
Study on Electrochemical Insulin Sensing Utilizing a DNA Aptamer-Immobilized Gold Electrode

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
2.1. Spectroscopic Activity of IGA3


2.2. Electrochemical Response of the IGA3-Hemin Complex
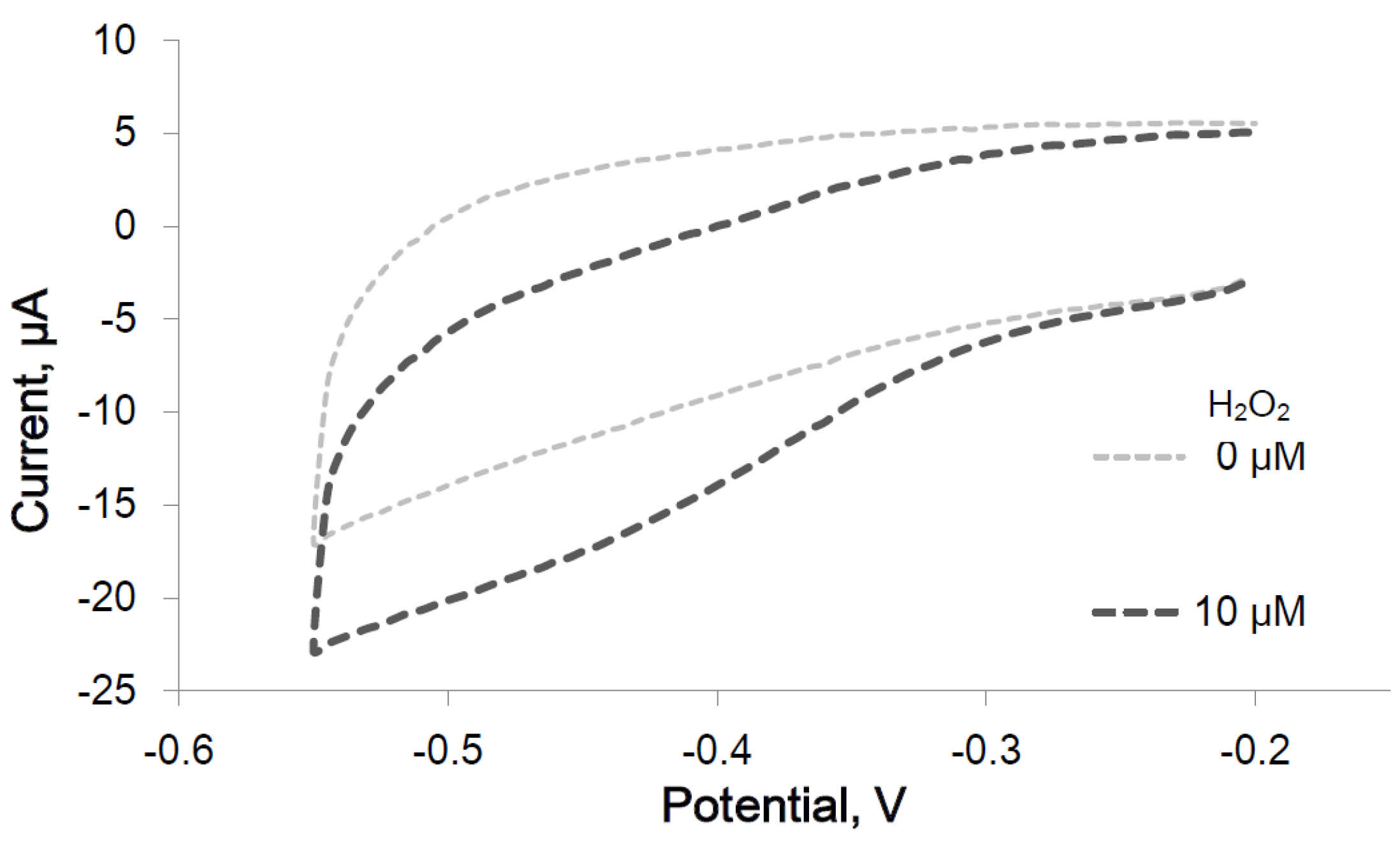
2.2.1. Peroxidase Activity of the Immobilized IGA3-Hemin Complex


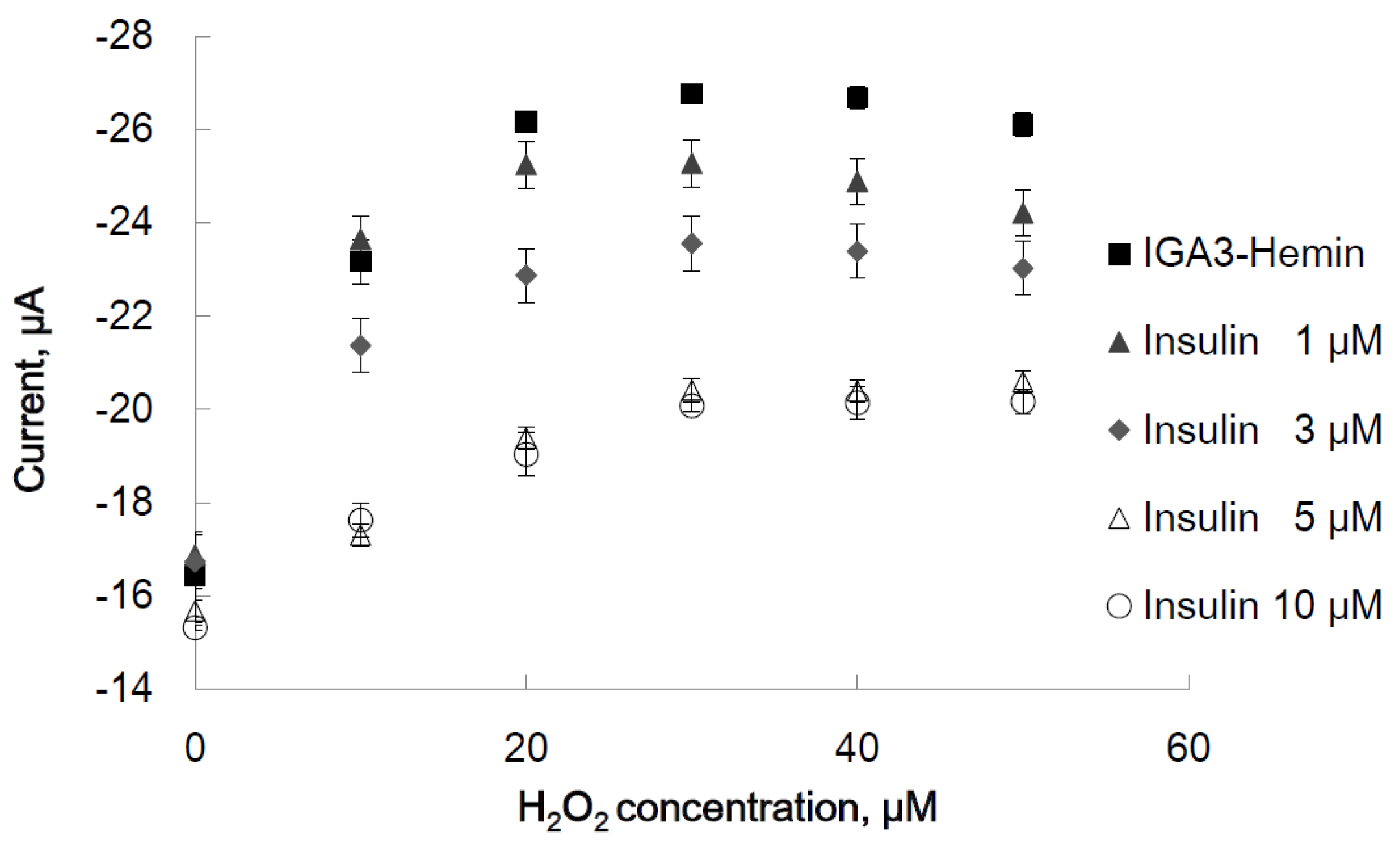
2.2.2. Effect of Insulin on Peroxidase Activity of the IGA3-Hemin Complex

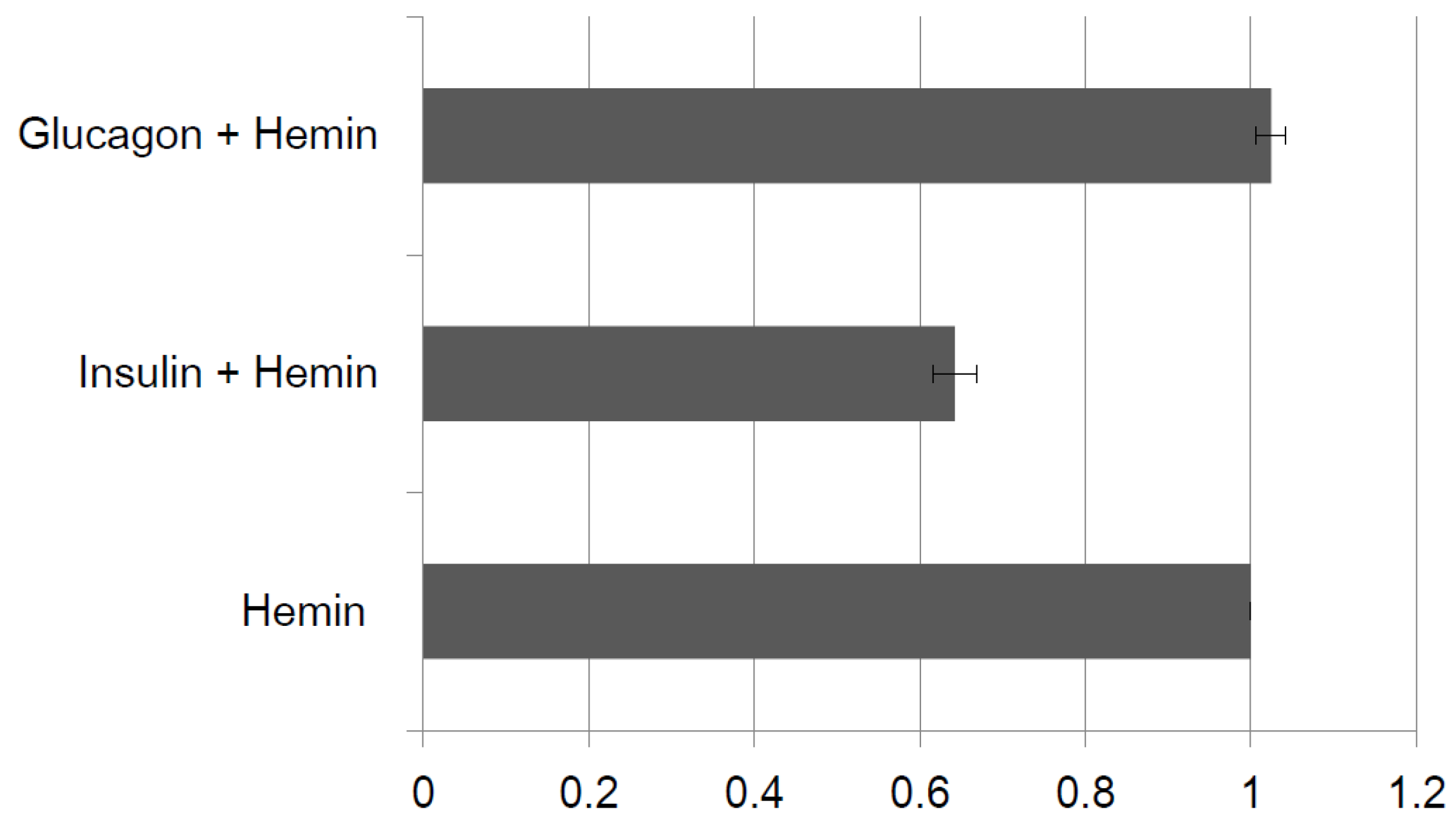
2.2.3. Specific Binding to Insulin

3. Experimental Section
3.1. Materials
3.2. Spectrophotometric Measurement of Peroxidase Activity
3.3. Preparation of Hemin-Binding DNA Aptamer-Modified Electrode
3.4. Electrochemical Measurements
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhihui, P.; Min, L.; Yi, N.; Le, D. Rapid fluorescent detection of escherichia coli K88 based on DNA aptamer library as direct and specific reporter combined with immuno magnetic separation. J. Fluores. 2014, 24, 1159–1168. [Google Scholar]
- Mahlknecht, G.; Seta, M.; Yarden, Y. Aptamer targeting the ERBB2 receptor tyrosine kinase for applications in tumor therapy. Gene Ther. Solid Cancers Method Mol. Biol. 2015, 1317, 3–15. [Google Scholar]
- Bruno, J.G.; Carrillo, M.P.; Richarte, A.M.; Phillips, T.; Andrews, C.; Lee, J.S. Development, screening, and analysis of DNA aptamer libraries potentially useful for diagnosis and passive immunity of arboviruses. BMC Res. Notes 2012, 633. [Google Scholar] [CrossRef]
- Zhao, S.; Yang, W.; Lai, R.Y. A folding-based electrochemical aptasensor for detection of vascular endothelial growth factor in human whole blood. Biosens. Bioelectron. 2011, 26, 2442–2447. [Google Scholar]
- Tokunaga, T.; Namiki, S.; Yamada, K.; Imaishi, T.; Nonaka, H.; Hirose, K.; Sando, S. Cell surface-anchored fluorescent aptamer sensor enables imaging of chemical transmitter dynamics. J. Am. Chem. Soc. 2012, 134, 9561–9564. [Google Scholar]
- Roy, J.; Chirania, P.; Ganguly, S.; Huang, H. A DNA aptamer sensor for 8-oxo-7,8-dihydroguanine. Bioorg. Med. Chem. Lett. 2012, 22, 863–867. [Google Scholar]
- Xie, S.; Yuan, R.; Chai, Y.; Bai, L.; Yuan, Y.; Wang, Y. Label-free electrochemical aptasensor for sensitive thrombin detection using layer-by-layer self-assembled multilayers with toluidine blue-graphene composites and gold nanoparticles. Talanta 2012, 98, 7–13. [Google Scholar]
- Arnott, S.; Hukins, D.W. Optimised parameters for A-DNA and B-DNA. Biochem. Biophys. Res. Commun. 1972, 47, 1504–1509. [Google Scholar] [CrossRef]
- Arnott, S.; Selsing, E. The conformation of C-DNA. J. Mol. Biol. 1975, 98, 265–269. [Google Scholar] [CrossRef]
- Wang, A.H.-J.; Quingley, G.J.; Kolpak, F.J.; van der Marel, G.; van Boon, J.; Rich, H.A. Left-handed double helical DNA: Variations in the backbone conformation. Science 1981, 211, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Watson, J. Molecular Biology of the Gene, 3rd ed.; W.A. Benjamin, Inc.: Menlo Park, CA, USA, 1977. [Google Scholar]
- Gellert, M.; Lipsett, D.M.N.; Davies, R. Helix formation by guanylic acid. Proc. Natl. Acad. Sci. USA 1962, 48, 2013–2018. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.R.; Rachuraman, M.K.; Cech, T.R. Monovalent cation-induced structure of telomeric DNA: The G-quartet model. Cell 1989, 59, 871–880. [Google Scholar] [CrossRef]
- Travascio, P.; Li, Y.; Sen, D. DNA-enhanced peroxidase activity of a DNA-aptamer-hemin complex. Chem. Biol. 1998, 5, 505–517. [Google Scholar] [CrossRef]
- Kubo, I.; Eguchi, T.; Hoshino, Y.; Lin, M.; Abe, H.; Itoh, Y. Peroxidase activity of G-quadruplex hemin-binding DNA aptamers determined by electrochemical measurement. ECS Trans. 2013, 50, 1–7. [Google Scholar] [CrossRef]
- Yoshida, W.; Mochizuki, E.; Takase, M.; Hasegawa, H.; Morita, Y.; Yamazaki, H.; Sode, K.; Ikebukuro, K. Selection of DNA aptamers against insulin and construction of an aptameric enzyme subunit for insulin sensing. Biosens. Bioelectron. 2009, 24, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubo, I.; Eguchi, T. Study on Electrochemical Insulin Sensing Utilizing a DNA Aptamer-Immobilized Gold Electrode. Materials 2015, 8, 4710-4719. https://doi.org/10.3390/ma8084710
Kubo I, Eguchi T. Study on Electrochemical Insulin Sensing Utilizing a DNA Aptamer-Immobilized Gold Electrode. Materials. 2015; 8(8):4710-4719. https://doi.org/10.3390/ma8084710
Chicago/Turabian StyleKubo, Izumi, and Taiga Eguchi. 2015. "Study on Electrochemical Insulin Sensing Utilizing a DNA Aptamer-Immobilized Gold Electrode" Materials 8, no. 8: 4710-4719. https://doi.org/10.3390/ma8084710
APA StyleKubo, I., & Eguchi, T. (2015). Study on Electrochemical Insulin Sensing Utilizing a DNA Aptamer-Immobilized Gold Electrode. Materials, 8(8), 4710-4719. https://doi.org/10.3390/ma8084710