The Effects of DNA Methylation Inhibition on Flower Development in the Dioecious Plant Salix Viminalis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Treatments
2.2. Genomic DNA Extraction and Methylation-Sensitive Amplified Polymorphism Analysis
2.3. Phytohormone Extraction and Measurement
2.4. Carbohydrate Measurement
2.5. Data Analysis
3. Results
3.1. DNA Methylation during the Flower Transition and Flower Development Stages
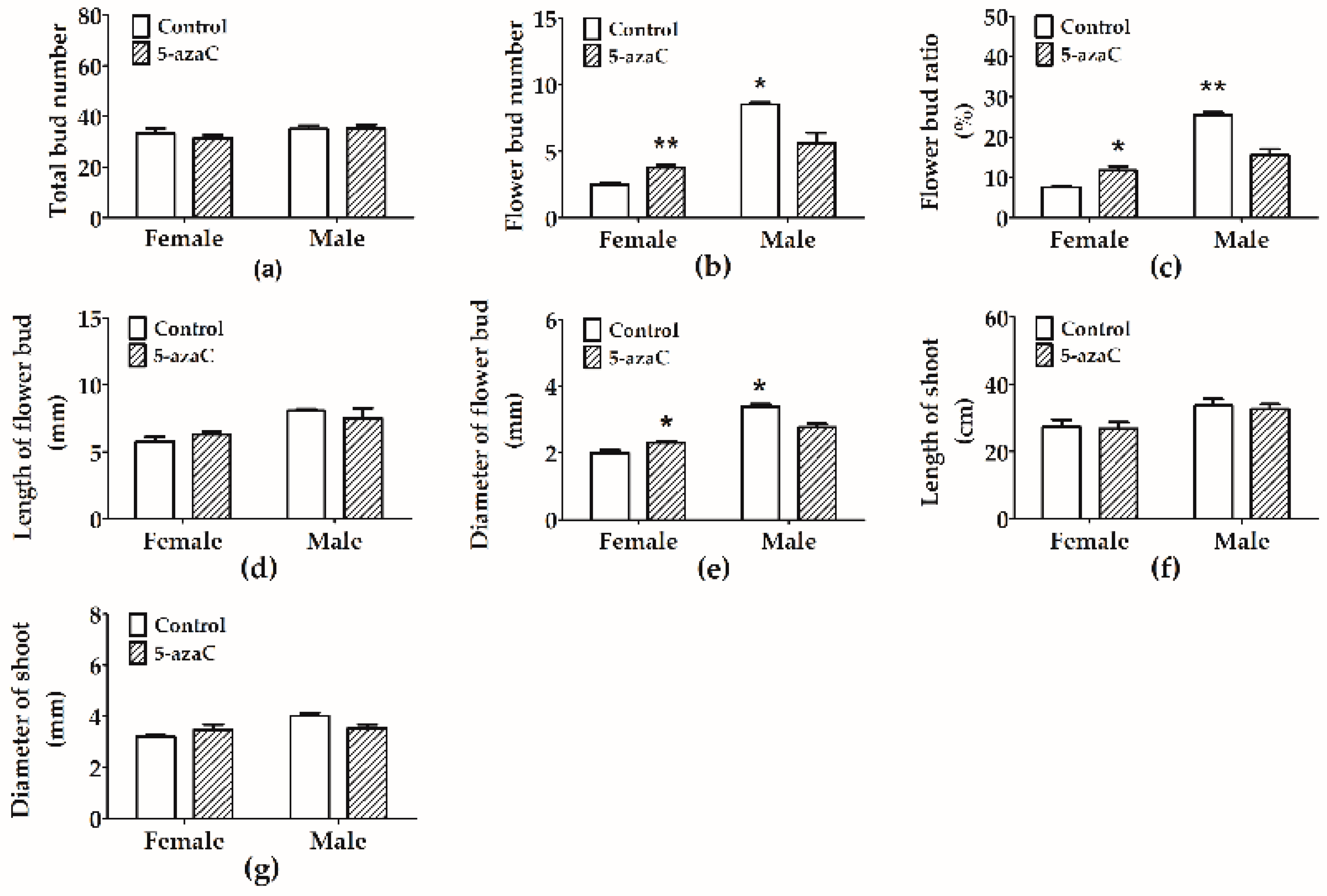
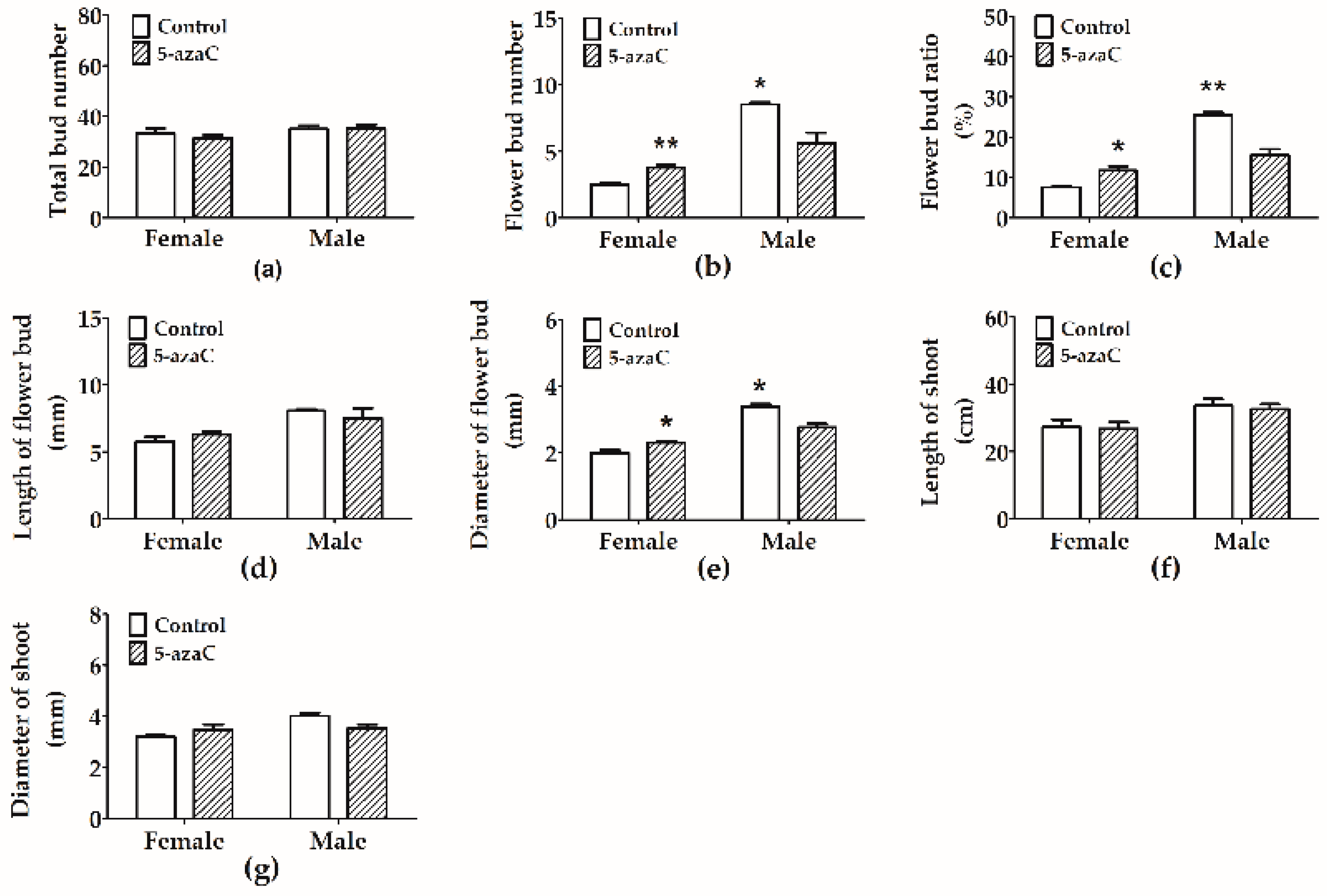
3.2. Effects of 5-Azac on Flower Bud Development
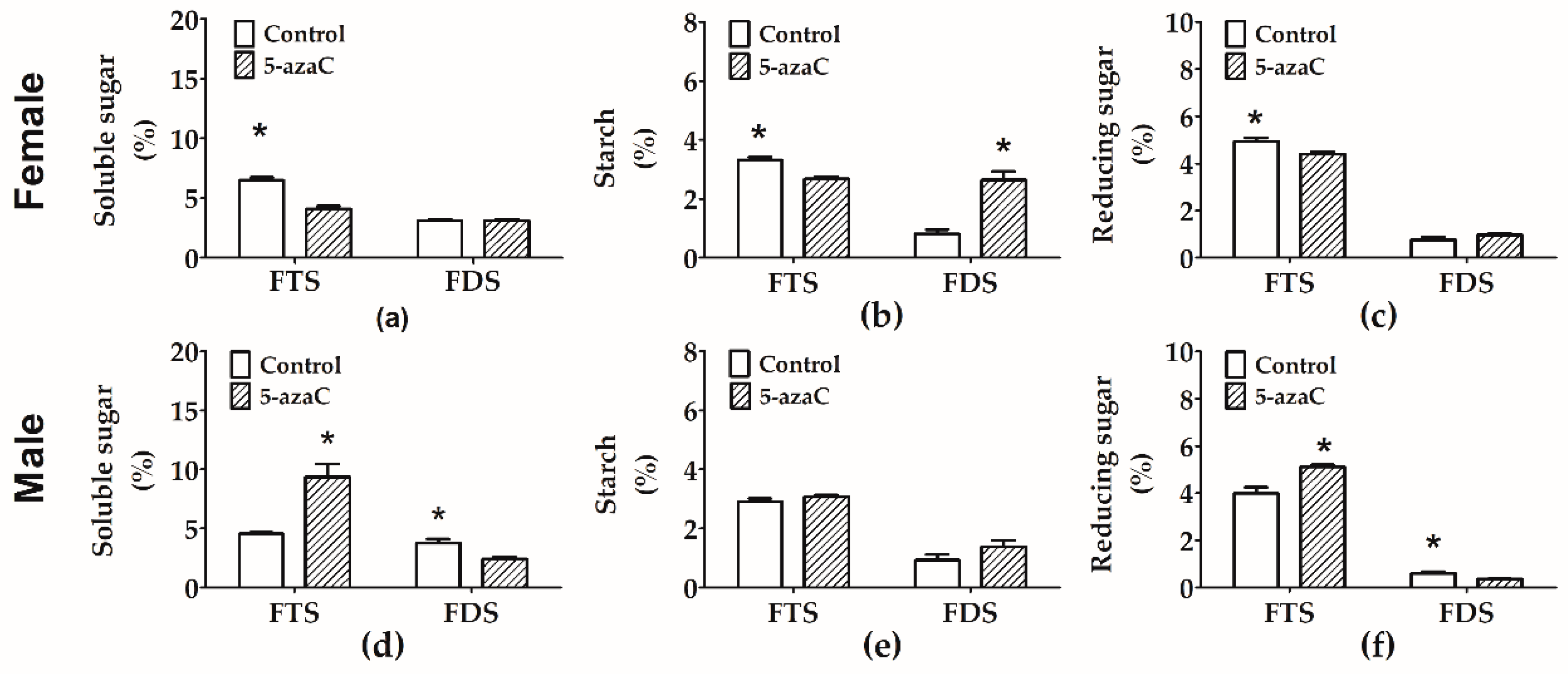
3.3. Effects of 5-Azac on Carbohydrate Contents
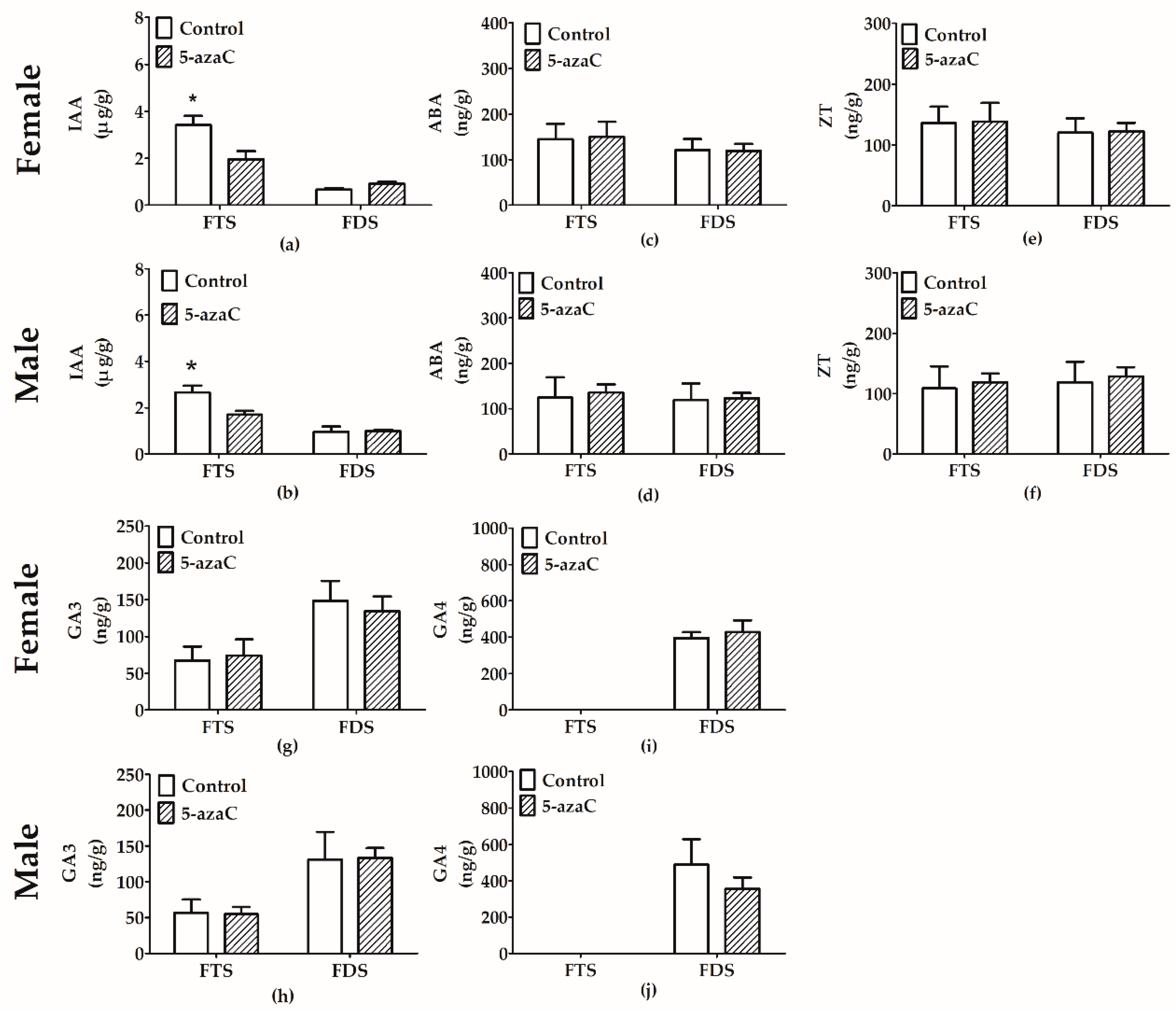
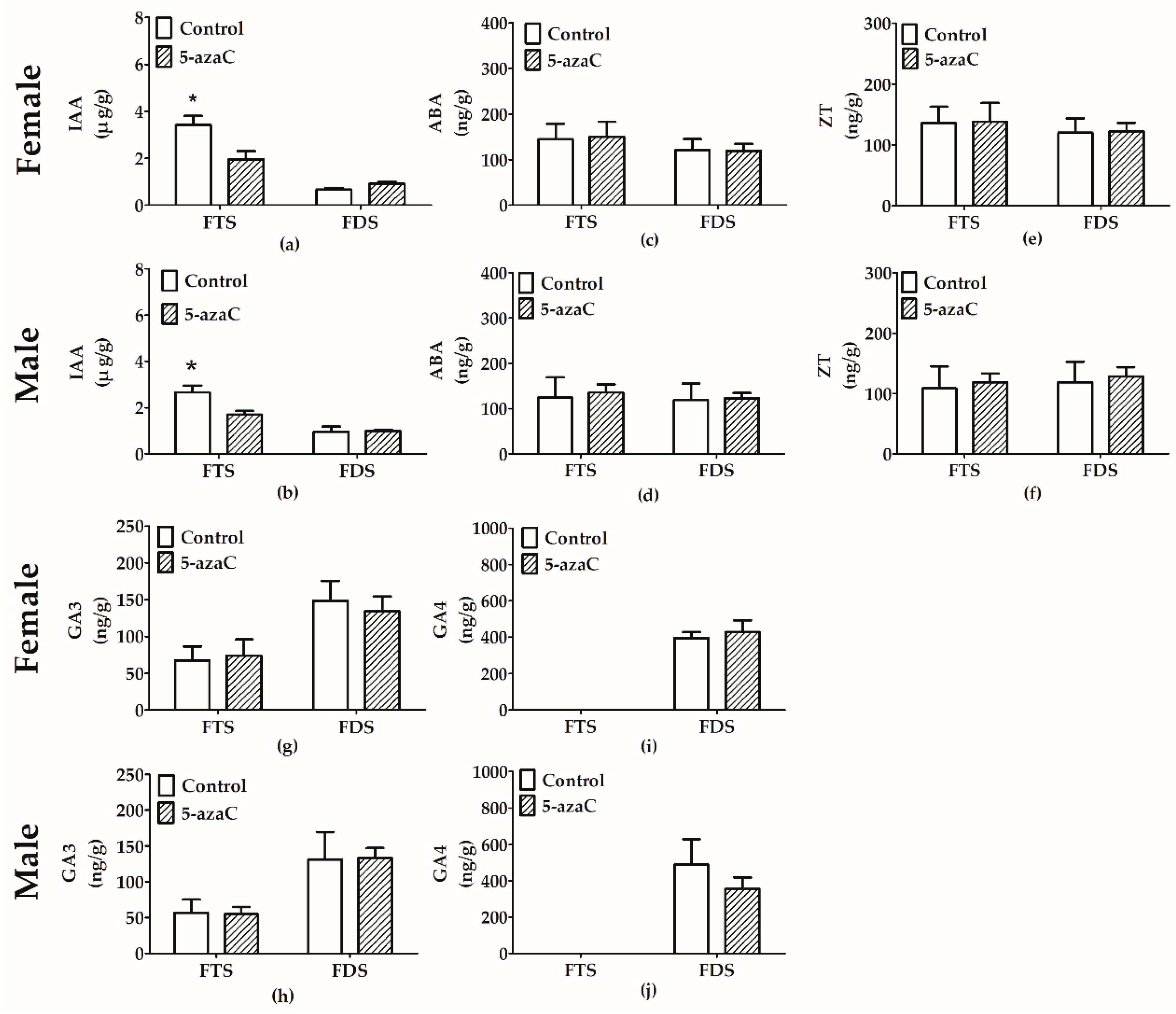
3.4. Effect of 5-Azac on Phytohormone Levels
4. Discussion
4.1. DNA Methylation and Flower Development
4.2. The Relationship between Endogenous Carbohydrates, Hormones and Floral Development
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yamaguchi, N.; Winter, C.M.; Wu, M.F.; Kanno, Y.; Yamaguchi, A.; Seo, M.; Wagner, D. Gibberellin acts positively then negatively to control onset of flower formation in Arabidopsis. Science 2014, 344, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, J.D.; Sedgley, M.; Olesen, T. Regulation of floral initiation in horticultural trees. J. Exp. Bot. 2008, 59, 3215–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, M.; Honsho, C.; Kanzaki, S.; Shimizu, K.; Utsunomiya, N. Isolation and expression analysis of flowering locus t-like and gibberellin metabolism genes in biennial-bearing mango trees. Sci. Hortic. 2012, 139, 108–117. [Google Scholar] [CrossRef]
- Southwick, S.M.; Glozer, K. Reducing flowering with gibberellins to increase fruit size in stone fruit trees: Applications and implications in fruit production. Horttechnology 2000, 109, 744–751. [Google Scholar] [CrossRef]
- Xing, L.B.; Zhang, D.; Li, Y.M.; Shen, Y.W.; Zhao, C.P.; Ma, J.J.; An, N.; Han, M.Y. Transcription profiles reveal sugar and hormone signaling pathways mediating flower induction in apple (Malus domestica borkh.). Plant Cell Physiol. 2015, 56, 2052–2068. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.C.; Koenig, D.; Chitwood, D.H.; Sinha, N.R. A sister of pin1 gene in tomato (Solanum lycopersicum) defines leaf and flower organ initiation patterns by maintaining epidermal auxin flux. Dev. Biol. 2016, 419, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Godin, C.; Azpeitia, E.; Farcot, E. An Introduction to Modelling Flower Primordium Initiation. Available online: https://hal.inria.fr/hal-01254880 (accessed on 25 January 2019).
- Heisler, M.G.; Ohno, C.; Das, P.; Sieber, P.; Reddy, G.V.; Long, J.A.; Meyerowitz, E.M. Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the arabidopsis inflorescence meristem. Curr. Biol. 2005, 15, 1899–1911. [Google Scholar] [CrossRef] [PubMed]
- Corbesier, L.; Lejeune, P.; Bernier, G. The role of carbohydrates in the induction of flowering in Arabidopsis thaliana: Comparison between the wild type and a starchless mutant. Planta 1998, 206, 131–137. [Google Scholar] [CrossRef]
- Bernier, G.; Perilleux, C. A physiological overview of the genetics of flowering time control. Plant Biotechnol. J. 2005, 3, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Araya, T.; Noguchi, K.; Terashima, I. Effect of nitrogen nutrition on the carbohydrate repression of photosynthesis in leaves of Phaseolus vulgaris L. J. Plant Res. 2010, 123, 371–379. [Google Scholar] [CrossRef]
- Dielen, V.; Lecouvet, V.; Dupont, S.; Kinet, J.M. In vitro control of floral transition in tomato (Lycopersicon esculentum mill.), the model for autonomously flowering plants, using the late flowering uniflora mutant. J. Exp. Bot. 2001, 52, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ma, F.; Shu, H.; Han, M. Branch bending affected floral bud development and nutrient accumulation in shoot terminals of ‘fuji’ and ‘gala’ apples. Acta Physiol. Plant. 2017, 39. [Google Scholar] [CrossRef]
- Yang, H.; Chang, F.; You, C.; Cui, J.; Zhu, G.; Wang, L.; Zheng, Y.; Qi, J.; Ma, H. Whole-genome DNA methylation patterns and complex associations with gene structure and expression during flower development in arabidopsis. Plant J. 2015, 81, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Soppe, W.J.; Jacobsen, S.E.; Alonso-Blanco, C.; Jackson, J.P.; Kakutani, T.; Koornneef, M.; Peeters, A.J. The late flowering phenotype of FWA mutants is caused by gain-of-function epigenetic alleles of a homeodomain gene. Mol. Cell 2000, 6, 791–802. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, Z.; Zhu, J.-K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ma, Z.; Zhang, Z.; Cheng, L.; Zhang, X.; Li, T. Small RNA-sequencing links physiological changes and rddm process to vegetative-to-floral transition in apple. Front. Plant Sci. 2017, 8, 873. [Google Scholar] [CrossRef]
- Campos-Rivero, G.; Osorio-Montalvo, P.; Sanchez-Borges, R.; Us-Camas, R.; Duarte-Ake, F.; De-la-Pena, C. Plant hormone signaling in flowering: An epigenetic point of view. J. Plant Physiol. 2017, 214, 16–27. [Google Scholar] [CrossRef]
- Jaligot, E.; Rival, A.; Beulé, T.; Dussert, S.; Verdeil, J.L. Somaclonal variation in oil palm (Elaeis guineensis jacq.): The DNA methylation hypothesis. Plant Cell Rep. 2000, 19, 684–690. [Google Scholar] [CrossRef]
- Jaligot, E.; Adler, S.; Debladis, É.; Beulé, T.; Richaud, F.; Ilbert, P.; Finnegan, E.J.; Rival, A. Epigenetic imbalance and the floral developmental abnormality of the in vitro-regenerated oil palm Elaeis guineensis. Ann. Bot. 2011, 108, 1453–1462. [Google Scholar] [CrossRef]
- Eeuwens, C.J.; Lord, S.; Donough, C.R.; Rao, V.; Vallejo, G.; Nelson, S. Effects of tissue culture conditions during embryoid multiplication on the incidence of “mantled” flowering in clonally propagated oil palm. Plant Cell Tissue Organ Cult. 2002, 70, 311–323. [Google Scholar] [CrossRef]
- Li, C.H.; Yu, N.; Jiang, S.M.; Shangguan, X.X.; Wang, L.J.; Chen, X.Y. Down-regulation of s-adenosyl-l-homocysteine hydrolase reveals a role of cytokinin in promoting transmethylation reactions. Planta 2008, 228, 125–136. [Google Scholar] [CrossRef]
- Meijon, M.; Canal, M.J.; Valledor, L.; Rodriguez, R.; Feito, I. Epigenetic and physiological effects of gibberellin inhibitors and chemical pruners on the floral transition of azalea. Physiol. Plant. 2011, 141, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Blázquez, M.A.; Soowal, L.N.; Lee, I.; Weigel, D. Leafy expression and flower initiation in Arabidopsis. Development 1997, 124, 3835–3844. [Google Scholar]
- Zhang, J.Z.; Mei, L.; Liu, R.; Khan, M.R.; Hu, C.G. Possible involvement of locus-specific methylation on expression regulation of leafy homologous gene (cilfy) during precocious trifoliate orange phase change process. PLoS ONE 2014, 9, e88558. [Google Scholar] [CrossRef]
- Stresemann, C.; Brueckner, B.; Musch, T.; Stopper, H.; Lyko, F. Functional diversity of DNA methyltransferase inhibitors in human cancer cell lines. Cancer Res. 2006, 66, 2794–2800. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Shiraya, T.; Wada, K.C.; Takeno, K. Induction of flowering by DNA demethylation in Perilla frutescens and Silene armeria: Heritability of 5-azacytidine-induced effects and alteration of the DNA methylation state by photoperiodic conditions. Plant Sci. 2010, 178, 321–326. [Google Scholar] [CrossRef]
- Burn, J.E.; Bagnall, D.J.; Metzger, J.D.; Dennis, E.S.; Peacock, W.J. DNA methylation, vernalization and the initiation of flowering. Proc. Natl. Acad. Sci. USA 1993, 90, 287–291. [Google Scholar] [CrossRef]
- Kondo, H.; Ozaki, H.; Itoh, K.; Kato, A.; Takeno, K. Flowering induced by 5-azacytidine, a DNA demethylating reagent in a short-day plant, perilla frutescens var. Crispa. Physiol. Plant. 2006, 127, 130–137. [Google Scholar] [CrossRef]
- Kondo, H.; Miura, T.; Wada, K.C.; Takeno, K. Induction of flowering by 5-azacytidine in some plant species: Relationship between the stability of photoperiodically induced flowering and flower-inducing effect of DNA demethylation. Physiol. Plant. 2007, 131, 462–469. [Google Scholar] [CrossRef]
- Janousek, B.; Siroky, J.; Vyskot, B. Epigenetic control of sexual phenotype in a dioecious plant, Melandrium album. Mol. Gen. Genet. 1996, 250, 483–490. [Google Scholar] [CrossRef]
- Li, S.F.; Zhang, G.J.; Yuan, J.H.; Deng, C.L.; Lu, L.D.; Gao, W.J. Effect of 5-azac on the growth, flowering time and sexual phenotype of spinach. Russ. J. Plant Physiol. 2015, 62, 670–675. [Google Scholar] [CrossRef]
- Foster, T.; Johnston, R.; Seleznyova, A. A morphological and quantitative characterization of early floral development in apple (Malus x domestica borkh.). Ann. Bot. 2003, 92, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Heide, O.M.; Prestrud, A.K. Low temperature but not photoperiod, controls growth cessation and dormancy induction and release in apple and pear. Tree Physiol. 2005, 25, 109–114. [Google Scholar] [CrossRef]
- Kurokura, T.; Mimida, N.; Battey, N.H.; Hytonen, T. The regulation of seasonal flowering in the Rosaceae. J. Exp. Bot. 2013, 64, 4131–4141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Ma, K.; Ci, D.; Chen, Q.; Tian, J.; Zhang, D. Sexual dimorphic floral development in dioecious plants revealed by transcriptome, phytohormone and DNA methylation analysis in Populus tomentosa. Plant Mol. Biol. 2013, 83, 559–576. [Google Scholar] [CrossRef]
- Delph, L.F.; Gehring, J.L.; Arntz, A.M.; Levri, M.; Frey, F.M. Genetic correlations with floral display lead to sexual dimorphism in the cost of reproduction. Am. Nat. 2005, 166 (Suppl. 4), S31–S41. [Google Scholar] [CrossRef]
- Thomas, S.C.; LaFrankie, J.V. Sex, size and interyear variation in flowering among dioecious trees of the Malayan raom forest. Ecology 1993, 74, 1529–1537. [Google Scholar] [CrossRef]
- Reynalópez, G.E.; Simpson, J.; Ruizherrera, J. Differences in DNA methylation patterns are detectable during the dimorphic transition of fungi by amplification of restriction polymorphisms. Mol. Gen. Genet. 1997, 253, 703. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M. Aflp: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407. [Google Scholar] [CrossRef]
- Chen, X.Q.; Yi, M.; Fang, C.; Song, W.Q.; Lei, Z. Analysis of DNA methylation patterns of PLBS derived from Cymbidium hybridium based on MSAP. Plant Cell Tissue Organ. Cult. 2009, 98, 67–77. [Google Scholar] [CrossRef]
- Chakrabarty, D.; Yu, K.W.; Paek, K.Y. Detection of DNA methylation changes during somatic embryogenesis of siberian ginseng (Eleuterococcus senticosus). Plant Sci. 2003, 165, 61–68. [Google Scholar] [CrossRef]
- Tang, X.M.; Tao, X.; Wang, Y.; Ma, D.W.; Li, D.; Yang, H.; Ma, X.R. Analysis of DNA methylation of perennial ryegrass under drought using the methylation-sensitive amplification polymorphism (MSAP) technique. Mol. Genet. Genom. 2014, 289, 1075–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, X.; Wang, X.; Yang, G.; Wu, Y.; Su, S.; Li, S.; Liu, H.; Yuan, Y. Analysis of the DNA methylation of maize (Zea mays L.) in response to cold stress based on methylation-sensitive amplified polymorphisms. J. Plant Biol. 2013, 56, 32–38. [Google Scholar] [CrossRef]
- Wang, Y.; Rong, H.; Xie, T.; Jiang, J.; Wu, J.; Wang, Y. Comparison of DNA methylation in the developing seeds of yellow- and black-seeded Brassica napus through MSAP analysis. Euphytica 2016, 209, 157–169. [Google Scholar] [CrossRef]
- Baurens, F.C.; Nicolleau, J.; Legavre, T.; Verdeil, J.L.; Monteuuis, O. Genomic DNA methylation of juvenile and mature Acacia mangium micropropagated in vitro with reference to leaf morphology as a phase change marker. Tree Physiol. 2004, 24, 401–407. [Google Scholar] [CrossRef]
- Diao, S.; Wang, Y.; Ding, C.; Chang, Y.; Liang, L.; Gao, Y.; Zhang, B.; Su, X. No consistent daily variation in DNA methylation detected in Populus nigra leaves by methylation-sensitive amplification polymorphism analysis. J. For. Res. 2016, 28, 653–660. [Google Scholar] [CrossRef]
- Berlin, S.; Trybush, S.O.; Fogelqvist, J.; Gyllenstrand, N.; Hallingbäck, H.R.; Åhman, I.; Nordh, N.E.; Shield, I.; Powers, S.J.; Weih, M. Genetic diversity, population structure and phenotypic variation in european Salix viminalis L. (salicaceae). Tree Genet. Genomes 2014, 10, 1595–1610. [Google Scholar] [CrossRef]
- Mleczek, M.; Rutkowski, P.; Rissmann, I.; Kaczmarek, Z.; Golinski, P.; Szentner, K.; Strażyńska, K.; Stachowiak, A. Biomass productivity and phytoremediation potential of Salix alba and Salix viminalis. Biomass Bioenergy 2010, 34, 1410–1418. [Google Scholar] [CrossRef]
- Junttila, O. Flower bud differentiation in salix pentandra as affected by photoperiod, temperature and growth regulators. Physiol. Plant. 1980, 49, 127–134. [Google Scholar] [CrossRef]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a ctab DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef]
- Wen, X.; Luo, K.; Xiao, S.; Ai, N.; Wang, S.; Fan, X. Qualitative analysis of chemical constituents in traditional Chinese medicine analogous formula cheng-qi decoctions by liquid chromatography-mass spectrometry. Biomed. Chromatogr. 2016, 30, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Clegg, K.M. The application of the anthrone reagent to the estimation of starch in cereals. J. Sci. Food Agric. 1956, 7, 40–44. [Google Scholar] [CrossRef]
- Lindsay, H. A colorimetric estimation of reducing sugars in potatoes with 3,5-dinitrosalicylic acid. Potato Res. 1973, 16, 176–179. [Google Scholar] [CrossRef]
- Eriksson, S.; Bohlenius, H.; Moritz, T.; Nilsson, O. Ga4 is the active gibberellin in the regulation of leafy transcription and Arabidopsis floral initiation. Plant Cell 2006, 18, 2172–2181. [Google Scholar] [CrossRef]
- Jean Finnegan, E.; Kovac, K.A.; Jaligot, E.; Sheldon, C.C.; James Peacock, W.; Dennis, E.S. The downregulation of flowering locus c (flc) expression in plants with low levels of DNA methylation and by vernalization occurs by distinct mechanisms. Plant J. 2005, 44, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chen, X.; Zhang, H.; Ding, Y. Genome-wide analysis of DNA methylation in photoperiod- and thermo-sensitive male sterile rice peiai 64s. BMC Genom. 2015, 16, 102. [Google Scholar] [CrossRef]
- Fraga, M.F.; Canal, M.J.; Rodriguez, R. Phase-change related epigenetic and physiological changes in Pinus radiata d. Don. Planta 2002, 215, 672–678. [Google Scholar] [CrossRef]
- Hasbún, R.; Valledor, L.; Berdasco, M.; Santamaria, E.; Cañal, M.J.; Rodriguez, R.; Rios, D.; Sánchez, M. In vitro proliferation and genome DNA methylation in adult chestnuts. Acta Hortic. 2005, 693, 333–340. [Google Scholar] [CrossRef]
- Hasbún, R.; Valledor, L.; Santamaría, E.; Cañal, M.J.; Rodríguez, R.; Berdasco, M. Dynamics of DNA methylation in chestnut trees development. Acta Hortic. 2007, 760, 563–566. [Google Scholar] [CrossRef]
- Wassenegger, M. RNA-directed de novo methylation of genomic sequences in plants. Cell 1994, 76, 567–576. [Google Scholar] [CrossRef]
- Lewsey, M.G.; Hardcastle, T.J.; Melnyk, C.W.; Molnar, A.; Valli, A.; Urich, M.A.; Nery, J.R.; Baulcombe, D.C.; Ecker, J.R. Mobile small RNAS regulate genome-wide DNA methylation. Proc. Natl. Acad. Sci. USA 2016, 113, E801–E810. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Qi, F.; Yan, C.; Zhan, Y. Sexually different morphological, physiological and molecular responses of fraxinus mandshurica flowers to floral development and chilling stress. Plant Physiol. Biochem. 2016, 99, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Tian, M.; Ci, D.; Zhang, D. Methylation of microrna genes regulates gene expression in bisexual flower development in andromonoecious poplar. J. Exp. Bot. 2015, 66, 1891–1905. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Huang, J.; Yang, Y.; Yao, Y.A. Transcriptome analysis of flower sex differentiation in Jatropha curcas L. Using RNA sequencing. PLoS ONE 2016, 11, e0145613. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Xu, X.; Zhang, Y.; Korpelainen, H.; Li, C. Nitrogen deposition limits photosynthetic response to elevated co2 differentially in a dioecious species. Oecologia 2011, 165, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, G.; Zang, R.; Korpelainen, H.; Berninger, F. Sex-related differences in leaf morphological and physiological responses in Hippophae rhamnoides along an altitudinal gradient. Tree Physiol. 2007, 27, 399–406. [Google Scholar] [CrossRef]
- Dawson, T.E.; Bliss, L.C. Patterns of water use and the tissue water relations in the dioecious shrub, Salix arctica: The physiological basis for habitat partitioning between the sexes. Oecologia 1989, 79, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Bai, X.; Yang, W.; Wang, W.; Chen, Z.; Ma, J.; Ma, T. Single-base-resolution methylomes of Populus euphratica reveal the association between DNA methylation and salt stress. Tree Genet. Genomes 2018, 14. [Google Scholar] [CrossRef]
- Akagi, T.; Henry, I.M.; Tao, R.; Comai, L. Plant genetics. A y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 2014, 346, 646–650. [Google Scholar] [CrossRef]
- Matsoukas, I.G.; Massiah, A.J.; Thomas, B. Starch metabolism and antiflorigenic signals modulate the juvenile-to-adult phase transition in Arabidopsis. Plant Cell Environ. 2013, 36, 1802–1811. [Google Scholar] [CrossRef] [PubMed]
- Shalom, L.; Samuels, S.; Zur, N.; Shlizerman, L.; Zemach, H.; Weissberg, M.; Ophir, R.; Blumwald, E.; Sadka, A. Alternate bearing in citrus: Changes in the expression of flowering control genes and in global gene expression in on-versus off-crop trees. PLoS ONE 2012, 7, e46930. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.Q.; Zhang, Y.J.; Zhou, S.R.; Hu, W.Y.; Wu, X.T.; Ye, Y.J.; Wu, X.X.; Xiao, Y.P.; Li, X.; Xue, H.W. Global analysis reveals the crucial roles of DNA methylation during rice seed development. Plant Physiol. 2015, 168, 1417–1432. [Google Scholar] [CrossRef] [PubMed]
- Reinders, J.; Wulff, B.B.; Mirouze, M.; Mari-Ordonez, A.; Dapp, M.; Rozhon, W.; Bucher, E.; Theiler, G.; Paszkowski, J. Compromised stability of DNA methylation and transposon immobilization in mosaic Arabidopsis epigenomes. Genes Dev. 2009, 23, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Q.; Gu, J.; Lv, Y. Vernalization of oriental hybrid lily ‘sorbonne’: Changes in physiology metabolic activity and molecular mechanism. Mol. Biol. Rep. 2014, 41, 6619–6634. [Google Scholar] [CrossRef]
- Lai, Y.S.; Zhang, X.; Zhang, W.; Shen, D.; Wang, H.; Xia, Y.; Qiu, Y.; Song, J.; Wang, C.; Li, X. The association of changes in DNA methylation with temperature-dependent sex determination in cucumber. J. Exp. Bot. 2017, 68, 2899–2912. [Google Scholar] [CrossRef] [PubMed]
- Mutasagöttgens, E.; Hedden, P. Gibberellin as a factor in floral regulatory networks. J. Exp. Bot. 2009, 60, 1979–1989. [Google Scholar] [CrossRef] [Green Version]
- King, R.W.; Moritz, T.; Evans, L.T.; Martin, J.; Andersen, C.H.; Blundell, C.; Kardailsky, I.; Chandler, P.M. Regulation of flowering in the long-day grass Lolium temulentum by gibberellins and the flowering locus t gene. Plant Physiol. 2006, 141, 498–507. [Google Scholar] [CrossRef]
- GarcíA-Pallas, I.; Val, J.; Blanco, A. The inhibition of flower bud differentiation in ‘crimson gold’ nectarine with ga 3 as an alternative to hand thinning. Sci. Hortic. 2001, 90, 265–278. [Google Scholar] [CrossRef]
- Krizek, B.A. Auxin regulation of arabidopsis flower development involves members of the aintegumenta-like/plethora (ail/plt) family. J. Exp. Bot. 2011, 62, 3311–3319. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhao, Y. A role for auxin in flower development. J. Integr. Plant Biol. 2007, 49, 99–104. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Wu, M.F.; Winter, C.M.; Berns, M.C.; Nole-Wilson, S.; Yamaguchi, A.; Coupland, G.; Krizek, B.A.; Wagner, D. A molecular framework for auxin-mediated initiation of flower primordia. Dev. Cell 2013, 24, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, Y.; Duan, B.; Korpelainen, H.; Li, C. Sex-related adaptive responses of populus cathayana to photoperiod transitions. Plant Cell Environ. 2009, 32, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Dong, T.; Huang, K.; Yang, Y.; Li, D.; Xu, X.; He, X. Sex-specific floral morphology, biomass and phytohormones associated with altitude in dioecious Populus cathayana populations. Ecol. Evol. 2017, 7, 3976–3986. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EcoRI Primer Sequences (5′–3′) | HpaII/MspI Primer Sequence (5′–3′) | ||
|---|---|---|---|
| Adapter | CTCGTAGACTGCGTACC | Adapter | GACGATGAGTCTCGAT |
| AATTGGTACGCAGTCTAC | CGATCGAGACTCAT | ||
| E-pre | GACTGCGTACCAATTCA | HM-pre | ATGAGTCTCGATCGG |
| E1 | GACTGCGTACCAATTCAGG | HM1 | ATGAGTCTCGATCGGAAT |
| E2 | GACTGCGTACCAATTCACG | HM2 | ATGAGTCTCGATCGGACG |
| E3 | GACTGCGTACCAATTCAAC | HM3 | ATGAGTCTCGATCGGATC |
| E4 | GACTGCGTACCAATTCACA | HM4 | ATGAGTCTCGATCGGATT |
| E6 | GACTGCGTACCAATTCACT | HM6 | ATGAGTCTCGATCGGTCA |
| E7 | GACTGCGTACCAATTCAAG | HM7 | ATGAGTCTCGATCGGTGC |
| HM8 | ATGAGTCTCGATCGGTCCA | ||
| Stage | Sex | Material | Type I | Type II | Type III | Type IV | Total Loci |
|---|---|---|---|---|---|---|---|
| HPA+/MSP+ | HPA+/MSP- | HPA-/MSP+ | HPA-/MSP- | ||||
| FTS | Female | Control | 412 ± 9b | 78 ± 4b | 157 ± 7b | 177 ± 7c | 825 |
| 5-azaC | 423 ± 9b | 75 ± 4bc | 151 ± 4b | 185 ± 8c | 825 | ||
| Male | Control | 440 ± 8b | 58 ± 7c | 153 ± 3b | 184 ± 1c | 835 | |
| 5-azaC | 432 ± 10b | 58 ± 2c | 158 ± 5b | 187 ± 4c | 835 | ||
| FDS | Female | Control | 437 ± 19b | 118 ± 4a | 195 ± 16a | 391 ± 4a | 1142 |
| 5-azaC | 513 ± 11a | 104 ± 7a | 156 ± 11b | 369 ± 2b | 1142 | ||
| Male | Control | 452 ± 14b | 115 ± 11a | 206 ± 12a | 387 ± 13a | 1160 | |
| 5-azaC | 532 ± 18a | 108 ± 5a | 193 ± 6a | 327 ± 11b | 1160 |
| Stage | Sex | Material | Unmethylated (%) | Hemimethylated (%) | Fully Methylated (%) | Total Methylated (%) |
|---|---|---|---|---|---|---|
| FTS | Female | Control | 49.98 ± 1.05a | 9.49 ± 0.44ab | 40.53 ± 0.89c | 50.02 ± 1.05c |
| 5-azaC | 51.23 ± 1.93a | 8.08 ± 0.53bc | 40.69 ± 1.39c | 48.77 ± 1.11c | ||
| Male | Control | 52.73 ± 0.93a | 6.99 ± 0.84c | 40.28 ± 0.33c | 47.27 ± 0.93c | |
| 5-azaC | 51.78 ± 1.18a | 6.99 ± 0.21c | 41.24 ± 0.98c | 48.22 ± 1.18c | ||
| FDS | Female | Control | 38.30 ± 1.67c | 10.36 ± 0.36a | 51.34 ± 1.75a | 61.70 ± 1.67a |
| 5-azaC | 44.92 ± 0.96b | 9.11 ± 0.61ab | 45.97 ± 0.89b | 55.08 ± 0.96b | ||
| Male | Control | 38.94 ± 1.21c | 9.91 ± 0.98ab | 51.15 ± 0.49a | 61.06 ± 1.21a | |
| 5-azaC | 45.83 ± 1.54b | 9.34 ± 0.42ab | 44.83 ± 1.21b | 54.17 ± 1.54b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.-H.; Peng, X.-Y.; Yu, Y.-C.; Sun, Z.-Y.; Han, L. The Effects of DNA Methylation Inhibition on Flower Development in the Dioecious Plant Salix Viminalis. Forests 2019, 10, 173. https://doi.org/10.3390/f10020173
Cheng Y-H, Peng X-Y, Yu Y-C, Sun Z-Y, Han L. The Effects of DNA Methylation Inhibition on Flower Development in the Dioecious Plant Salix Viminalis. Forests. 2019; 10(2):173. https://doi.org/10.3390/f10020173
Chicago/Turabian StyleCheng, Yun-He, Xiang-Yong Peng, Yong-Chang Yu, Zhen-Yuan Sun, and Lei Han. 2019. "The Effects of DNA Methylation Inhibition on Flower Development in the Dioecious Plant Salix Viminalis" Forests 10, no. 2: 173. https://doi.org/10.3390/f10020173
APA StyleCheng, Y. -H., Peng, X. -Y., Yu, Y. -C., Sun, Z. -Y., & Han, L. (2019). The Effects of DNA Methylation Inhibition on Flower Development in the Dioecious Plant Salix Viminalis. Forests, 10(2), 173. https://doi.org/10.3390/f10020173