrescu, L.; Mernea, M.A.; Calborean, O.; Mihailescu, D.F.; Sima, C.; Dinischiotu, A. Dynamic analysis of the interactions between Si/SiO2 quantum dots and biomolecules for improving applications based on nano-bio interfaces. Sci. Rep. 2018, 8, 5289. [Google Scholar] [CrossRef] [PubMed]
- Mills-Goodlet, R.; Johnson, L.; Hoppe, I.J.; Regl, C.; Geppert, M.; Schenck, M.; Huber, S.; Hauser, M.; Ferreira, F.; Hűsing, N.; et al. The nanotopography of SiO2 particles impacts the selectivity and 3D fold of bound allergens. Nanoscale 2021, 13, 20508–20520. [Google Scholar] [CrossRef]
- Ngo, V.; Duennwald, M.L. Nrf2 and Oxidative Stress: A General Overview of Mechanisms and Implications in Human Disease. Antioxidants 2022, 11, 2345. [Google Scholar] [CrossRef]
- Harvey, C.J.; Thimmulappa, R.K.; Singh, A.; Blake, D.J.; Ling, G.; Wakabayashi, N.; Fujii, J.; Myers, A.; Biswal, S. Nrf2-regulated glutathione recycling independent of biosynthesis is critical for cell survival during oxidative stress. Free Radic. Biol. Med. 2009, 46, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Zaafan, M.A.; Haridy, A.R.; Abdelhamid, A.M. Amitriptyline attenuates bleomycin-induced pulmonary fibrosis: Modulation of the expression of NF-κβ, iNOS, and Nrf2. Naunyn. Schmiedeberg’s Arch. Pharmacol. 2019, 392, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, H.; Dai, X.; Fang, S.; Wang, X.; Zhang, Y.; Yao, H.; Zhang, X.; Chao, J. p53/PUMA expression in human pulmonary fibroblasts mediates cell activation and migration in silicosis. Sci. Rep. 2015, 5, 16900. [Google Scholar] [CrossRef] [PubMed]
- Tovy, A.; Spiro, A.; McCarthy, R.; Shipony, Z.; Aylon, Y.; Allton, K.; Ainbinder, E.; Furth, N.; Tanay, A.; Barton, M.; et al. p53 is essential for DNA methylation homeostasis in naïve embryonic stem cells, and its loss promotes clonal heterogeneity. Genes Dev. 2017, 31, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Pogribna, M.; Koonce, N.A.; Mathew, A.; Word, B.; Patri, A.K.; Lyn-Cook, B.; Hammons, G. Effect of titanium dioxide nanoparticles on DNA methylation in multiple human cell lines. Nanotoxicology 2020, 14, 534–553. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Z.; Jia, J.; Du, T.; Zhang, N.; Tang, Y.; Fang, Y.; Fang, D. Overview of histone modification. Adv. Exp. Med. Biol. 2021, 1283, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cristian, R.-E.; Balta, C.; Herman, H.; Trica, B.; Sbarcea, B.G.; Hermenean, A.; Dinischiotu, A.; Stan, M.S. In Vivo Assessment of Hepatic and Kidney Toxicity Induced by Silicon Quantum Dots in Mice. Nanomaterials 2024, 14, 457. [Google Scholar] [CrossRef]
- Gao, F.; Ma, N.; Zhou, H.; Wang, Q.; Zhang, H.; Wang, P.; Hou, H.; Wen, H.; Li, L. Zinc oxide nanoparticles-induced epigenetic change and G2/M arrest are associated with apoptosis in human epidermal keratinocytes. Int. J. Nanomed. 2016, 11, 3859–3874. [Google Scholar] [CrossRef]
- Soo Choi, H.; Liu, W.; Misra, P.; Tanaka, E.; Zimmer, J.P.; Itty Ipe, B.; Bawendi, M.G.; Frangioni, J.V. Renal clearance of quantum dots. Nat. Biotechnol. 2007, 25, 1165–1170. [Google Scholar] [CrossRef]
- Decuzzi, P.; Godin, B.; Tanaka, T.; Lee, S.Y.; Chiappini, C.; Liu, X.; Ferrari, M. Size and shape effects in the biodistribution of intravascularly injected particles. J. Control. Release 2010, 141, 320–327. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristian, R.-E.; Balta, C.; Herman, H.; Ciceu, A.; Trica, B.; Sbarcea, B.G.; Miutescu, E.; Hermenean, A.; Dinischiotu, A.; Stan, M.S. Exploring In Vivo Pulmonary and Splenic Toxicity Profiles of Silicon Quantum Dots in Mice. Materials 2024, 17, 2778. https://doi.org/10.3390/ma17112778
Cristian R-E, Balta C, Herman H, Ciceu A, Trica B, Sbarcea BG, Miutescu E, Hermenean A, Dinischiotu A, Stan MS. Exploring In Vivo Pulmonary and Splenic Toxicity Profiles of Silicon Quantum Dots in Mice. Materials. 2024; 17(11):2778. https://doi.org/10.3390/ma17112778
Chicago/Turabian StyleCristian, Roxana-Elena, Cornel Balta, Hildegard Herman, Alina Ciceu, Bogdan Trica, Beatrice G. Sbarcea, Eftimie Miutescu, Anca Hermenean, Anca Dinischiotu, and Miruna S. Stan. 2024. "Exploring In Vivo Pulmonary and Splenic Toxicity Profiles of Silicon Quantum Dots in Mice" Materials 17, no. 11: 2778. https://doi.org/10.3390/ma17112778