1. Introduction
Chinese fir (
Cunninghamia lanceolata (Lamb) Hook) is one of the major commercial conifer species in China [
1,
2]. It is a typical subtropical coniferous tree in Southern China [
3,
4]. Chinese fir is the most important valued timber species due to its fast growth, high yield, and excellent wood quality (i.e., straight and decay resistant) as well as having significant cultural and historical values to the people [
3]. The plantations of Chinese fir are the most important forests in Southern China, covering an area of over 12 million ha or about 6.5% of the world’s plantation forests [
2]. The plantations area of Chinese fir has been enlarged because of the increasing demand for timber and economic development [
2]. Across China, the plantations of Chinese fir have been utilized for more than one thousand years [
3,
5]. During the last decade, the focus of Chinese fir plantations management has shifted from timber production to ecological functions [
6]. Since the 1980s, decreasing production of Chinese fir plantations has been reported, primarily due to soil degradation [
7,
8]. Currently, the sustainability of Chinese fir plantations is threatened by biodiversity reduction, production loss, soil degradation, and a lack of self-regeneration [
4,
9]. Therefore, understanding the factors that control regeneration, systematic breeding and selection of the best potential trees for future breeding programs are a major research priority for this tree.
Chinese fir has received considerable breeding effort over the past 50 years [
10]. The systematic breeding programs started in 1960s and remarkable successes were achieved [
11]. However, some biological characteristics inherent to genetic breeding program, such as long generation time, highly genetic load, and inbreeding depression have effects on the breeding interventions [
11,
12]. Such effects call for an understanding of pollen grains as these structures are sexual reproductive units and the carriers of male genetic material in Chinese fir. Because of their biological functions, pollen grains play a vital role in the breeding programs of Chinese fir like any other tree species [
13]. In a genetic breeding program, knowledge on pollen viability is an essential factor for artificial hybridization [
14]. Understanding the viability and capacity of pollen germination and pollen tube growth are fundamental for studies of reproductive biology and genetics breeding of plants [
15,
16]. Pollen germination potential has been defined as the ability of viable pollen to germinate under suitable conditions [
17]. Pollen germination and pollen tube growth are important materials for morphological, physiological, biotechnological, ecological, evolutional, biochemical, and molecular biological studies [
15]. Therefore, determination of pollen viability is crucial in tree breeding. The determination involves investigating chemical changes during germination of pollen and growth of pollen tubes through the use of direct methods such as the induction of in vitro germination [
18].
In vitro pollen germination provides a novel approach and strategy to accelerate the genetic improvement of tree breeding. It is a very convenient and effective technique for studying many basic and applied aspects of pollen biology [
19]. This technique is the best option for selecting viable and potential pollen that can be used for cross-pollination [
20]. The conditions required for in vitro pollen germination vary across species. It has been reported that pollen germination and pollen tube growth can be influenced by many internal and external factors [
21,
22]. The extrinsic factors which affect the germination of pollen include incubation time, optimal temperature, and medium composition [
23]. Organic and inorganic substances such as sucrose, boric acid, calcium nitrate, potassium nitrate, and magnesium sulfate exert an effect on the in vitro pollen germination [
24,
25,
26]. In addition, pH has influences on in vitro pollen germination [
16,
27,
28]. Indeed, the quality of pollen, the optimum concentrations of the media and environmental conditions have effects on germination of pollen and tube growth [
20].
Various methods and media with different components have been suggested by many plant biologists and researchers [
19,
23,
29,
30,
31]. Recently, there are a growing number of studies carried out in Chinese fir. The majority of these studies have focused on flowering characteristics and genetic variation [
4,
10,
32,
33,
34,
35]. On the other hand, Shen et al. [
36], investigated the effect of sucrose, boric acid, and calcium ions on pollen germination. Yanfang et al. [
37], studied pollen germination and preservation characteristics under different storage conditions. Fragallah et al. [
38], studied metabolomic analysis during pollen germination and tube growth. Despite these examples, a more detailed study about the influence of sucrose, boric acid concentrations, pH, and optimal incubation time is not yet reported. Therefore, this study aimed to investigate the effect of sucrose, boric acid, pH, and time of incubation on germination and pollen tube growth of Chinese fir pollen grains. This study is valuable for providing information about pollen germination requirements to support further improvement of artificial pollination and breeding programs of Chinese fir.
2. Materials and Methods
2.1. Description of the Study Area
The study was carried out in a third-generation clonal seed orchard of Chinese fir belonging to state-owned forest farm established in 2008 at YouXi County in Fujian province, China (latitude 25°50′–26°26′ N, longitude 117°48′–118°39′ E). The total area of the orchard is 188 acres divided into 13 districts. The trees population consists of 70 clones originating from superior plus-trees (breeding parents). The grafts were collected from Yangkou forest farm germplasm resources to produce high genetic quality seeds. Grafts were planted at a spacing of 4 m × 4 m, in the different districts of the orchard. The climate of the area is a subtropical monsoon. The annual temperature ranges from 15.8–19.6 °C. The annual rainfall ranges from 1400–1488 mm; the relative humidity is 83%, annual solar radiation 95–102 Kcal/cm2, annual sunshine hours are 1764.6 TJ. The wind is southeast in summer and northwest in winter; wind speed is 1.1–1.7 m/s.
2.2. Collection of Pollen Samples
Male cones were collected during the pollination period from the end of February to mid-March 2017. The pollen grains were collected from mature male cones after ripening. The male cones were hung upside down in open plastic bags, labeled separately. In addition, samples were taken from clones (CL-4, 7, 10, 11, 17, 25, 35, 44, and 51). After the collection of the pollen for all clones, the pollen samples were placed in plastic bags. Then, fresh pollen grains were shed, poured into 1.5 mL centrifuge tubes, and sealed with parafilm. The tubes were put into silica gel and kept at 4 °C until use.
2.3. In Vitro Pollen Germination
In preliminary experiments, in vitro pollen germination tests were performed on pollen of 30 different clones in a basic media. Briefly, the basic media consisted of 5% sucrose, 0.5% agar, 100 mL distilled water, and pH adjusted to 5.8 by using a digital pH-meter after adjusting the solution using HCl and NaOH. All media components were dissolved in boiling water and poured into petri dishes. Then, pollen grains were distributed on the surface of the cooling media. For the germination test, the petri dishes were inoculated in a growth chamber for 12 h in light and 12 h in dark cycles. The relative humidity and temperature conditions set for inoculation were 70% and 25 °C, respectively, as previously described by Fragallah et al. [
38].
After establishing the basic media for the 30 clones, 9 clones (CL-4, 7, 10, 11, 17, 25, 35, 44, and 51) were chosen based on their pollen germination ability. Clones (CL-4, 17, and 25) had germination rates above 80%. Clones (CL-11, 44, and 35) had germination rates above 65 and less than 80%. Clones (CL-7, 10, and 51) had germination rates less than 65% (non-published data). Furthermore, the basic media was considered a control and coded as (A) type media. Then, a total of 9 different culture media with different concentrations of sucrose (0, 10 and 15%) coded as (B, C, and D type media), boric acid (0.01, 0.1, and 0.2%) coded as (E, F, and G type media), and levels of pH (4.5, 5, and 7) coded as (H, I, and J type media) were prepared to study the effects of the media concentrations on pollen germination and tube growth (
Table 1).
The experiment was carried out in a completely randomized design with four replications, each represented by one petri dish at each time intervals at 1, 12, 24, and 48 h. Then, the petri dishes were placed under light microscope and photographed with Nikon, SMZ 127 (version 4.50) under 8× magnifications. A pollen grain was considered germinated when the pollen tube length was at least equal to or greater than the grain diameter [
39]. All pollen grains in each photo were counted to calculate germination percentages. Germinated and non-germinated pollen was counted from each petri dish after 1, 12, 24, and 48 h of incubation. Pollen germination was determined and the percentages were recorded by dividing the number of germinated pollen grains per field of view by the total number of pollen per field of view and expressed as percentages, as previously described by Kakani et al. [
39].
2.4. Pollen Tube Growth Measurements
The pollen tube growth was measured by using Image Pro-Plus 6.0 imaging software system for analysis and processing photos. The mean of pollen tube growth (µm) was calculated as the average lengths of 20 pollen tubes randomly measured from each photo at 1, 12, 24, and 48 h intervals.
2.5. Statistical Analysis
The data of pollen germination and pollen tube growth were input into Co-Stat software. Then, the replicated average values of pollen germination and tube growth data were subjected to analysis of variance (ANOVA) to determine the significance of measured parameters. Duncan multiple range (DMR) test was used for mean separation. Furthermore, simple linear regression analysis model was developed. A linear regression model attempted to explain the relationship between two or more variables using a straight line. The dependent variable was referred to as the response. The slope and the intercept of the line are called regression coefficients. The equation used was Y = a + bx; Where (Y) is the dependent variable, (X) is the independent variable (i.e., it is plotted on the X axis), (b) is the slope of the line, and (a) is the y-intercept.
3. Results
3.1. Effects of Sucrose Concentrations on Pollen Germination and Tube Growth
Table 2 and
Table 3 show mean values (± standard error (SE)) of the effect of sucrose concentration on pollen germination and tube growth after 48 h of incubation. The effect was significant and most obvious at sucrose concentrations of 10 and 15%. Sucrose concentrations of 0.0 and 5% (control media) had a fewer effects on pollen germination rates and tube growth.
The maximum pollen germination rate was obtained from CL-17 (90.29%), followed by CL-4 (89.56%) in D type media for 15% sucrose, while CL-51 recorded the minimum germination rate (58.50%). In C type media for sucrose concentration 10%, the germination rate was maximum in CL-17 (90.16%) followed by CL-25 (87.57). The minimum (61.48%) was recorded in CL-51. In B type media for sucrose concentration 0%, the germination rate was higher in CL-4 (81.49%) followed by CL-17 (80.82%). The minimum germination rate was (44.16%) for CL-51. In A type media for 5% sucrose, CL-4 showed the maximum rate of pollen germination (84.89%) followed by CL-17 (84.09%), while CL-10 (52.92%) and CL-7 (51.42%) provided the minimum germination rates.
The maximum average of pollen tube growth after 48 h of incubation was obtained by CL-17 (0.78 µm) followed by CL-4 (0.76 µm) in D type media for sucrose 15%. The minimum rate was observed in CL-51 (0.41 µm). In C type media for sucrose concentration 10% the maximum growth was 0.73 µm by CL-17 followed by CL-4 (0.66 µm). In B type media for 0%, the maximum pollen tube growth was observed in CL-4 (0.45 µm) followed by CL-17 (0.44 µm), while the minimum tube growth recorded by CL-7 (0.32 µm). In A type media for 5% sucrose, the maximum tube growth was observed for CL-4 (0.53 µm) followed by CL-17 (0.52 µm), while CL-7 recorded the minimum tube length of 0.34 µm.
3.2. Effects of Boric Acid Concentration on Pollen Germination and Tube Growth
Table 4 and
Table 5 show mean values (± SE) of the effect of boric acid concentrations on pollen germination and tube growth after 48 h of incubation. Boric acid significantly promoted the germination of pollen and tube growth. Promotion of germination was most notable at boric acid concentration of 0.01%. Pollen germination rate and tube growth increased significantly compared with that in the boric acid-deficient media (control media). Boric acid completely inhibited pollen germination and tube growth at 0.2%.
The maximum pollen germination rate was recorded in CL-17 (92.97%) followed with CL-4 (83.34%) in E type media for 0.01% boric concentration. The minimum rate was found in CL-51 (64.48%). In F type media with 0.1% boric concentration, CL-4 gave the highest germination rate of 85.70%, followed by CL-17 (85.16%) and the lowest was recorded by CL-7 (50.34%). In G type media with 0.2% boric acid concentration, the highest germination rate of 83.31% reported by CL-4 followed by CL-17 (83.31%) while the lowest germination rate was obtained from CL-7 (35.91%). In boric acid-deficient A type media, CL-4 showed the maximum rate of pollen germination of 84.89% followed by CL-17 (84.09%), while CL-10 (52.92%) and CL-7 (51.42%) had the lowest germination rates.
In E type medium with 0.01% boric acid, the maximum average pollen tube growth was recorded by CL-17 (0.91 µm), followed by CL-4 (0.69 µm) while the minimum tube growth was recorded in CL-51 (0.45 µm). In F type medium with 0.1% boric acid, the maximum pollen tube growth was obtained from CL-17 (0.59 µm) followed by CL-4 (0.47 µm), and the minimum was recorded in CL-7 (0.34 µm). In G type medium with 0.2% boric acid, the maximum pollen tube growth was obtained from CL-4 (0.47 µm) followed by CL-17 (0.40 µm), and the minimum was recorded CL-51 (0.32 µm). In boric acid-deficient A type media, the maximum tube growth was reported by CL-4 (0.53 µm) followed by CL-17 (0.52 µm), while CL-7 recorded the minimum tube length of 0.34 µm.
3.3. Effects of Different pH Levels on Pollen Germination and Tube Growth
Table 6 and
Table 7 show mean values (±SE) of the effects of pH levels on pollen germination rates and tube growth after 48 h of incubation. The effect of pH level on pollen germination and tube growth was significant. The culture media with a level of pH 7.0 and 5.0 presented better effects on pollen germination and tube growth than culture media of pH 4.5 and 5.8 pH.
The maximum pollen germination rate was reported by CL-4 (88.89%) followed with CL-4 (88.56%) in J type media for the level of pH 7.0. The minimum rate was 57.31% reported in CL-51. In I type media at pH 5.0, CL-4 gave the highest germination rate of 87.64% followed by CL-17 (88.56%) and the lowest was recorded by CL-51 (55.04%). In H type media at pH 4.5, the highest germination rate was 80.82% reported by CL-4 followed by CL-17 (79.82%), while the lowest germination rate was obtained from CL-44 (40.51%). In A type media at pH 5.8, CL-4 showed the maximum rate of pollen germination (84.89%) followed by CL-17 (84.09%), while CL-10 (52.92%) and CL-7 (51.42%) provided the minimum germination rates.
CL-4 recorded the maximum average of tube growth at 0.78 µm in J type media at pH 7.0 followed by CL-17 (0.74 µm), while the minimum was recorded in CL-51 and CL-44 (0.40 µm). In I type media at pH 5.0, the maximum pollen tube growth was obtained from CL-17 (0.72 µm) followed by CL-4 (0.61 µm). The minimum tube growth was observed in CL-44 (0.40 µm). In H type media at pH 4.5, the maximum pollen tube growth was obtained from CL-17 and CL-4 (0.46 µm), while the minimum was recorded by CL-44 (0.36 µm). In A type media at pH 5.8, the maximum tube growth was reported by CL-4 (0.53 µm) followed by CL-17 (0.52 µm), while CL-7 recorded the minimum tube length (0.34 µm).
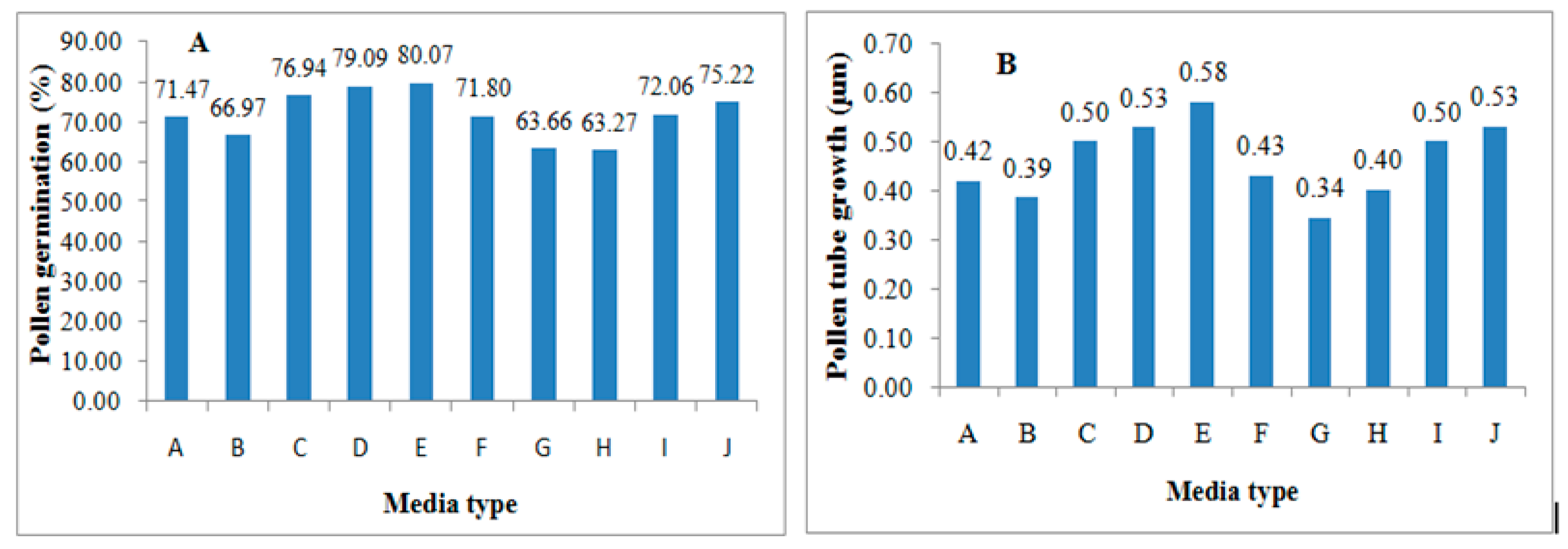
3.4. The Overall Effect of Media Types on Pollen Germination and Tube Growth
Table 8 means (±SE) and
Figure 1 show the effects of different media types on pollen germination and tube growth. On the overall basis, the C and D type media containing 10% and 15% sucrose performed a bit well regarding germination and pollen tube growth as compared to A and B type media for sucrose concentrations 5 and 0%.
For boric acid media concentrations, E type media for 0.01% was the best among other concentrations. The G type media for the 0.2% concentration performed poorly regarding the germination and tube growth. For media of different pH, it was quite remarkable that J type media at pH 7.0 was best among the culture media. On the contrary, H type media at pH 4.5 had poor performance.
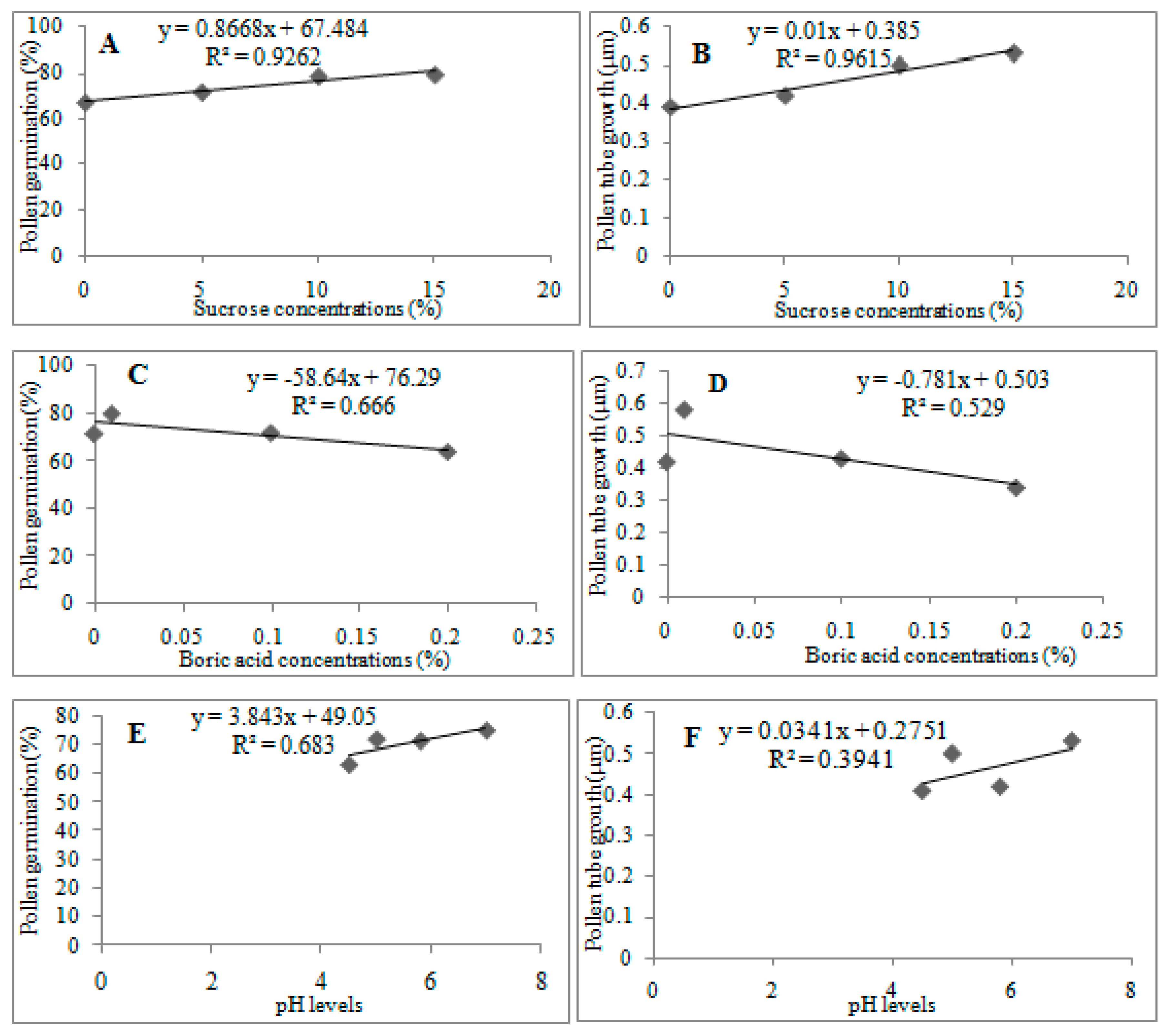
3.5. Relationships between Sucrose, Boric Acid, and pH and Pollen Germination and Tube Growth
The results in
Figure 2 show the relationships between sucrose, boric acid, and pH and pollen germination and tube growth: Sucrose concentrations, pollen germination, and tube growth showed strong and positive relationship (A,B), boric acid concentrations, pollen germination, and tube growth, showed strong and negative relationship (C,D), strong and positive relationship between pH levels and pollen germination and tube growth was observed (E,F).
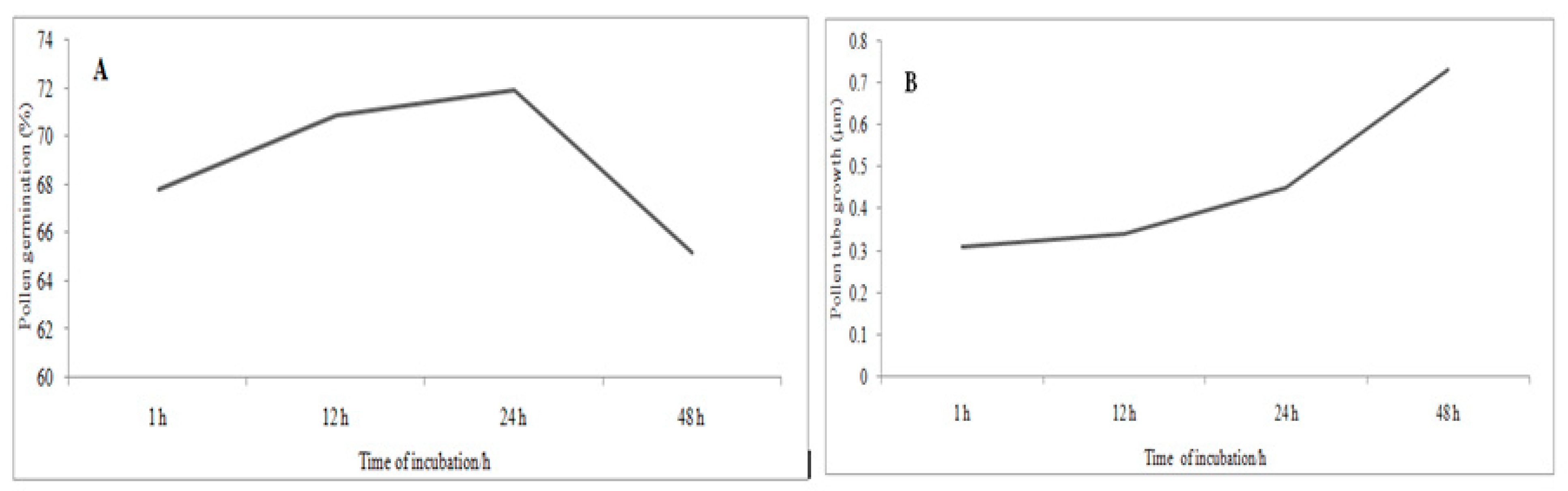
3.6. The status of Pollen Germination and Tube Growth at Different Time of Incubation
Figure 3 shows the status of pollen germination and tube growth at different times of incubation. Pollen grains began to germinate immediately after being placed in the media. After 1 h, germination rates increased and reached the maximum at 24 h, with a significant decrease at 48 h. On the other hand, pollen tube growth starts to elongate at 24 h and dramatically increased over time and reached the maximum growth at 48 h of incubation.
3.7. Effect of Incubation Time on Pollen Germination and Tube Growth
Table 9 show the effect of incubation time on in vitro pollen germination and tube growth. Based on overall mean of all nine clones, pollen germination in the first hour of incubation was 67.79%, and this growth increased to 70.83% when the incubation time increased to 12 h. Thereafter, up to 24 h, the germination rates slightly increased to 71.88%, and then severely decreased to 65.15% at 48 h. A similar trend was noticed, as the incubation time increased the growth of pollen tubes increased. The pollen tube growth at 1 h of incubation was 0.31 µm, and then rose to 0.34 µm when the time increased to 12 h. At 24 h, the pollen tube growth increased to 0.45 µm. The maximum tube growth 0.73 µm was achieved at the end of 48 h of incubation (
Figure 4).
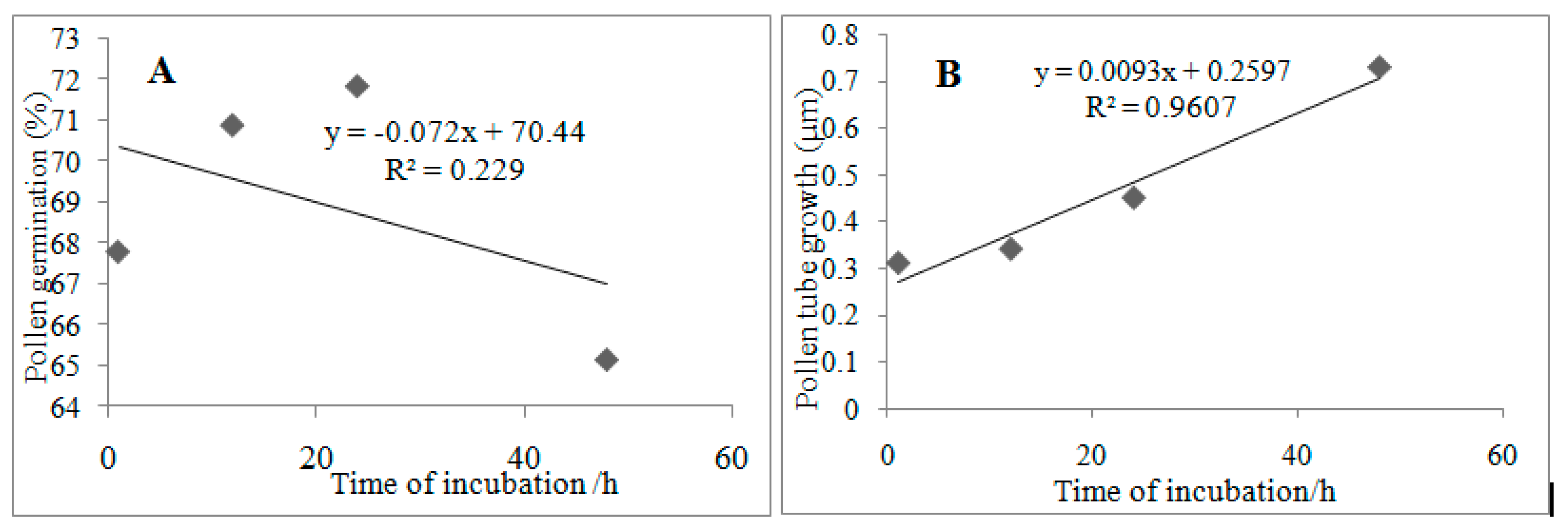
3.8. Relationships between the Time of Incubation, Pollen Germination, and Tube Growth
Figure 5 shows the relationships between the time of incubation, pollen germination, and tube growth. The time of incubation and pollen germination showed negative relationship
A, while the time of incubation and pollen germination showed strong and positive relationship
B.
4. Discussion
Understanding the requirements for pollen germination and tube growth is of great importance for cross-pollination. The requirements of pollen germination vary appreciably from pollen grains of one species to another. In general, each pollen grain has its own requirements and might need different media. Various germination media tests and culture conditions may affect the pollen germination of a given species. The media preparations for in vitro germination play a vital role in pollen germination and tube growth [
15]. In general, the media preparation for pollen germination varies according to the plant species and many internal and external factors [
15]. Thus, there are a number of factors which have been reported to influence in vitro germination of pollen grains. For example, humidity, temperature, genotypic differences, vigor and physiological stage of the plant and the age of the flower and ingredients of the substrate used for germination have an effect on pollen germination [
40]. Sahar and Spiegel-Roy, [
41] reported that some nutrients (e.g., calcium, magnesium sulphate, potassium nitrate, and boric acid) influenced pollen germination. In this study, various combinations of culture media were tested for the provision of optimum germination conditions required by the pollen of Chinese fir. However, to study the effects of different medium concentrations on pollen germination and tube growth, in vitro pollen germination was performed in different concentrations of sucrose separately and in combination with varying boric acid concentration and pH. Based on the results obtained from the basic media, the germination percentages varied across the studied clones. There were significant differences on germination percentage and pollen tube growth at
p < 0.05 level. The reason for such differences might be attributed to the variation in the genetic background of the studied clones.
4.1. Effects of Sucrose Concentration on Pollen Germination and Tube Growth
The supplementation of sucrose in culture media is vital because it serves as the main nutrient for the germination and formation of pollen tubes [
23]. Sucrose at an appropriate concentration plays a dominant role in both pollen germination and pollen tube growth. A suitable concentration of sucrose can favor both germination and pollen tube development, whereas high concentrations of sucrose in a media can inhibit both processes [
23]. The optimum sugar concentration for pollen germination differs considerably among different plants and species [
31]. The differences in pollen germination within the same culture media might be attributed to an imbalanced osmotic pressure of the culture media [
42]. The results of this study revealed that there are significant differences in pollen germination and tube growth among sucrose concentrations. In general, pollen germination and tube growth increased with an increase of the sucrose concentration. The maximum germination and tube growth of pollen were obtained in media of 15 and 10% sucrose concentrations (
Table 2 and
Table 3). Furthermore, strong positive relationships were found between sucrose concentrations and pollen germination and tube growth. This relationship indicates that with increase in sucrose concentrations, the pollen germination and tube growth increase as well (
Figure 2). The possible reason for the positive relationships may be related to the supply of metabolic energy and the biosynthesis of organic compounds necessary for cell growth, agreeing with [
27,
43,
44]. On the other hand, these results disagree with [
36,
45], probably because of the differences on the concentrations and genetic variation of clones used.
4.2. Effects of Boric Acid Concentrations on Pollen Germination and Tube Growth
Boric acid is believed to promote pollen germination and tube growth [
29]. Boric acid is essential to germination and tube growth of pollen [
44]. An absence of boric acid normally affects pollen germination, and this has been confirmed in many plant species [
46]. In this study, the results showed that boric acid promoted pollen germination and tube growth. Boric acid in a concentration of 0.01% significantly increased pollen germination rates and tube growth compared with that in the boric acid-deficient media. Pollen germination and tube growth were severely and completely inhibited by 0.2% boric acid (
Table 4 and
Table 5). In addition, strong negative relationships were found between boric acid concentration and pollen germination and tube growth. This implies that the germination and growth of pollen considerably improved through the addition of boric acid at appropriate concentrations. If the optimal concentration of boric acid is not used or if higher concentrations are added, pollen germination and tube growth would be hindered (
Figure 2). This could be the reason for the significant increase in germination rate and tube growth of pollen in lower concentrations. Another explanation for these relationships could be related to the formation of the ionisable sugar–borate complex, which reacts with the plasma membrane, promoting greater growth of the pollen tube. These findings agree with those confirmed by [
29,
36,
47,
48], who noted that only low concentrations of boric acid stimulated pollen germination and pollen tube growth, whereas higher concentrations inhibited germination and tube growth.
4.3. Effects of Different pH Levels on Pollen Germination and Tube Growth
The pH of the media for in vitro germination is an important factor controlling pollen germination and pollen tube development in different plant species. The importance of determination of the ideal pH in the physiological processes that involve pollen grains is linked to the resulting higher germination percentage [
16]. Ramos et al. [
49], stated that there is an increase in the germination of pollen with an increase in the pH of the culture media. Boavida & McCormick, [
27] reported that pH has a significant effect on pollen germination and tube growth. Burke et al. [
50] tested the effect of pH on in vitro pollen germination and elongation of the pollen tube, and stated no differences between the tested pH values. Fan et al. [
51], tested several components and reported pH as an important factor contributing to pollen germination success and for pollen tube growth. In our study, the tested pH levels (4.5, 5.0, 5.8, and 7.0) had positive effects on pollen germination and tube growth. Indeed, we found that pollen germination and tube growth improved drastically when the pH levels were increased to pH 7.0 (
Table 6 and
Table 7). The interpretation of this positive relationship is that, as the pH level increases, the germination and pollen tube growth increased, agreeing with the results of [
27,
28,
49].
4.4. Effect of Incubation Time on Pollen Germination and Tube Growth
Optimum time of incubation is one of the basic requirements of pollen germination and tube growth [
43]. A prolonged incubation period is necessary to overcome dormancy. Many researchers have studied the effect of incubation time on in vitro pollen germination and tube growth [
23,
38,
43]. Lin et al. [
23], reported that incubation time played a vital role in pollen germination and pollen tube growth of oil palm. In this study, our results revealed that pollen germination rate and elongation of pollen tube increased with increase of incubation time (
Figure 4). The increase suggests that a certain degree of dormancy may be present in some viable pollen. The other possible assumption is that some of the pollen may have reached physiological maturity and a longer incubation period is not required for the pollen to germinate, and the prolonged time of incubation is only required for pollen tube elongation. Our data confirm the results reported by [
23,
38,
51,
52], who proved that there is an increase in germination percentages of pollen grains during the time of incubation.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}