Understory Plant Functional Types Alter Stoichiometry Correlations between Litter and Soil in Chinese Fir Plantations with N and P Addition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Experimental Design
2.3. Sample Collection
2.4. Litter and Soil Nutrient Measurement
2.5. Data Analysis
3. Results
3.1. Availability of Soil Nutrients and Carbon, Nitrogen, and Phosphorus (CNP) Concentration of Plant Litter and Soil
3.2. Effects of N and P Addition on Carbon, Nitrogen, and Phosphorus (C, N, and P) Concentrations and Stoichiometric Ratios of Different Functional Groups
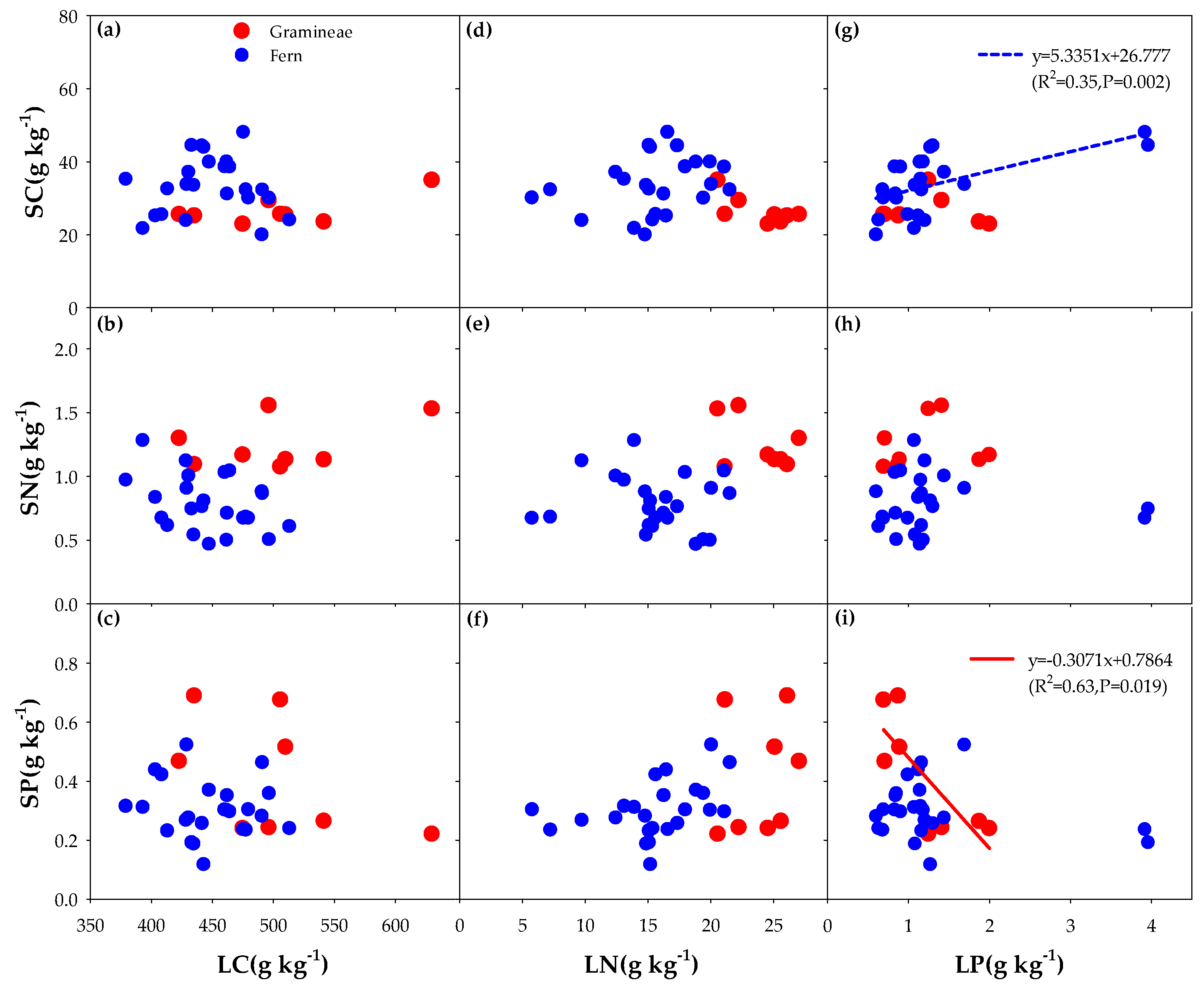
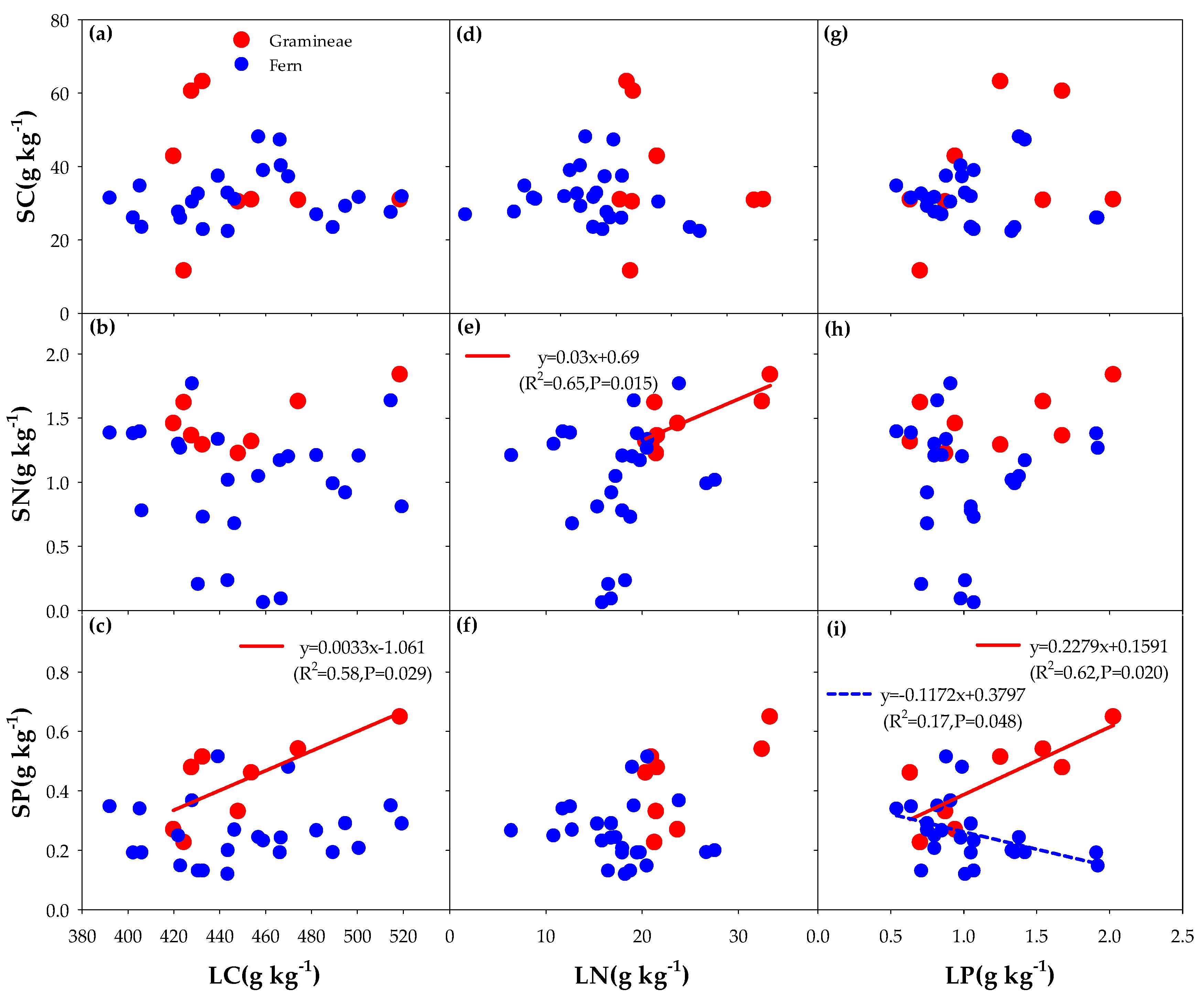
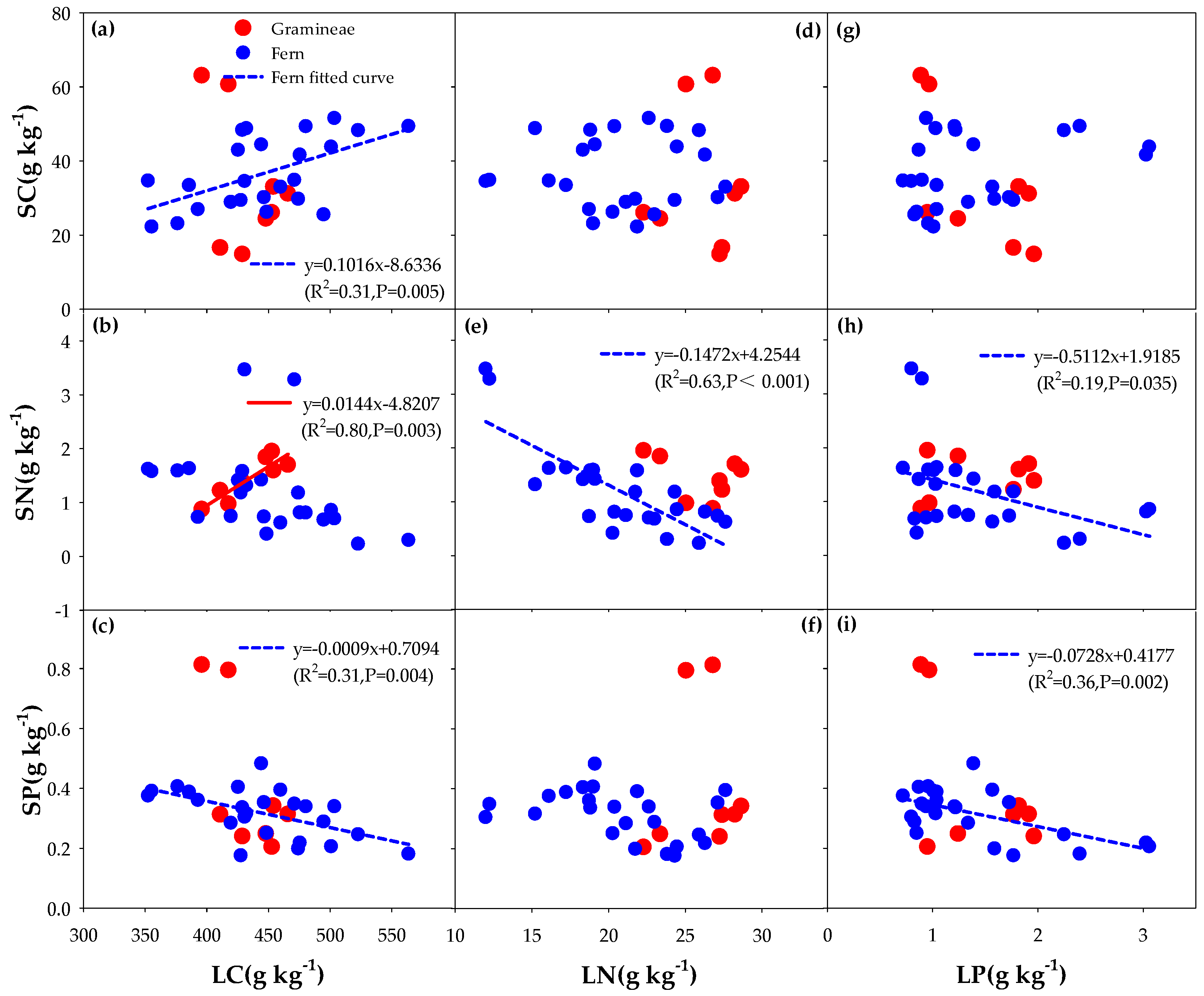
3.3. Correlation between the Litter of Different Functional Groups and Soil Carbon, Nitrogen, and Phosphorus (C, N, and P) with the Addition of N and P
3.4. Effects of N and P Addition on Carbon, Nitrogen, and Phosphorus (C, N, and P) Concentrations and Stoichiometric Ratios of the Soils Associated with Different Plant Functional Groups
4. Discussion
4.1. Effects of Different Understory Vegetation Types on Stoichiometric Ratio
4.2. Effect of Nitrogen (N) and Phosphorus (P) Addition on Stoichiometric Ratio
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| C | carbon |
| N | nitrogen |
| P | phosphorus |
| DOC | dissolved organic carbon |
| TOC | total organic carbon |
| TN | total nitrogen |
| TP | total phosphorus |
| AN | available nitrogen |
| AP | available phosphorus |
| LC | total litter organic carbon |
| LN | total litter nitrogen |
| LP | total litter phosphorus |
| SC | total soil organic carbon |
| SN | total soil nitrogen |
| SP | total soil phosphorus |
| Sp | understory species |
| F | plant functional group |
| B&R | block as random effect |
| L C/N | litter carbon to nitrogen ratio |
| L C/P | litter carbon to phosphorus ratio |
| L N/P | litter nitrogen to phosphorus ratio |
| S C/N | soil carbon to nitrogen ratio |
| S C/P | soil carbon to phosphorus ratio |
| S N/P | soil nitrogen to phosphorus ratio |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No P Addition | P Addition | ||||
|---|---|---|---|---|---|---|
| CK | N | CK | N | |||
| Lophatherum gracile (Gramineae) | Litter | C | 502.10 ± 22.99 | 449.88 ± 11.72 | 474.09 ± 13.99 | 434.28 ± 8.70 |
| N | 24.04 ± 0.85 | 24.38 ± 1.89 | 23.00 ± 1.63 | 26.14 ± 0.82 | ||
| P | 1.21 ± 0.18 | 1.21 ± 0.18 | 1.69 ± 0.40 | 1.44 ± 0.17 | ||
| soil | C | 26.61 ± 1.37 | 37.68 ± 6.08 | 35.88 ± 8.32 | 33.71 ± 6.54 | |
| N | 1.25 ± 0.07 | 1.47 ± 0.07 | 1.12 ± 0.03 | 1.44 ± 0.14 | ||
| P | 0.41 ± 0.07 | 0.43 ± 0.05 | 0.37 ± 0.03 | 0.41 ± 0.09 | ||
| Woodwardia japonica (Fern) | Litter | C | 420.27 ± 12.05 | 421.31 ± 9.00 | 442.22 ± 19.48 | 393.95 ± 11.78 |
| N | 15.36 ± 1.39 | 17.56 ± 1.23 | 16.43 ± 0.93 | 18.95 ± 0.67 | ||
| P | 1.23 ± 0.08 | 1.13 ± 0.18 | 1.33 ± 0.13 | 1.05 ± 0.08 | ||
| soil | C | 29.38 ± 2.08 | 29.88 ± 2.15 | 25.22 ± 1.75 | 32.07 ± 2.97 | |
| N | 0.96 ± 0.07 | 1.18 ± 0.10 | 1.00 ± 0.13 | 1.34 ± 0.14 | ||
| P | 0.38 ± 0.03 | 0.29 ± 0.05 | 0.40 ± 0.03 | 0.39 ± 0.02 | ||
| Dryopteris atrata (Fern) | Litter | C | 443.67 ± 6.65 | 457.00 ± 6.40 | 454.05 ± 15.97 | 483.93 ± 15.52 |
| N | 16.62 ± 0.68 | 19.82 ± 1.65 | 18.77 ± 1.14 | 25.17 ± 0.69 | ||
| P | 1.88 ± 0.45 | 1.16 ± 0.09 | 2.57 ± 0.42 | 2.18 ± 0.22 | ||
| soil | C | 40.89 ± 1.94 | 35.70 ± 3.45 | 31.08 ± 1.68 | 38.16 ± 3.02 | |
| N | 0.64 ± 0.05 | 0.60 ± 0.17 | 0.40 ± 0.03 | 0.74 ± 0.13 | ||
| P | 0.24 ± 0.03 | 0.19 ± 0.04 | 0.40 ± 0.08 | 0.25 ± 0.03 | ||
| Dicranopteris dichotoma (Fern) | Litter | C | 480.50 ± 6.65 | 476.03 ± 13.68 | 485.45 ± 19.59 | 461.29 ± 10.67 |
| N | 14.74 ± 1.94 | 15.37 ± 1.90 | 12.01 ± 1.54 | 18.08 ± 1.55 | ||
| P | 0.75 ± 0.04 | 0.84 ± 0.04 | 1.53 ± 0.17 | 0.97 ± 0.06 | ||
| soil | C | 30.64 ± 2.27 | 29.51 ± 0.70 | 34.01 ± 4.97 | 39.89 ± 3.83 | |
| N | 0.77 ± 0.07 | 1.19 ± 0.144 | 1.18 ± 0.15 | 1.53 ± 0.42 | ||
| P | 0.30 ± 0.02 | 0.29 ± 0.02 | 0.38 ± 0.03 | 0.32 ± 0.01 | ||

References
- Elser, J.; Sterner, R.; Fagan, W.; Markow, T.; Cotner, J.; Hobbie, S.; Odell, G.; Weider, L.; Gorokhova, E.; Harrison, J. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladanai, S.; Ågren, G.I.; Olsson, B.A. Relationships between tree and soil properties in picea abies and pinus sylvestris forests in sweden. Ecosystems 2010, 13, 302–316. [Google Scholar] [CrossRef]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C: N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Siddique, I.; Vieira, I.C.G.; Schmidt, S.; Lamb, D.; Carvalho, C.J.R.; Figueiredo, R.D.O.; Blomberg, S.; Davidson, E.A. Nitrogen and phosphorus additions negatively affect tree species diversity in tropical forest regrowth trajectories. Ecology 2010, 91, 2121–2131. [Google Scholar] [CrossRef]
- Wei, X.; Blanco, J.A.; Jiang, H.; Kimmins, J.H. Effects of nitrogen deposition on carbon sequestration in Chinese fir forest ecosystems. Sci. Total Environ. 2012, 416, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Litton, C.M.; Raich, J.W.; Ryan, M.G. Carbon allocation in forest ecosystems. Glob. Chang. Biol. 2007, 13, 2089–2109. [Google Scholar] [CrossRef] [Green Version]
- Clark, C.M.; Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 2008, 451, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Sardans, J.; Rivas-Ubach, A.; Janssens, I.A. The human-induced imbalance between C, N and P in Earth’s life system. Glob. Chang. Biol. 2012, 18, 3–6. [Google Scholar] [CrossRef]
- Strengbom, J.; Walheim, M.; Näsholm, T.; Ericson, L. Regional differences in the occurrence of understorey species reflect nitrogen deposition in Swedish forests. Ambio 2003, 32, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Menge, D.N.L.; Field, C.B. Simulated global changes alter phosphorus demand in annual grassland. Glob. Chang. Biol. 2007, 13, 2582–2591. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives. Biogeochemistry 2012, 111, 1–39. [Google Scholar] [CrossRef]
- Mayor, J.R.; Wright, S.J.; Turner, B.L. Species-specific responses of foliar nutrients to long-term N and P additions in a lowland tropical forest. J. Ecol. 2014, 102, 36–44. [Google Scholar] [CrossRef]
- Huang, W.; Zhou, G.; Liu, J.; Zhang, D.; Xu, Z.; Liu, S. Effects of elevated carbon dioxide and nitrogen addition on foliar stoichiometry of nitrogen and phosphorus of five tree species in subtropical model forest ecosystems. Environ. Pollut. 2012, 168, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Koller, E.K.; Press, M.C.; Callaghan, T.V.; Phoenix, G.K. Tight coupling between shoot level foliar N and P, leaf area, and shoot growth in Arctic dwarf shrubs under simulated climate change. Ecosystems 2016, 19, 326–338. [Google Scholar] [CrossRef]
- Zhang, Q.; Xie, J.; Lyu, M.; Xiong, D.; Wang, J.; Chen, Y.; Li, Y.; Wang, M.; Yang, Y. Short-term effects of soil warming and nitrogen addition on the N:P stoichiometry of Cunninghamia lanceolata in subtropical regions. Plant Soil 2016, 411, 395–407. [Google Scholar] [CrossRef]
- Braun, S.; Thomas, V.F.; Quiring, R.; Flückiger, W. Does nitrogen deposition increase forest production? The role of phosphorus. Environ. Pollut. 2010, 158, 2043–2052. [Google Scholar] [CrossRef]
- Chen, F.S.; Niklas, K.J.; Zeng, D.H. Important foliar traits depend on species-grouping: Analysis of a remnant temperate forest at the Keerqin sandy lands, China. Plant Soil 2011, 340, 337–345. [Google Scholar] [CrossRef]
- Heuck, C.; Smolka, G.; Whalen, E.D.; Frey, S.; Gundersen, P.; Moldan, F.; Fernandez, I.J.; Spohn, M. Effects of long-term nitrogen addition on phosphorus cycling in organic soil horizons of temperate forests. Biogeochemistry 2018, 141, 167–181. [Google Scholar] [CrossRef]
- Zhang, J.; Yan, X.; Su, F.; Li, Z.; Wang, Y.; Wei, Y.; Ji, Y.; Yang, Y.; Zhou, X.; Guo, H.; et al. Long-term N and P additions alter the scaling of plant nitrogen to phosphorus in a Tibetan alpine meadow. Sci. Total Environ. 2018, 625, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Craft, C.B.; Vymazal, J.; Richardson, C.J. Response of everglades plant communities to nitrogen and phosphorus additions. Wetlands 1995, 15, 258–271. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Wang, W.Q.; Wang, C.; Sardans, J.; Zeng, C.S.; Tong, C.; Peñuelas, J. Plant invasive success associated with higher N-use efficiency and stoichiometric shifts in the soil–plant system in the Minjiang River tidal estuarine wetlands of China. Wetl. Ecol. Manag. 2015, 23, 865–880. [Google Scholar] [CrossRef]
- Wang, F.; Chen, F.; Wang, G.G.; Mao, R.; Fang, X.; Wang, H.; Bu, W. Effects of experimental nitrogen addition on nutrients and nonstructural carbohydrates of dominant understory plants in a Chinese fir plantation. Forests 2019, 10, 155. [Google Scholar] [CrossRef]
- Takafumi, H.; Hiura, T. Effects of disturbance history and environmental factors on the diversity and productivity of understory vegetation in a cool-temperate forest in Japan. For. Ecol. Manag. 2009, 257, 843–857. [Google Scholar] [CrossRef] [Green Version]
- Dobson, A.M.; Blossey, B.; Richardson, J.B. Invasive earthworms change nutrient availability and uptake by forest understory plants. Plant Soil 2017, 421, 175–190. [Google Scholar] [CrossRef]
- Wang, F.; Zou, B.; Li, H.; Li, Z. The effect of understory removal on microclimate and soil properties in two subtropical lumber plantations. J. For. Res. 2014, 19, 238–243. [Google Scholar] [CrossRef]
- Jules, M.J.; Sawyer, J.O.; Jules, E.S. Assessing the relationships between stand development and understory vegetation using a 420-year chronosequence. For. Ecol. Manag. 2008, 255, 2384–2393. [Google Scholar] [CrossRef]
- Lu, X.; Mo, J.; Gilliam, F.S.; Zhou, G.; Fang, Y. Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest. Glob. Chang. Biol. 2010, 16, 2688–2700. [Google Scholar] [CrossRef] [Green Version]
- Hurd, T.M.; Brach, A.R.; Raynal, D.J. Response of understory vegetation of Adirondack forests to nitrogen additions. Can. J. For. Res. 1998, 28, 799–807. [Google Scholar] [CrossRef]
- Rainey, S.M.; Nadelhoffer, K.J.; Silver, W.L.; Downs, M.R. Effects of chronic nitrogen additions on understory species in a red pine plantation. Ecol. Appl. 1999, 9, 949–957. [Google Scholar] [CrossRef]
- Bi, J.; Blanco, J.A.; Seely, B.; Kimmins, J.P.; Ding, Y.; Welham, C. Yield decline in Chinese-fir plantations: A simulation investigation with implications for model complexity. Can. J. For. Res. 2007, 37, 1615–1630. [Google Scholar] [CrossRef]
- Lu, Z.H.; Wu, G.; Ma, X.; Bai, G.X. Current situation of Chinese forestry tactics and strategy of sustainable development. J. For. Res. 2002, 13, 319–322. [Google Scholar]
- Chen, G.S.; Yang, Z.J.; Gao, R.; Xie, J.S.; Guo, J.F.; Huang, Z.Q.; Yang, Y.S. Carbon storage in a chronosequence of Chinese fir plantations in southern China. For. Ecol. Manag. 2013, 300, 68–76. [Google Scholar] [CrossRef]
- Tang, X.; Pérez-Cruzado, C.; Fehrmann, L.; Álvarez-González, J.G.; Lu, Y.; Kleinn, C. Development of a compatible taper function and stand-level merchantable volume model for Chinese fir plantations. PLoS ONE 2016, 11, e0147610. [Google Scholar] [CrossRef]
- Peng, Y.; Tian, D.; Thomas, S.C. Forest management and soil respiration: Implications for carbon sequestration. Environ. Rev. 2008, 16, 93–111. [Google Scholar] [CrossRef]
- Powers, R.F.; Busse, M.D.; McFarlane, K.J.; Zhang, J.W.; Young, D.H. Long-term effects of silviculture on soil carbon storage: Does vegetation control make a difference? Forestry 2013, 86, 47–58. [Google Scholar] [CrossRef]
- Chen, F.S.; Niklas, K.J.; Liu, Y.; Fang, X.M.; Wan, S.Z.; Wang, H. Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree Physiol. 2015, 35, 1106–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Tian, H. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. J. Geophys. Res. Space Phys. 2007, 112, 10–15. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, X.; Li, D.; Wang, H.; Chen, F.; Fu, X.; Fang, X.; Sun, X.; Yu, G. Impacts of nitrogen and phosphorus additions on the abundance and community structure of ammonia oxidizers and denitrifying bacteria in Chinese fir plantations. Soil Biol. Biochem. 2016, 103, 284–293. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; Agriculture Press: Beijing, China, 2008. (In Chinese) [Google Scholar]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Fisher, R.A.; Wardle, D.A. Plant communities as drivers of soil respiration: Pathways, mechanisms, and significance for global change. Biogeosciences 2011, 8, 2047–2061. [Google Scholar] [CrossRef]
- Meuleman, A.F.M.; Koerselman, W. The Vegetation N:P Ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar]
- Marklein, A.R.; Houlton, B.Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2011, 193, 696–704. [Google Scholar] [CrossRef]
- Lu, M.; Yang, Y.; Luo, Y.; Fang, C.; Zhou, X.; Chen, J.; Yang, X.; Li, B. Responses of ecosystem nitrogen cycle to nitrogen addition: A meta-analysis. New Phytol. 2011, 189, 1040–1050. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Chen, D.; Huang, G.; Zhou, L.; Fu, S. Understory plants can make substantial contributions to soil respiration: Evidence from two subtropical plantations. Soil Biol. Biochem. 2011, 43, 2355–2357. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Wang, X.; Sun, Y.X.; Zhou, L.; Lin, Y.; Fu, S. Effects of understory removal and tree girdling on soil microbial community composition and litter decomposition in two Eucalyptus plantations in South China. Funct. Ecol. 2011, 25, 921–931. [Google Scholar] [CrossRef]
- Yimer, F.; Ledin, S.; Abdelkadir, A. Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetation in the Bale Mountains, Ethiopia. Geoderma 2006, 135, 335–344. [Google Scholar] [CrossRef]
- Zhao, J.; Wan, S.; Li, Z.; Shao, Y.; Xu, G.; Liu, Z.; Zhou, L.; Fu, S. Dicranopteris-dominated understory as major driver of intensive forest ecosystem in humid subtropical and tropical region. Soil Biol. Biochem. 2012, 49, 78–87. [Google Scholar] [CrossRef]
- Pan, P.; Zhao, F.; Ning, J.; Zhang, L.; Ouyang, X.; Zang, H. Impact of understory vegetation on soil carbon and nitrogen dynamic in aerially seeded Pinus massoniana plantations. PLoS ONE 2018, 13, e0191952. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Stark, J.M. Plant species effects and carbon and nitrogen cycling in a sagebrush–crested wheatgrass soil. Soil Biol. Biochem. 2000, 32, 47–57. [Google Scholar] [CrossRef]
- Finzi, A.C.; Austin, A.T.; Cleland, E.E.; Frey, S.D.; Houlton, B.Z.; Wallenstein, M.D. Responses and feedbacks of coupled biogeochemical cycles to climate change: Examples from terrestrial ecosystems. Front. Ecol. Environ. 2011, 9, 61–67. [Google Scholar] [CrossRef]
- Penuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; Van Der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusià, J.; et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed]
- Klausmeier, C.A.; Litchman, E.; Daufresne, T.; Levin, S.A. Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton. Nature 2004, 429, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Moe, S.J.; Stelzer, R.S.; Forman, M.R.; Harpole, W.S.; Daufresne, T.; Yoshida, T. Recent advances in ecological stoichiometry: Insights for population and community ecology. Oikos 2005, 109, 29–39. [Google Scholar] [CrossRef]
- Van Heerwaarden, L.M.; Toet, S.; Aerts, R. Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization. J. Ecol. 2003, 91, 1060–1070. [Google Scholar] [CrossRef]
- Gusewell, S. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspect. Plant Ecol. Evol. Syst. 2002, 5, 37–61. [Google Scholar] [CrossRef]
- Bubier, J.L.; Smith, R.; Juutinen, S.; Moore, T.R.; Minocha, R.; Long, S.; Minocha, S. Effects of nutrient addition on leaf chemistry, morphology, and photosynthetic capacity of three bog shrubs. Oecologia 2011, 167, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J.; Cobb, E.D. N, P, and C stoichiometry of Eranthis hyemalis (Ranunculaceae) and the allometry of plant growth. Am. J. Bot. 2005, 92, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Bracken, M.E.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Factors | Lophatherum gracile Brongn. (Gramineae) | Woodwardia japonica (L.f.) Sm. (Fern) | Dryopteris atrata (Kunze) Ching (Fern) | Dicranopteris dichotoma (Thunb.) Berhn.) (Fern) |
|---|---|---|---|---|
| DOC (mg kg−1) | 88.05 ± 6.88 | 100.79 ± 8.23 | 103.53 ± 10.59 | 85.67 ± 4.64 |
| NH4+-N (mg kg−1) | 5.26 ± 0.40 | 5.01 ± 0.40 | 5.23 ± 0.32 | 5.45 ± 0.51 |
| NO3−-N (mg kg−1) | 1.92 ± 0.19 A | 1.41 ± 0.09 B | 1.40 ± 0.09 B | 1.42 ± 0.15 B |
| AN (mg kg−1) | 7.18 ± 0.50 | 6.42 ± 0.44 | 6.63 ± 0.37 | 6.87 ± 0.64 |
| AP (mg kg−1) | 1.17 ± 0.15 A | 1.15 ± 0.18 A | 1.34 ± 0.20 A | 0.56 ± 0.09 B |
| pH | 3.32 ± 0.05 | 3.31 ± 0.04 | 3.37 ± 0.04 | 3.33 ± 0.04 |
| LC (g kg−1) | 465.09 ± 8.61 A | 419.44 ± 7.18 B | 459.67 ± 6.32 A | 475.82 ± 6.62 A |
| LN (g kg−1) | 24.39 ± 0.69 A | 17.07 ± 0.57 C | 20.10 ± 0.77 B | 15.05 ± 0.92 C |
| LP (g kg−1) | 1.39 ± 0.12 B | 1.18 ± 0.06 BC | 1.94 ± 0.18 A | 1.02 ± 0.07 C |
| SC (g kg−1) | 31.57 ± 2.10 BC | 27.87 ± 0.93 C | 37.78 ± 1.39 A | 35.88 ± 2.00 AB |
| SN (g kg−1) | 1.30 ± 0.04 A | 1.10 ± 0.05 B | 0.62 ± 0.05 C | 1.22 ± 0.10 AB |
| SP (g kg−1) | 0.43 ± 0.02 A | 0.39 ± 0.02 A | 0.29 ± 0.02 B | 0.32 ± 0.01 B |
| Factor | df | LC (g kg−1) | LN (g kg−1) | LP (g kg−1) | L C/N | L C/P | L N/P | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS | F | MS | F | MS | F | MS | F | MS | F | MS | F | ||
| N | 2 | 8256 | 5.92 *** | 113 | 10.10 *** | 0.70 | 2.30 | 376 | 5.19 ** | 323 | 0.02 | 90.33 | 2.47 |
| P | 1 | 26.78 | 0.02 | 17.18 | 1.54 | 10.37 | 34.22 *** | 56.65 | 0.78 | 618,936 | 37.03 *** | 982 | 26.84 *** |
| N*P | 2 | 2440 | 26.71 | 28.54 | 2.56 | 0.81 | 2.66 | 70.56 | 0.98 | 27,073 | 1.62 | 156 | 4.26 * |
| f | 1 | 23,734 | 17.00 *** | 2066 | 185.37 *** | 0.01 | 0.04 | 2777 | 38.41 *** | 2941 | 0.18 | 1472 | 40.26 *** |
| N*f | 2 | 10,323 | 7.40 *** | 21.97 | 1.97 | 0.12 | 0.40 | 49.92 | 0.69 | 28,014 | 1.68 | 58.89 | 1.61 |
| P*f | 1 | 497.30 | 0.36 | 31.06 | 2.79 | 0.11 | 0.38 | 64.55 | 0.89 | 2928 | 0.18 | 71.35 | 1.95 |
| N*P*f | 2 | 3603 | 2.58 | 2.88 | 0.26 | 0.02 | 0.05 | 36.05 | 0.50 | 884 | 0.05 | 23.27 | 0.64 |
| Sp[f] | 2 | 47,217 | 33.83 *** | 398 | 35.69 *** | 13.89 | 45.85 *** | 2517 | 34.81 *** | 660,744 | 39.53 *** | 174 | 4.76 *** |
| Sp*N[f] | 4 | 2545 | 1.82 | 11.58 | 1.04 | 0.26 | 0.86 | 68.92 | 0.95 | 4379 | 0.26 | 1.59 | 0.04 |
| Sp*P[f] | 2 | 2714 | 1.94 | 19.89 | 1.79 | 3.35 | 11.04 *** | 7.05 | 0.10 | 38,456 | 2.30 | 80.97 | 2.21 |
| B&R | 3 | 5465 | 3.92 *** | 201 | 18.01 *** | 1.16 | 3.82 ** | 447 | 6.19 *** | 59,442 | 3.56* | 43.52 | 1.19 |
| Factor | df | SC (g kg−1) | SN (g kg−1) | SP (g kg−1) | S C/N | S C/P | S N/P | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS | F | MS | F | MS | F | MS | F | MS | F | MS | F | ||
| N | 2 | 182 | 1.41 | 0.90 | 5.59 ** | 0.05 | 3.49 * | 914 | 0.24 | 5964 | 2.92 | 18.93 | 7.39 *** |
| P | 1 | 109 | 0.84 | 0.12 | 0.73 | 0.05 | 3.29 | 1178 | 0.31 | 1865 | 0.91 | 3.38 | 1.32 |
| N*P | 2 | 16.63 | 0.13 | 0.33 | 2.03 | 0.01 | 0.64 | 1479 | 0.39 | 13.27 | 0.01 | 5.32 | 2.08 |
| f | 1 | 186 | 1.44 | 3.75 | 23.39 *** | 0.30 | 21.79 *** | 31,687 | 8.31 ** | 41,992 | 20.54 *** | 5.85 | 2.28 |
| N*f | 2 | 278 | 2.15 | 0.05 | 0.31 | 0.02 | 1.78 | 821 | 0.22 | 4381 | 2.14 | 2.97 | 1.16 |
| P*f | 1 | 38.80 | 0.30 | 0.62 | 3.86 * | 0.05 | 3.58 | 7680 | 2.01 | 11,111 | 5.43 * | 0.55 | 0.22 |
| N*P*f | 2 | 356 | 2.76 | 0.02 | 0.12 | 0.01 | 0.92 | 1074 | 0.28 | 2776 | 1.36 | 2.39 | 0.93 |
| Sp[f] | 2 | 1327 | 10.28 *** | 4.77 | 29.77 *** | 0.13 | 9.57 *** | 90,281 | 23.68 *** | 68,901 | 33.70 *** | 20.70 | 8.08 *** |
| Sp*N[f] | 4 | 176 | 1.36 | 0.12 | 0.78 | 0.01 | 1.07 | 5364 | 1.41 | 3802 | 1.86 | 2.05 | 0.80 |
| Sp*P[f] | 2 | 22.99 | 0.18 | 0.12 | 0.76 | 0.04 | 2.91 | 15,940 | 4.18 * | 8820 | 4.31 * | 5.02 | 1.96 |
| B&R | 3 | 325 | 2.52 | 1.41 | 8.83 *** | 0.17 | 12.44 *** | 26,816 | 7.04 *** | 6256 | 3.06 * | 33.72 | 13.16 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, J.; Fang, H.; Zhang, Q.; Chen, M.; Xu, X.; Pan, J.; Gao, Y.; Fang, X.; Guo, X.; Zhang, L. Understory Plant Functional Types Alter Stoichiometry Correlations between Litter and Soil in Chinese Fir Plantations with N and P Addition. Forests 2019, 10, 742. https://doi.org/10.3390/f10090742
Xie J, Fang H, Zhang Q, Chen M, Xu X, Pan J, Gao Y, Fang X, Guo X, Zhang L. Understory Plant Functional Types Alter Stoichiometry Correlations between Litter and Soil in Chinese Fir Plantations with N and P Addition. Forests. 2019; 10(9):742. https://doi.org/10.3390/f10090742
Chicago/Turabian StyleXie, Junyi, Haifu Fang, Qiang Zhang, Mengyun Chen, Xintong Xu, Jun Pan, Yu Gao, Xiangmin Fang, Xiaomin Guo, and Ling Zhang. 2019. "Understory Plant Functional Types Alter Stoichiometry Correlations between Litter and Soil in Chinese Fir Plantations with N and P Addition" Forests 10, no. 9: 742. https://doi.org/10.3390/f10090742