Litter Traits of Native and Non-Native Tropical Trees Influence Soil Carbon Dynamics in Timber Plantations in Panama

 ,
,
Abstract
:1. Introduction
- Differences in soil microbial biomass and soil respiration among plantations and litter types will be related to soil properties and litter decomposition rates.
- Leaf traits related to resource quality for microbial communities (such as lignin: nitrogen ratio and C content) will explain the variation in decomposition among species and litter mixtures.
- As a result of trait complementarity, litter mixtures will have higher decay rates than expected, based on the decay rates of individual constituent species.
- As plant inputs represent the main substrate for decomposers and as microbial communities adapt to available resources, native litter decomposes faster under the species of origin (homefield advantage), whereas the decomposition of all litter types is slower in teak plantations.
2. Materials and Methods
2.1. Study Site and Litter Mixtures
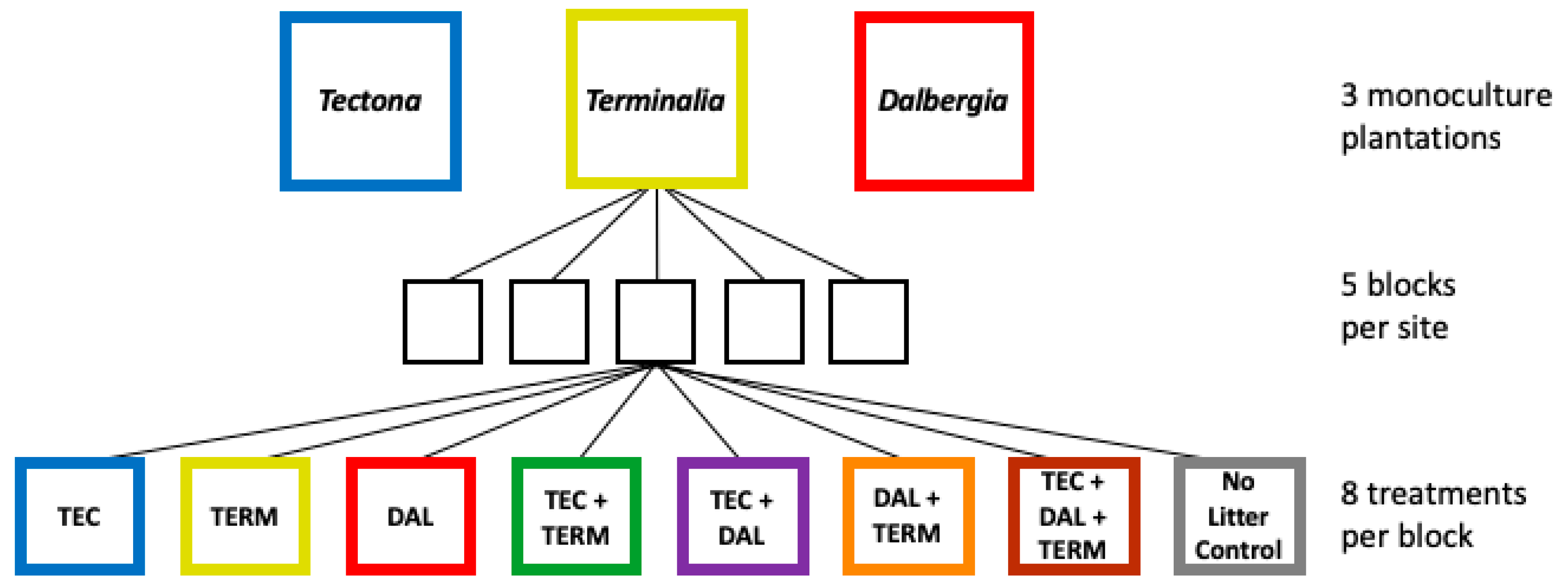
2.2. Experimental Design
2.3. Field Measurements and Sampling
2.4. Laboratory Analyses
2.4.1. Soil and Litter Nutrients
2.4.2. Soil Microbial Biomass by Fumigation-Extraction
2.4.3. Litter Fiber and Lignin
2.5. Data Analyses
Homefield Advantage and Non-Additive Effects of Species Mixtures
2.6. Statistical Analyses
3. Results
3.1. Site and Litter Characteristics
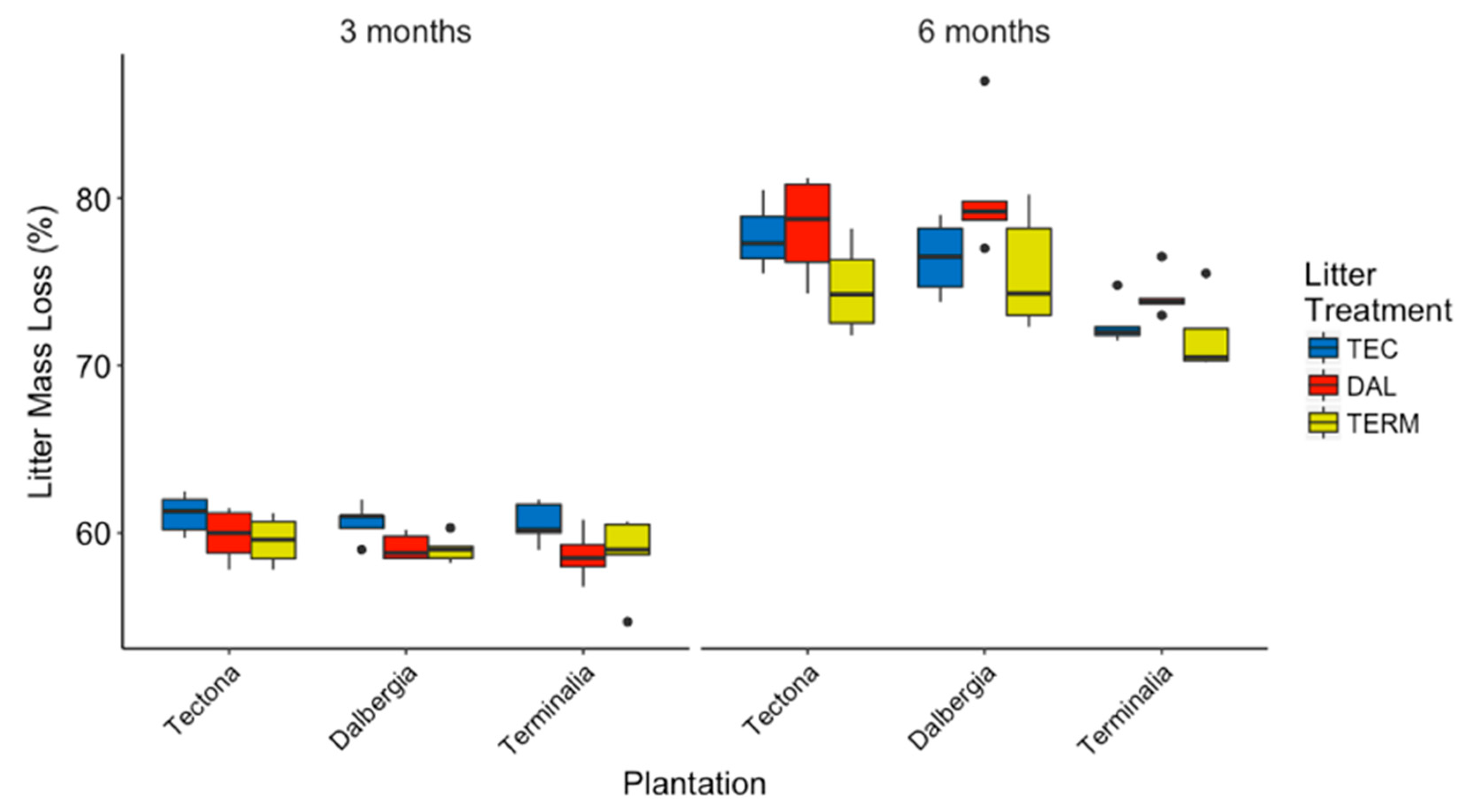
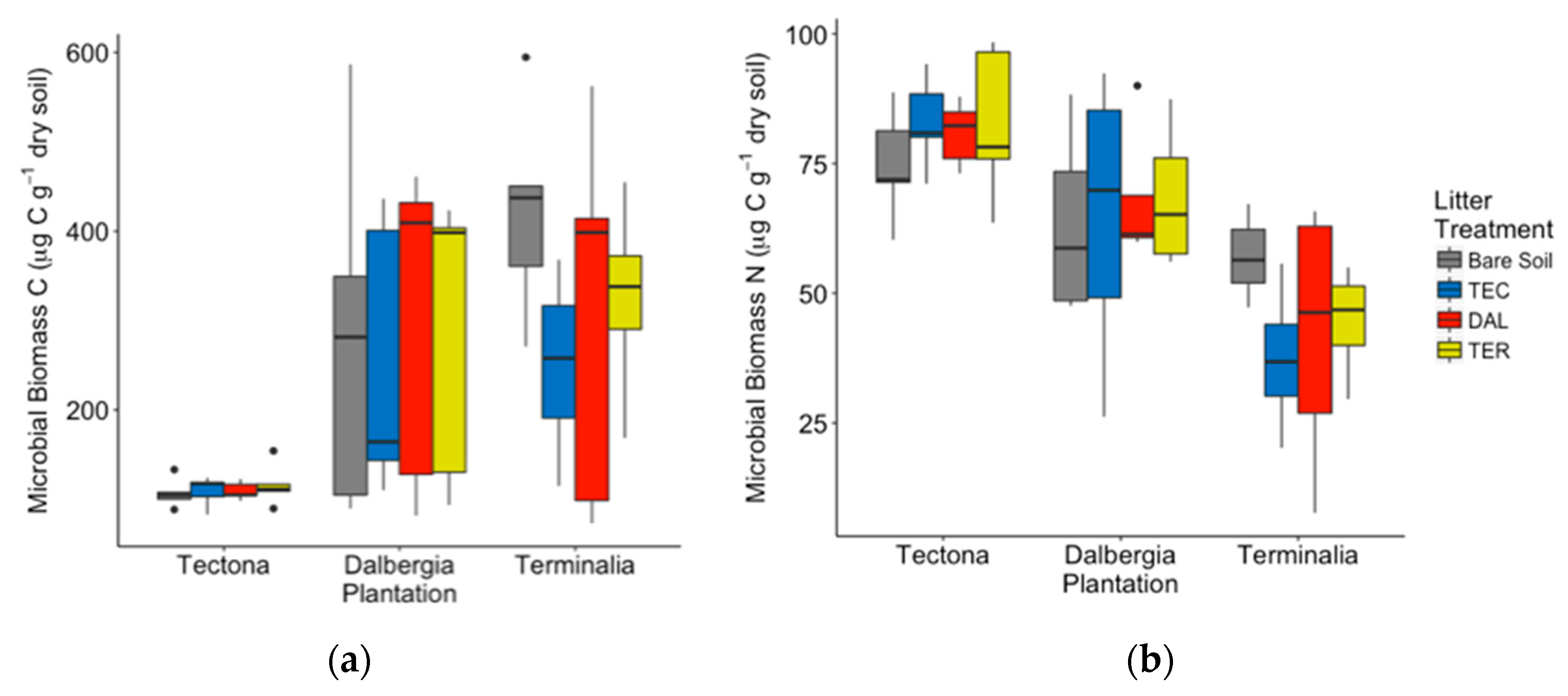
3.2. General Patterns for Individual Species
3.3. Homefield Advantage
3.4. Non-Additive Effects of Species Mixtures
4. Discussion
4.1. Differences in Decomposition and Soil Respiration among Plantation Types
4.2. Leaf Properties Explained some Variation in Litter Decomposition
4.3. Limited Evidence for Non-Additive Effects and Homefield Advantage during Decomposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stockmann, U.; Adams, M.A.; Crawford, J.W.; Field, D.J.; Henakaarchchi, N.; Jenkins, M.; Minasny, B.; McBratney, A.B.; De Courcelles, V.D.R.; Singh, K.; et al. The knowns, known unknowns and unknowns of sequestration of soil organic carbon. Agric. Ecosyst. Environ. 2013, 164, 80–99. [Google Scholar] [CrossRef]
- Dixon, R.K.; Brown, S.; Houghton, R.A.; Solomon, A.M.; Trexier, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Mayaux, P.; Holmgren, P.; Achard, F.; Eva, H.; Stibig, H.; Branthomme, A. Tropical forest cover change in the 1990s and options for future monitoring. Philos. Trans. R. Soc. B-Biol. Sci. 2005, 360, 373–384. [Google Scholar] [CrossRef] [PubMed]
- IUCN The Bonn Challenge. Available online: http://www.bonnchallenge.org/content/challenge (accessed on 7 January 2019).
- Food and Agricultural Organization. Global Forest Resources Assessment 2000: Main Report Food and Agricultural Organisation of the United Nations, National Forestry Action Plans and Forest Resources Assessment, FAO. Available online: http://www.fao.org/forestry/site/fra2000report (accessed on 7 January 2019).
- Stefanski, S.F.; Shi, X.; Hall, J.S.; Hernandez, A.; Fenichel, E.P. Teak-cattle production tradeoffs for Panama Canal Watershed small scale producers. For. Policy Econ. 2015, 56, 48–56. [Google Scholar] [CrossRef]
- Potvin, C.; Mancilla, L.; Buchmann, N.; Monteza, J.; Moore, T.; Murphy, M.; Oelmann, Y.; Scherer-Lorenzen, M.; Turner, B.L.; Wilcke, W.; et al. An ecosystem approach to biodiversity effects: Carbon pools in a tropical tree plantation. For. Ecol. Manag. 2011, 261, 1614–1624. [Google Scholar] [CrossRef]
- Hall, J.S.; Ashton, M.S.; Garen, E.J.; Jose, S. The ecology and ecosystem services of native trees: Implications for reforestation and land restoration in Mesoamerica. For. Ecol. Manag. 2011, 261, 1553–1557. [Google Scholar] [CrossRef]
- Schwendenmann, L.; Pendall, E.; Potvin, C. Surface soil organic carbon pools, mineralization and CO2 efflux rates under different land-use types in Central Panama. In Stability of Tropical Rainforest Margins. Environmental Science and Engineering (Environmental Science); Tscharntke, T., Leuschner, C., Zeller, M., Guhardja, E., Bidin, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Kraenzel, M.; Castillo, A.; Moore, T.; Potvin, C. Carbon storage of harvest-age teak (Tectona grandis) plantations, Panama. For. Ecol. Manag. 2003, 173, 213–225. [Google Scholar] [CrossRef]
- Ruiz-Jaen, M.C.; Potvin, C. Can we predict carbon stocks in tropical ecosystems from tree diversity? Comparing species and functional diversity in a plantation and a natural forest. New Phytol. 2011, 189, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.; Tiunov, A.; Scheu, S. Biodiversity and litter decomposition in terrestrial ecosystems. Annu. Rev. Ecol. Evol. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Swift, R.S. Sequestration of carbon by soil. Soil Sci. 2001, 166, 858–871. [Google Scholar] [CrossRef]
- Berg, B.; Berg, M.P.; Bottner, P.; Box, E.; Breymeyer, A.; Ca de Anta, R.; Couteaux, M.; Escudero, A.; Gallardo, A.; Kratzet, W.; et al. Litter mass loss rates in pine forest of Europe and Eastern United States: Some relationships with climate and litter quality. Biogeochemistry 1993, 20, 127–159. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Cornelissen, J.H.C.; Vendramini, F.; Cabido, M.; Castellanos, A. Chemistry and toughness predict leaf litter decomposition rates over a wide spectrum of functional types and taxa in central Argentina. Plant Soil 2000, 218, 21–30. [Google Scholar] [CrossRef]
- Talbot, J.M.; Treseder, K.K. Interactions among lignin, cellulose, and nitrogen drive litter chemistry-decay relationships. Ecology 2012, 93, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Laird-Hopkins, B.C.; Bréchet, L.M.; Trujillo, B.C.; Sayer, E.J. Tree functional diversity affects litter decomposition and arthropod community composition in a tropical forest. Biotropica 2017, 49, 903–911. [Google Scholar] [CrossRef]
- Fornara, D.A.; Tilman, D.; Hobbie, S.E. Linkages between plant functional composition, fine root processes and potential soil N mineralization rates. J. Ecol. 2009, 97, 48–56. [Google Scholar] [CrossRef]
- Hobbie, S.E. Effects of plant species on nutrient cycling. Trends Ecol. Evol. 1992, 7, 336–339. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Jørgensen, H.B. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. J. Ecol. 2010, 98, 754–763. [Google Scholar] [CrossRef]
- Sayer, E.J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biol. Rev. Camb. Philos. Soc. 2006, 81, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Liu, L.L.; Sayer, E.J. Variability of above-ground litter inputs alters soil physicochemical and biological processes: A meta-analysis of litterfall-manipulation experiments. Biogeosciences 2013, 10, 7423–7433. [Google Scholar] [CrossRef]
- Ayres, E.; Steltzer, H.; Berg, S.; Wall, D.H. Soil biota accelerate decomposition in high-elevation forests by specializing in the breakdown of litter produced by the plant species above them. J. Ecol. 2009, 97, 901–912. [Google Scholar] [CrossRef]
- Freschet, G.T.; Aerts, R.; Cornelissen, J.H.C. Multiple mechanisms for trait effects on litter decomposition: Moving beyond home-field advantage with a new hypothesis. J. Ecol. 2012, 100, 619–630. [Google Scholar] [CrossRef]
- Cummings, J.A.; Parker, I.M.; Gilbert, G.S. Allelopathy: A tool for weed management in forest restoration. Plant Ecol. 2013, 213, 1975–1989. [Google Scholar] [CrossRef]
- Fanin, N.; Hättenschwiler, S.; Barantal, S.; Schimann, H.; Fromin, N. Does variability in litter quality determine soil microbial respiration in an Amazonian rainforest? Soil Biol. Biochem. 2011, 43, 1014–1022. [Google Scholar] [CrossRef]
- Liang, J.; Lu, Z.; Yu, Z.; Wang, J.; Wang, X. Effects of leaf litter extraction fluid from dominant forest tree species on functional characteristics of soil microbial communities. J. For. Res. 2016, 27, 81–90. [Google Scholar] [CrossRef]
- Mayoral, C.; van Breugel, M.; Cerezo, A.; Hall, J.S. Survival and growth of five Neotropical timber species in monocultures and mixtures. For. Ecol. Manag. 2017, 403, 1–11. [Google Scholar] [CrossRef]
- Turner, B.L.; Engelbrecht, B.M.J. Soil organic phosphorus in lowland tropical rain forests. Biogeochemistry 2011, 103, 297–315. [Google Scholar] [CrossRef]
- Ogden, F.L.; Crouch, T.D.; Stallard, R.F.; Hall, J.S. Effect of land cover and use on dry season river runoff, runoff efficiency, and peak storm runoff in the seasonal tropics of Central Panama. Water Resour. Res. 2013, 49, 8443–8462. [Google Scholar] [CrossRef]
- Batterman, S.A.; Hall, J.S.; Turner, B.L.; Hedin, L.O.; LaHaela Walter, J.K.; Sheldon, P.; van Breugel, M. Phosphatase activity and nitrogen fixation reflect species differences, not nutrient trading or nutrient balance, across tropical rainforest trees. Ecol. Lett. 2018, 21, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Sayer, E.J.; Tanner, E.V.J. A new approach to trenching experiments for measuring root–rhizosphere respiration in a lowland tropical forest. Soil Biol. Biochem. 2010, 42, 347–352. [Google Scholar] [CrossRef]
- Jones, D.L.; Willett, B.M.J. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. JDS 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 12 December 2018).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.3-2, 2015. Available online: https://CRAN.R-project.org/package=vegan. Retrieved from https://github.com/vegandevs/vegan (accessed on 12 December 2018).
- Bates, D.M.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed- Effects Models using lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley & Sons: Chichester, UK, 2007. [Google Scholar]
- Sayer, E.J.; Tanner, E.V.J.; Lacey, A.L. Effects of litter manipulation on early-stage decomposition and meso-arthropod abundance in a tropical moist forest. For. Ecol. Manag. 2006, 229, 285–293. [Google Scholar] [CrossRef]
- Hawkes, C.V.; Belnap, J.; D’Antonio, C.; Firestone, M.K. Arbuscular mycorrhizal assemblages in native plant roots change in the presence of invasive exotic grasses. Plant Soil 2006, 281, 369–380. [Google Scholar] [CrossRef]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-Induced Resistance and Priming of Plant Defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Leela, P.; Arumugam, K. Allelopathic influence of teak (Tectona Grandis L.) leaves on growth responses of green gram (Vigna Radiata (L.) Wilczek) and Chilli (Capisum Frutescens L.). Int. J. Curr. Biotechnol. 2014, 2, 55–58. [Google Scholar]
- Ajeesh, R.; Santhoshkumar, A.V.; Gopal, S.; Binu, N.K. Screening of selected native arbuscular mycorrhizal fungi at different levels for their symbiotic efficiency with tectona grandis seedlings. J. Trop. For. Sci. 2017, 29, 395–403. [Google Scholar]
- Högberg, M.N.; Högberg, P. Extramatrical ectomycorrhizal mycelium contributes one-third of microbial biomass and produces, together with associated roots, half the dissolved organic carbon in a forest soil. New Phytol. 2002, 154, 791–795. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Takahashi, E.N.; Ryan, M.G. Tree-girdling to separate root and heterotrophic respiration in two Eucalyptus stands in Brazil. Oecologia 2006, 148, 447–454. [Google Scholar] [CrossRef] [PubMed]
- King, J.; Pregitzer, K.; Zak, D.; Sober, J.; Isebrands, J.; Dickson, R.; Hendrey, G.; Karnosky, D. Fine-root biomass and fluxes of soil carbon in young stands of paper birch and trembling aspen as affected by elevated atmospheric CO2 and tropospheric O3. Oecologia 2001, 128, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Trumbore, E.; Davidson, A.; Camargo, D.; Nepstad, C. Belowground cycling of carbon in forests and pastures of Eastern Amazonia. Global Biogeochem. Cycles 1995, 9, 515–528. [Google Scholar] [CrossRef]
- Yi, Z.; Fu, S.; Yi, W.; Zhou, G.; Mo, J.; Zhang, D.; Ding, M.; Wang, X.; Zhou, L. Partitioning soil respiration of subtropical forests with different successional stages in south China. For. Ecol. Manag. 2007, 243, 178–186. [Google Scholar] [CrossRef]
- Ferreira, V.; Gulis, V.; Graça, M.A.S. Whole-stream nitrate addition affects litter decomposition and associated fungi but not invertebrates. Oecologia 2006, 149, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Bréchet, L.; Le Dantec, V.; Ponton, S.; Goret, J.Y.; Sayer, E.; Bonal, D.; Freycon, V.; Roy, J.; Epron, D. Short- and Long-term Influence of Litter Quality and Quantity on Simulated Heterotrophic Soil Respiration in a Lowland Tropical Forest. Ecosystems 2017, 20, 1190–1204. [Google Scholar] [CrossRef]
- Hjältén, J. Simulating Herbivory: Problems and Possibilities. In Insects and Ecosystem Function; Weisser, W.W., Siemann, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 243–255. [Google Scholar] [CrossRef]
- Paul, G.S.; Montagnini, F.; Berlyn, G.P.; Craven, D.J.; van Breugel, M.; Hall, J.S. Foliar herbivory and leaf traits of five native tree species in a young plantation of Central Panama. New For. 2012, 43, 69–87. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Coq, S.; Barantal, S.; Handa, I.T. Leaf traits and decomposition in tropical rainforest: Revisiting some commonly held views towards a new hypothesis. New Phytol. 2010, 189, 950–965. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bonner, K.I.; Barker, G.M. Linkages between plant litter decomposition, litter quality, and vegetation responses to herbivores. Funct. Ecol. 2002, 16, 585–595. [Google Scholar] [CrossRef]
- Hobbie, S.; Vitousek, P.M. Nutrient limitation of decomposition in Hawaiian forests. Ecology 2000, 81, 1867–1877. [Google Scholar] [CrossRef]
- Gartner, T.B.; Cardon, Z.G. Decomposition Dynamics in Mixed-Species Leaf Litter. Oikos 2012, 104, 230–246. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Gasser, P. Soil animals alter plant litter diversity effects on decomposition. Proc. Natl. Acad. Sci. USA 2005, 102, 1519–1524. [Google Scholar] [CrossRef] [PubMed]
- Joly, F.X.; Fromin, N.; Kiikkilä, O.; Hättenschwiler, S. Diversity of leaf litter leachates from temperate forest trees and its consequences for soil microbial activity. Biogeochemistry 2016, 129, 373–388. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property/Plantation | Tectona | Dalbergia | Terminalia |
|---|---|---|---|
| Total C (%) | 4.58 ± 0.20 | 4.38 ± 0.08 | 4.90 ± 0.23 |
| Total N (%) | 0.41 ± 0.02 A | 0.45 ± 0.01 B | 0.44 ± 0.02 B |
| C:N ratio | 11.29 ± 0.13 a | 9.69 ±0.07 b | 11.07 ± 0.32 a |
| P (mg kg−1) | 3.35 ± 0.30 | 3.09 ± 0.17 | 3.75 ± 0.34 |
| K (mg kg−1) | 759 ± 72 a | 501 ± 58 b | 651 ± 55 a |
| pH | 5.064 ± 0.08 | 4.93 ± 0.08 | 5.032 ± 0.06 |
| Litter Nutrients\Litter Type | TEC | DAL | TERM |
|---|---|---|---|
| Total C % | 49.31 | 46.91 | 47.28 |
| Total N % | 1.48 | 2.24 | 1.03 |
| C:N ratio | 33.2 | 20.9 | 46 |
| P % | 0.0894 | 0.0399 | 0.0568 |
| K % | 0.468 | 0.479 | 0.673 |
| Fibre % | 29.9 | 22.8 | 24.5 |
| Lignin (L) % | 13.6 | 8.1 | 10.2 |
| L:N | 10.18 | 3.86 | 10.29 |
| Ca % | 1.23 | 1.35 | 2.16 |
| Mg % | 0.36 | 0.322 | 0.215 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerdraon, D.; Drewer, J.; Castro, B.; Wallwork, A.; Hall, J.S.; Sayer, E.J. Litter Traits of Native and Non-Native Tropical Trees Influence Soil Carbon Dynamics in Timber Plantations in Panama. Forests 2019, 10, 209. https://doi.org/10.3390/f10030209
Kerdraon D, Drewer J, Castro B, Wallwork A, Hall JS, Sayer EJ. Litter Traits of Native and Non-Native Tropical Trees Influence Soil Carbon Dynamics in Timber Plantations in Panama. Forests. 2019; 10(3):209. https://doi.org/10.3390/f10030209
Chicago/Turabian StyleKerdraon, Deirdre, Julia Drewer, Biancolini Castro, Abby Wallwork, Jefferson S. Hall, and Emma J. Sayer. 2019. "Litter Traits of Native and Non-Native Tropical Trees Influence Soil Carbon Dynamics in Timber Plantations in Panama" Forests 10, no. 3: 209. https://doi.org/10.3390/f10030209
APA StyleKerdraon, D., Drewer, J., Castro, B., Wallwork, A., Hall, J. S., & Sayer, E. J. (2019). Litter Traits of Native and Non-Native Tropical Trees Influence Soil Carbon Dynamics in Timber Plantations in Panama. Forests, 10(3), 209. https://doi.org/10.3390/f10030209