Pedunculate Oak Leaf Miners’ Community: Urban vs. Rural Habitat


Abstract
:
1. Introduction
2. Materials and Methods
2.1. The Host Plant and the Investigated Insects
2.2. Study Sites
2.3. Sampling Design
2.4. Characteristics of the Leaf Miner Community Analyzed
- 1.
- Species richness (number of species identified);
- 2.
- The abundance of individual species (Ai) (number of mines per 100 leaves), calculated as:
- 3.
- The abundance of leaf miners (Ab) calculated as a sum of all individual species’ abundance;
- 4.
- Shannon index of diversity (H), calculated as:
- 5.
- Frequency of occurrence (Fq), calculated as:
2.5. Statistical Analysis
3. Results
3.1. Overall Species Richness, Frequency of Occurrence, and Abundance of Leaf Miners
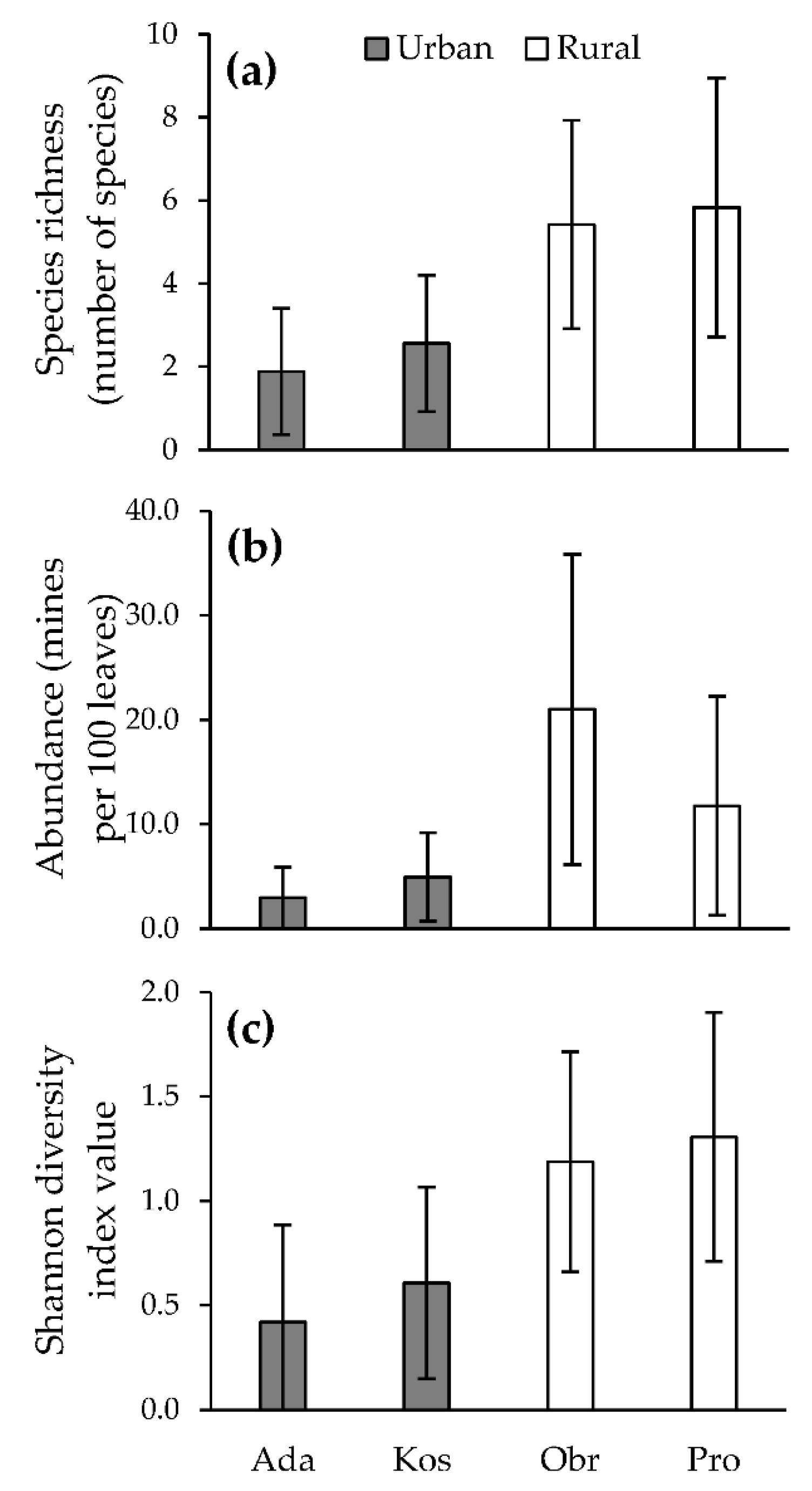
3.2. Leaf Miner Community and Individual Species of Leaf Miners in Urban and Rural Habitats
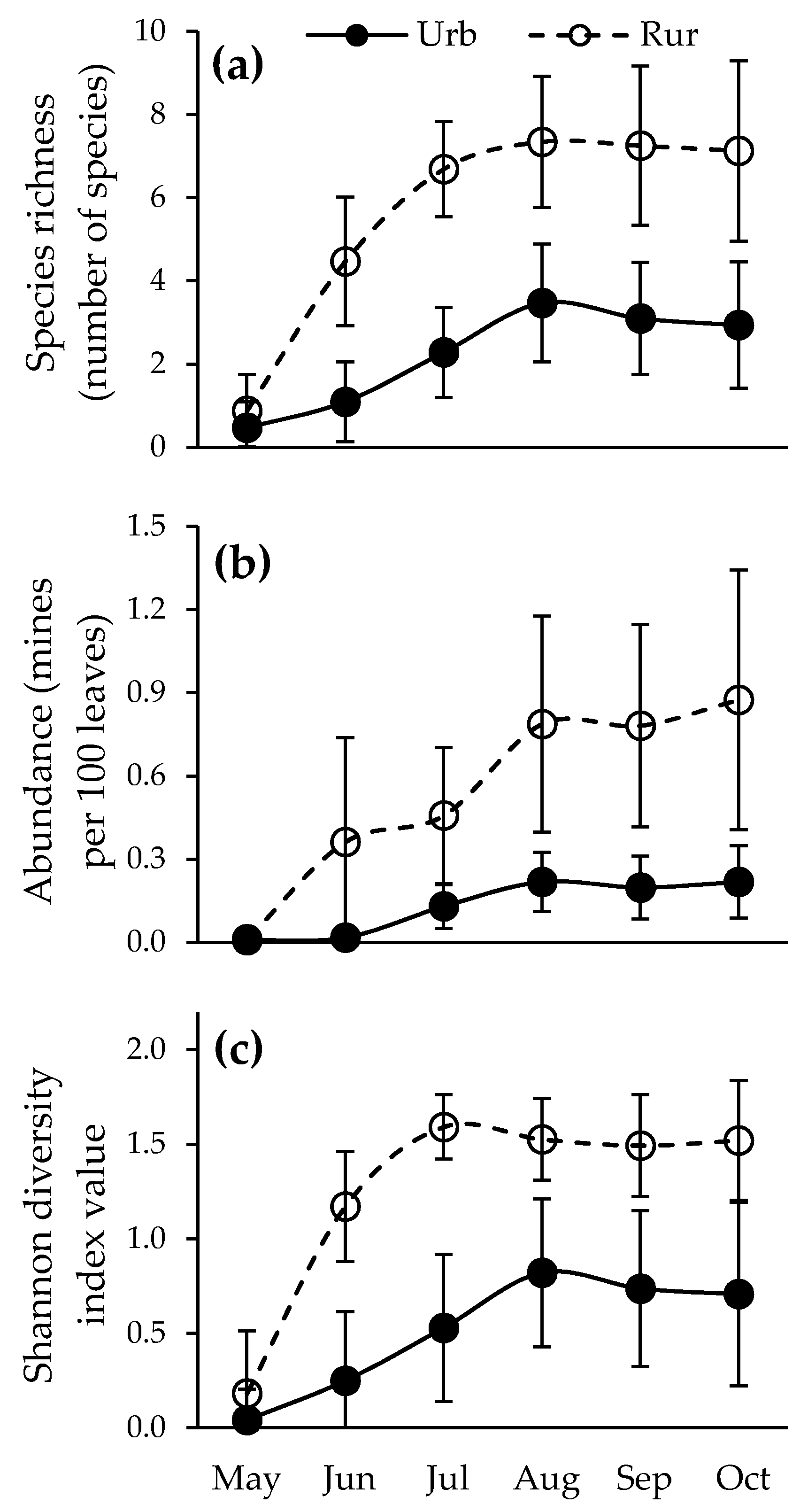
3.3. Seasonal Dynamics of Leaf Miners’ Species Richness, Abundance, and Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Primack, R. Where is the World’s Biological Diversity Found? In Essentials of Conservation Biology; Primack, R., Ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2014; pp. 51–68. ISBN 978-1605352893. [Google Scholar]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- DeFries, R.S.; Rudel, T.; Uriarte, M.; Hansen, M. Deforestation driven by urban population growth and agricultural trade in the twenty-first century. Nat. Geosci. 2010, 3, 178–181. [Google Scholar] [CrossRef]
- Eltom, I.M.; Elfaig, A.H.I.; Salih, A.A.M. Urban Development and Deforestation: Urban Development and Deforestation: Evidences from El-Obeid Town (1970–2010), Western Sudan. Int. J. Sci. Res. Publ. 2013, 3, 1–9. [Google Scholar]
- Sejati, A.W.; Buchori, I.; Rudiarto, I. The Impact of Urbanization to Forest Degradation in Metropolitan Semarang: A Preliminary Study. IOP Conf. Ser. Earth Environ. Sci. 2018, 123, 012011. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2020; FAO: Rome, Italy, 2020; ISBN 978-92-5-132974-0. [Google Scholar]
- Contreras-Hermosilla, A. The Underlying Causes of Forest Decline. CIFOR Occas. Pap. 2000, 30, 1–25. [Google Scholar]
- Wheeler, A.G. College Campuses: Patches of Insect Diversity, Opportunities for Entomological Discovery, and Means for Enhancing Ecological Literacy. Am. Entomol. 2008, 54, 18–35. [Google Scholar] [CrossRef] [Green Version]
- Rickman, J.K.; Connor, E.F. The effect of urbanization on the quality of remnant habitats for leaf-mining Lepidoptera on Quercus agrifolia. Ecography 2003, 26, 777–787. [Google Scholar] [CrossRef]
- Hunter, P. The human impact on biological diversity: How species adapt to urban challenges sheds light on evolution and provides clues about conservation. EMBO Rep. 2007, 8, 316–318. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.; Fang, Y.; Li, K. Impacts of urbanization process on insect diversity. Biodivers. Sci. 2013, 21, 260–268. [Google Scholar] [CrossRef]
- Su, Z.; Li, X.; Zhou, W.; Ouyang, Z. Effect of Landscape Pattern on Insect Species Density within Urban Green Spaces in Beijing, China. PLoS ONE 2015, 10, e0119276. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Mata, L.; Threlfall, C.G.; Williams, N.S.G.; Hahs, A.K.; Malipatil, M.; Stork, N.E.; Livesley, S.J. Conserving herbivorous and predatory insects in urban green spaces. Sci. Rep. 2017, 7, 40970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, T.H.; Wang, J.R.; Gu, J.Q.; Li, T.Q.; Lv, K.; Zhou, G.X.; Xu, Z.H. Effects of habitat fragmentation on the functional diversity of insects in Thousand Island Lake, China. Entomol. Res. 2019, 49, 93–104. [Google Scholar] [CrossRef]
- Moreira, X.; Abdala-Roberts, L.; Berny Mier y Teran, J.C.; Covelo, F.; de la Mata, R.; Francisco, M.; Hardwick, B.; Pires, R.M.; Roslin, T.; Schigel, D.S.; et al. Impacts of urbanization on insect herbivory and plant defences in oak trees. Oikos 2019, 128, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, D.L.; Pearse, I.S.; Baty, J.H. Drivers of specialist herbivore diversity across 10 cities. Landsc. Urban Plan. 2012, 108, 123–130. [Google Scholar] [CrossRef]
- McIntyre, N.E. Ecology of Urban Arthropods: A Review and a Call to Action. Ecol. Popul. Biol. 2000, 93, 825–835. [Google Scholar] [CrossRef]
- Eötvös, C.B.; Lövei, G.L.; Magura, T. Predation Pressure on Sentinel Insect Prey Along a Riverside Urbanization Gradient in Hungary. Insects 2020, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Stucky, B.J.; Deck, J.; Baiser, B.; Guralnick, R.P. The effect of urbanization on plant phenology depends on regional temperature. Nat. Ecol. Evol. 2019, 3, 1661–1667. [Google Scholar] [CrossRef]
- Jaworski, T.; Hilszczański, J. The effect of temperature and humidity changes on insects development their impact on forest ecosystems in the expected climate change. For. Res. Pap. 2014, 74, 345–355. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Kozlov, M.V.; Lanta, V.; Zverev, V.; Rainio, K.; Kunavin, M.A.; Zvereva, E.L. Decreased losses of woody plant foliage to insects in large urban areas are explained by bird predation. Glob. Chang. Biol. 2017, 23, 4354–4364. [Google Scholar] [CrossRef] [PubMed]
- Buczkowski, G.; Richmond, D.S. The Effect of Urbanization on Ant Abundance and Diversity: A Temporal Examination of Factors Affecting Biodiversity. PLoS ONE 2012, 7, e41729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntyre, N.E.; Rango, J.; Fagan, W.F.; Faeth, S.H. Ground arthropod community structure in a heterogeneous urban environment. Landsc. Urban Plan. 2001, 52, 257–274. [Google Scholar] [CrossRef]
- Eötvös, C.B.; Magura, T.; Lövei, G.L. A meta-analysis indicates reduced predation pressure with increasing urbanization. Landsc. Urban Plan. 2018, 180, 54–59. [Google Scholar] [CrossRef]
- Lee, C.M.; Park, J.W.; Kwon, T.S.; Kim, S.S.; Ryu, J.W.; Jung, S.J.; Lee, S.K. Diversity and density of butterfly communities in urban green areas: An analytical approach using GIS. Zool. Stud. 2015, 54, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Denys, C.; Schmidt, H. Insect communities on experimental mugwort (Artemisia vulgaris L.) plots along an urban gradient. Oecologia 1998, 113, 269–277. [Google Scholar] [CrossRef]
- Hering, E.M. Biology of the Leaf Miners; Dr W. Junk: Hague, The Netherlands, 1951. [Google Scholar]
- Hering, E.M. Bestimmungstabellen der Blattminen von Europa Einschliesslich des Mittelmeerbeckens und der Kanarishen Inseln, Band I, II und III; Dr W. Junk: Hague, The Netherlands, 1957; ISBN ISBN 9789061939818. [Google Scholar]
- Brändle, M.; Amarell, U.; Auge, H.; Klotz, S.; Brandl, R. Plant and insect diversity along a pollution gradient: Understanding species richness across trophic levels. Biodivers. Conserv. 2001, 10, 1497–1511. [Google Scholar] [CrossRef]
- Ali, J.G.; Agrawal, A.A. Specialist versus generalist insect herbivores and plant defense. Trends Plant Sci. 2012, 17, 293–302. [Google Scholar] [CrossRef]
- Nakadai, R. Species diversity of herbivorous insects: A brief review to bridge the gap between theories focusing on the generation and maintenance of diversity. Ecol. Res. 2017, 32, 811–819. [Google Scholar] [CrossRef]
- Weisser, W.; Siemann, E. The Various Effects of Insects on Ecosystem Functioning. In Insects and Ecosystem Function; Weisser, W., Siemann, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 3–24. ISBN 978-3-540-74004-9. [Google Scholar]
- Jones, E.L.; Leather, S.R. Invertebrates in urban areas: A review. Eur. J. Entomol. 2012, 109, 463–478. [Google Scholar] [CrossRef] [Green Version]
- Siddig, A.A.H.; Ellison, A.M.; Ochs, A.; Villar-leeman, C.; Lau, M.K. How do ecologists select and use indicator species to monitor ecological change? Insights from 14 years of publication in Ecological Indicators. Ecol. Indic. 2016, 60, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, B.; Bailey, S.; Vaughan, I.P.; Memmott, J. A comparison of clearfelling and gradual thinning of plantations for the restoration of insect herbivores and woodland plants. J. Appl. Ecol. 2015, 52, 1538–1546. [Google Scholar] [CrossRef] [Green Version]
- Dobrosavljevic, J.; Markovic, C.; Bojic, S. Overview of leaf miner fauna in Serbia. In Proceedings of the VIII International Agriculture Symposium “AGROSYM 2017”; Kovacevic, D., Ed.; University of East Sarajevo, Faculty of Agriculture, Republic of Srpska, Bosnia: Jahorina, Bosnia and Herzegovina, 2018; pp. 1490–1498. [Google Scholar]
- Dobrosavljević, J.; Marković, Č. Hinatara nigripes (Konow) i Heterarthrus flavicollis (Gussakovskij) (Hymenoptera, Tenthredinidae), nove vrste lisnih minera u fauni Srbije. In Proceedings of the XI Simpozijum Entomologa Srbije; Glavendekić, M., Ed.; Entomološko Društvo Srbije: Goč, Serbia, 2017; pp. 79–80. [Google Scholar]
- Dobrosavljević, J.N.; Marković, Č.; Stojanović, A. Contribution To the Knowledge of Phyllonorycter issikii (Kumata, 1963) (Lepidoptera, Gracillariidae) in Serbia. Acta Entomol. Serbica 2018, 23, 25–32. [Google Scholar] [CrossRef]
- Marković, Č.; Dobrosavljević, J.; Vujičić, P.; Cebeci, H.H. Impact of regeneration by shelterwood cutting on the pedunculate oak (Quercus robur) leaf mining insect community. Biologia 2020. [Google Scholar] [CrossRef]
- Simonović, M.; Gaora, D. Miner vinove loze, Phyllocnistis vitegenella Clemens (Lepidoptera: Gracillariidae)—Nova vrsta u Srbiji. Biljn. Lek. 2019, 47, 337–344. [Google Scholar]
- Raupp, M.J.; Shrewsbury, P.M.; Herms, D.A. Ecology of Herbivorous Arthropods in Urban Landscapes. Annu. Rev. Entomol. 2010, 55, 19–38. [Google Scholar] [CrossRef] [Green Version]
- Fenoglio, M.S.; Salvo, A.; Estallo, E.L. Effects of urbanisation on the parasitoid community of a leafminer. Acta Oecol. 2009, 35, 318–326. [Google Scholar] [CrossRef]
- Kahn, D.M.; Cornell, H.V. Leafminers, Early Leaf Abscission, and Parasitoids: A Tritrophic Interaction. Ecology 1989, 70, 1219–1226. [Google Scholar] [CrossRef]
- Koricheva, J.; Larsson, S.; Haukioja, E. Insect Performance on Experimentally Stressed Woody Plants: A Meta-Analysis. Annu. Rev. Entomol. 1998, 43, 195–216. [Google Scholar] [CrossRef] [Green Version]
- Oke, T.R. City size and the urban heat island. Atmos. Environ. 1973, 7, 769–779. [Google Scholar] [CrossRef]
- Mulieri, P.R.; Patitucci, L.D.; Schnack, J.A.; Mariluis, J.C. Diversity and seasonal dynamics of an assemblage of sarcophagid diptera in a gradient of urbanization. J. Insect Sci. 2011, 11, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magura, T.; Lövei, G.L. The permeability of natural versus anthropogenic forest edges modulates the abundance of ground beetles of different dispersal power and habitat affinity. Diversity 2020, 12, 320. [Google Scholar] [CrossRef]
- Eaton, E.; Caudullo, G.; Oliveira, S.; de Rigo, D. Quercus robur and Quercus petraea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 160–163. ISBN 978-92-79-52833-0. [Google Scholar]
- Annighöfer, P.; Beckschäfer, P.; Vor, T.; Ammer, C. Regeneration Patterns of European Oak Species (Quercus petraea (Matt.) Liebl., Quercus robur L.) in Dependence of Environment and Neighborhood. PLoS ONE 2015, 10, e0134935. [Google Scholar] [CrossRef] [PubMed]
- Quine, C.; Atkinson, N.; Denman, S.; Desprez-loustau, L.; Jackson, R.; Kirby, K. Action Oak Knowledge Review: An Assessment of the Current Evidence on Oak Health, Identification of Evidence Gaps and Prioritisation of Research Needs; Atkinson, N., Desprez-Loustau, M.-L., Kirby, K., Denman, S., Jackson, R., Quine, C., Eds.; Action Oak: Haslemere, UK, 2019; ISBN 978-1-5272-4193-0. [Google Scholar]
- Pearson, D.L. Selecting indicator taxa for the quantitative assessment of biodiversity. Biodivers. Meas. Estim. 1995, 345, 75–79. [Google Scholar]
- Barantal, S.; Castagneyrol, B.; Durka, W.; Iason, G.; Morath, S.; Koricheva, J. Contrasting effects of tree species and genetic diversity on the leaf-miner communities associated with silver birch. Oecologia 2019, 189, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Ellis, W.N. Leafminers and Plant Galls of Europe. Available online: http://www.bladmineerders.nl/ (accessed on 20 October 2019).
- Baraniak, E.; Walczak, U.; Tryjanowski, P.; Zduniak, P. Effect of distance between host trees and leaf litter removal on population density of Cameraria ohridella Deschka & Dimic, 1986 (Lepidoptera, Gracillariidae)—Pest of chestnut (Aesculus sp.) trees. Pol. J. Ecol. 2004, 52, 569–574. [Google Scholar]
- Jovicich, E. Leafminer Pest-Generic Incursion Management Plan for the Australian Vegetable Industry; Horticulture Australia Ltd.: Sydney, Australia, 2009; ISBN 0-7341-2154-7. Available online: http://era.daf.qld.gov.au/id/eprint/2539/ (accessed on 16 March 2017).
- Da Rocha, J.M.R.; Almeida, J.R.; Lins, G.A.; Durval, A. Insects as indicators of environmental changing and pollution: A review of appropriate species and their monitoring. Holos Environ. 2010, 10, 250–262. [Google Scholar] [CrossRef]
- Gerlach, J.; Samways, M.; Pryke, J. Terrestrial invertebrates as bioindicators: An overview of available taxonomic groups. J. Insect Conserv. 2013, 17, 831–850. [Google Scholar] [CrossRef]
- Republic Hydrometeorological Service of Serbia. Available online: http://www.hidmet.gov.rs (accessed on 13 May 2020).
- Forest Directorate of Ministry of Agriculture Forestry and Water Management. Available online: https://upravazasume.gov.rs (accessed on 13 May 2020).
- Patočka, J.; Turčani, M. Lepidoptera Pupae. Central European Species; Apollo Books: Stenstrup, Denmark, 2005; ISBN 9788788757477. [Google Scholar]
- Laštůvka, A.; Zdeněk, L.; Liška, J.; Šumpich, J. Motýli a Housenky Střední Evropy V., Drobní Motýli I; Academia, Praha: Prague, Czech Republic, 2018; ISBN 978-80-200-2852-5. [Google Scholar]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Tack, A.; Ovaskainen, O.; Pulkkinen, P.; Roslin, T. Spatial location dominates over host plant genotype in structuring an herbivore community. Ecology 2010, 91, 2660–2672. [Google Scholar] [CrossRef]
- Gripenberg, S.; Ovaskainen, O.; Elly, M.; Roslin, T. Spatial population structure of a specialist leaf-mining moth. J. Anim. Ecol. 2008, 77, 757–767. [Google Scholar] [CrossRef]
- Bañuelos, M.J.; Kollmann, J. Effects of host-plant population size and plant sex on a specialist leaf-miner. Acta Oecol. 2011, 37, 58–64. [Google Scholar] [CrossRef]
- Valdés-Correcher, E.; van Halder, I.; Barbaro, L.; Castagneyrol, B.; Hampe, A. Insect herbivory and avian insectivory in novel native oak forests: Divergent effects of stand size and connectivity. For. Ecol. Manag. 2019, 445, 146–153. [Google Scholar] [CrossRef]
- Jactel, H.; Brockerhoff, E.G. Tree diversity reduces herbivory by forest insects. Ecol. Lett. 2007, 10, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Moreira, X.; Abdala-Roberts, L.; Rasmann, S.; Castagneyrol, B.; Mooney, K.A. Plant diversity effects on insect herbivores and their natural enemies: Current thinking, recent findings, and future directions. Curr. Opin. Insect Sci. 2016, 14, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Long, C.; Xu, J.; Guo, Q.; Zhihong, Z.; Bater, Z. Are dominant plant species more susceptible to leaf—Mining insects? A case study at Saihanwula Nature Reserve, China. Ecol. Evol. 2018, 8, 7633–7648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, M.E.; Petersen, M.J. Extending the ‘resource concentration hypothesis’ to the landscape-scale by considering dispersal mortality and fitness costs. Agric. Ecosyst. Environ. 2017, 249, 1–3. [Google Scholar] [CrossRef]
- Pincebourde, S.; Woods, H.A. Climate uncertainty on leaf surfaces: The biophysics of leaf microclimates and their consequences for leaf-dwelling organisms. Funct. Ecol. 2012, 26, 844–853. [Google Scholar] [CrossRef]
- Sinclair, R.J.; Hughes, L. Leaf miners: The hidden herbivores. Austral Ecol. 2010, 35, 300–313. [Google Scholar] [CrossRef]
- Preszler, R.W.; Price, P.W. A Test of Plant-Vigor, Plant-Stress, and Plant-Genotype Effects on Leaf-Miner Oviposition and Performance. Oikos 1995, 74, 485–495. [Google Scholar] [CrossRef]
- Bairstow, K.A.; Clarke, K.L.; McGeoch, M.A.; Andrew, N.R. Leaf miner and plant galler species richness on Acacia: Relative importance of plant traits and climate. Oecologia 2010, 163, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Krstić, M.R.; Kanjevac, B.R.; Babić, V.P. Effects of extremely high temperatures on some growth parameters of sessile oak (Quercus petraea/Matt./Liebl.) seedlings in northeastern Serbia. Arch. Biol. Sci. 2018, 70, 521–529. [Google Scholar] [CrossRef]
- Faeth, S. Novel aspects of host tree resistance to leafminers. In Forest Insect Guilds: Patterns of Interaction with Host Trees; Baranchikov, Y.N., Mattson, W.J., Hain, F.P., Eds.; General Technical Report NE-153; Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA, 1991; pp. 219–239. [Google Scholar]
- Southwood, R.T.E.; Wint, W.G.R.; Kennedy, C.E.J.; Greenwood, S.R. Seasonality, abundance, species richness and specificity of the phytophagous guild of insects on oak (Quercus) canopies. Eur. J. Entomol. 2004, 101, 43–50. [Google Scholar] [CrossRef]
- Muiruri, E.W.; Barantal, S.; Iason, G.R.; Salminen, J.P.; Perez-Fernandez, E.; Koricheva, J. Forest diversity effects on insect herbivores: Do leaf traits matter? New Phytol. 2019, 221, 2250–2260. [Google Scholar] [CrossRef] [Green Version]
- Castagneyrol, B.; Giffard, B.; Péré, C.; Jactel, H. Plant apparency, an overlooked driver of associational resistance to insect herbivory. J. Ecol. 2013, 101, 418–429. [Google Scholar] [CrossRef]
- Valdés-Correcher, E.; Bourdin, A.; González-Martínez, S.C.; Moreira, X.; Galmán, A.; Castagneyrol, B.; Hampe, A. Leaf chemical defences and insect herbivory in oak: Accounting for canopy position unravels marked genetic relatedness effects. Ann. Bot. 2020, 126, 865–872. [Google Scholar] [CrossRef]
- Valencia-Cuevas, L.; Tovar-Sánchez, E. Oak Canopy arthropod communities: Which factors shape its structure? Rev. Chil. Hist. Nat. 2015, 88. [Google Scholar] [CrossRef]
- Shimadzu, H.; Dornelas, M.; Henderson, P.A.; Magurran, A.E. Diversity is maintained by seasonal variation in species abundance. BMC Biol. 2013, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lemieux, J.; Cusson, M. Effects of Habitat-Forming Species Richness, Evenness, Identity, and Abundance on Benthic Intertidal Community Establishment and Productivity. PLoS ONE 2014, 9, e109261. [Google Scholar] [CrossRef] [Green Version]
- Bellamy, A.S.; Svensson, O.; van den Brink, P.J.; Gunnarsson, J.; Tedengren, M. Insect community composition and functional roles along a tropical agricultural production gradient. Environ. Sci. Pollut. Res. 2018, 25, 13426–13438. [Google Scholar] [CrossRef] [Green Version]
- Tsafack, N.; Di Biase, L.; Xie, Y.; Wang, X.; Fattorini, S. Carabid community stability is enhanced by carabid diversity but reduced by aridity in Chinese steppes. Acta Oecol. 2019, 99, 103450. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Order | Family | Species | Abundance | Significance of the Influence of the Habitat Type | |
|---|---|---|---|---|---|
| Urban | Rural | ||||
| Coleoptera | Curculionidae | Orchestes erythropus (Germ., 1821) | not present | 0.003 ± 0.040 | |
| O. pilosus (Fabr., 1781) | not present | 0.002 ± 0.023 | |||
| O. quercus (L., 1758) | not present | 0.008 ± 0.057 | |||
| O. subfasciatus Gyll., 1835 | not present | 0.019 ± 0.094 | |||
| Hymenoptera | Tenthredinidae | Profenusa pygmaea(Klug, 1816) | 0.080 ± 0.208 | 0.671 ± 1.267 | Z = −6.81 * |
| Lepidoptera | Bucculatricidae | Bucculatrix ulmella Zell., 1848 | 0.028 ± 0.141 | 0.131 ± 0.333 | Z = −2.56 * |
| Coleophora flavipennella (Dup., 1843) | not present | 0.003 ± 0.039 | |||
| C. ibipennella Zell., 1849 | not present | 0.004 ± 0.043 | |||
| C. kuehnella (Goeze, 1783) | not present | 0.007 ± 0.057 | |||
| Eriocraniidae | Dyseriocrania subpurpurella (Haw., 1828) | 0.011 ± 0.069 | 0.022 ± 0.113 | ||
| Gracillariidae | Acrocercops brongniardella (Fabr., 1798) | 0.021 ± 0.221 | 0.007 ± 0.071 | ||
| Caloptilia alchimiella (Scop., 1763) | 0.303 ± 0.630 | 2.704 ± 3.469 | Z = −9.66 * | ||
| Phyllonorycter harrisella (L., 1761) | 2.657 ± 2.673 | 1.185 ± 1.364 | Z = 5.12 | ||
| Ph. lautella (Zell., 1846) | not present | 0.015 ± 0.104 | |||
| Ph. messaniella (Zell., 1846) | 0.010 ± 0.103 | 0.034 ± 0.171 | |||
| Ph. muelleriella (Zell., 1839) | not present | 0.171 ± 0.534 | n/a | ||
| Ph. quercifoliella (Zell., 1839) | 0.187 ± 0.440 | 0.383 ± 0.768 | Z = −2.13 * | ||
| Ph. roboris (Zell., 1839) | 0.206 ± 0.549 | 2.981 ± 4.214 | Z * = −9.58 * | ||
| Heliozelidae | Heliozela sericiella (Haw., 1828) | 0.006 ± 0.058 | 0.014 ± 0.075 | ||
| Nepticulidae | Ectoedemia albifasciella (Hein., 1871) | not present | 0.005 ± 0.043 | ||
| E. caradjai (Gros., 1944) | 0.021 ± 0.122 | 2.021 ± 4.171 | Z = −6.20 * | ||
| E. quinquella (Bed., 1848) | not present | 0.013 ± 0.122 | |||
| E. subbimaculella (Haw., 1828) | not present | 0.034 ± 0.257 | |||
| Stigmella atricapitella (Haw., 1828) | 0.033 ± 0.189 | 0.009 ± 0.083 | |||
| S. basiguttella (Hein., 1862) | 0.004 ± 0.053 | 0.333 ± 0.758 | Z = −5.03 * | ||
| S. roborella (Johan., 1971) | 0.006 ± 0.067 | 0.021 ± 0.132 | |||
| S. ruficapitella (Haw., 1828) | 0.018 ± 0.146 | 0.010 ± 0.089 | |||
| Tischeriidae | Tischeria decidua Wck., 1876 | 0.050 ± 0.196 | 0.316 ± 0.577 | Z = −4.57 * | |
| T.dodonaea Stt., 1858 | 0.109 ± 0.595 | 0.094 ± 0.280 | Z = −0.43 ns | ||
| T. ekebladella (Bjerk., 1795) | 0.189 ± 0.461 | 5.155 ± 6.123 | Z = −12.37 * | ||
| Analyzed Parameter | Rank Sum Urban Habitat | Rank Sum Rural Habitat | U | Z | p-Level | Valid N Urban Habitat | Valid N Rural Habitat |
|---|---|---|---|---|---|---|---|
| Species richness | 25,491.5 | 48,428.5 | 6963.5 | −10.546 | 0.000 | 192 | 192 |
| Abundance | 26,995.5 | 46,924.5 | 8467.5 | −9.163 | 0.000 | 192 | 192 |
| Diversity | 24,828.5 | 49,091.5 | 6300.5 | −11.155 | 0.000 | 192 | 192 |
| Analyzed Parameter | Month | Rank Sum Urban Habitat | Rank Sum Rural Habitat | U | Z | p-Level | Valid N Urban Habitat | Valid N Rural Habitat | 2 * 1 Sided Exact p |
|---|---|---|---|---|---|---|---|---|---|
| Species Richness | May | 910.0 | 1170.0 | 382.0 | −1.746 | 0.081 | 32 | 32 | 0.082 |
| June | 551.5 | 1528.5 | 23.5 | −6.559 | 0.000 | 32 | 32 | 0.000 | |
| July | 528.0 | 1552.0 | 0.0 | −6.875 | 0.000 | 32 | 32 | 0.000 | |
| August | 660.5 | 1419.5 | 132.5 | −5.096 | 0.000 | 32 | 32 | 0.000 | |
| September | 631.5 | 1448.5 | 103.5 | −5.485 | 0.000 | 32 | 32 | 0.000 | |
| October | 609.5 | 1470.5 | 81.5 | −5.780 | 0.000 | 32 | 32 | 0.000 | |
| Abundance | May | 938.5 | 1141.5 | 410.5 | −1.363 | 0.173 | 32 | 32 | 0.174 |
| June | 550.0 | 1530.0 | 22.0 | −6.579 | 0.000 | 32 | 32 | 0.000 | |
| July | 633.0 | 1447.0 | 105.0 | −5.465 | 0.000 | 32 | 32 | 0.000 | |
| August | 652.0 | 1428.0 | 124.0 | −5.210 | 0.000 | 32 | 32 | 0.000 | |
| September | 625.0 | 1455.0 | 97.0 | −5.572 | 0.000 | 32 | 32 | 0.000 | |
| October | 594.5 | 1485.5 | 66.5 | −5.982 | 0.000 | 32 | 32 | 0.000 | |
| Diversity | May | 941.5 | 1138.5 | 413.5 | −1.323 | 0.186 | 32 | 32 | 0.187 |
| June | 557.5 | 1522.5 | 29.5 | −6.479 | 0.000 | 32 | 32 | 0.000 | |
| July | 532.0 | 1548.0 | 4.0 | −6.821 | 0.000 | 32 | 32 | 0.000 | |
| August | 634.0 | 1446.0 | 106.0 | −5.451 | 0.000 | 32 | 32 | 0.000 | |
| September | 626.5 | 1453.5 | 98.5 | −5.552 | 0.000 | 32 | 32 | 0.000 | |
| October | 652.5 | 1427.5 | 124.5 | −5.203 | 0.000 | 32 | 32 | 0.000 |
| Analyzed Parameter | Habitat Type | N | Df | Chi Sqr. | p |
|---|---|---|---|---|---|
| Species richness | Urban | 32 | 5 | 97.154 | 0.000 |
| Rural | 94.391 | 0.000 | |||
| Abundance | Urban | 122.455 | 0.000 | ||
| Rural | 109.589 | 0.000 | |||
| Diversity | Urban | 65.428 | 0.000 | ||
| Rural | 89.459 | 0.000 |
| Species | May | Jun | Jul | Aug | Sep | Oct | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| U | R | U | R | U | R | U | R | U | R | U | R | |
| Orchestes erythropus (Germ., 1821) | + | |||||||||||
| O. pilosus (Fabr., 1781) | + | |||||||||||
| O. quercus (L., 1758) | + | + | ||||||||||
| O. subfasciatus Gyll., 1835 | + | + | + | |||||||||
| Profenusa pygmaea (Klug, 1816) | + | + | + | + | + | + | + | + | + | + | + | + |
| Bucculatrix ulmella Zell., 1848 | + | + | + | + | + | + | + | + | + | |||
| Coleophora flavipennella (Dup., 1843) | + | |||||||||||
| C. ibipennella Zell., 1849 | + | |||||||||||
| C. kuehnella (Goeze, 1783) | + | + | + | |||||||||
| Dyseriocrania subpurpurella (Haw., 1828) | + | + | + | + | + | + | ||||||
| Acrocercops brongniardella (Fabr., 1798) | + | + | + | |||||||||
| Caloptilia alchimiella (Scop., 1763) | + | + | + | + | + | + | + | + | + | + | ||
| Phyllonorycter harrisella (L., 1761) | + | + | + | + | + | + | + | + | + | + | + | + |
| Ph. lautella (Zell., 1846) | + | + | ||||||||||
| Ph. messaniella (Zell., 1846) | + | + | ||||||||||
| Ph. muelleriella (Zell., 1839) | + | + | + | + | + | |||||||
| Ph. quercifoliella (Zell., 1839) | + | + | + | + | + | + | + | + | + | + | + | + |
| Ph. roboris (Zell., 1839) | + | + | + | + | + | + | + | + | + | + | ||
| Heliozela sericiella (Haw., 1828) | + | + | + | + | + | |||||||
| Ectoedemia albifasciella (Hein., 1871) | + | + | ||||||||||
| E. caradjai (Gros., 1944) | + | + | + | + | + | + | ||||||
| E. quinquella (Bed., 1848) | + | |||||||||||
| E. subbimaculella (Haw., 1828) | + | |||||||||||
| Stigmella atricapitella (Haw., 1828) | + | + | + | + | ||||||||
| S. basiguttella (Hein., 1862) | + | + | + | + | + | + | ||||||
| S. roborella (Johan., 1971) | + | + | + | + | + | + | ||||||
| S. ruficapitella (Haw., 1828) | + | + | + | + | + | |||||||
| Tischeria decidua Wck., 1876 | + | + | + | + | + | + | + | + | + | |||
| T.dodonaea Stt., 1858 | + | + | + | + | + | + | + | |||||
| T. ekebladella (Bjerk., 1795) | + | + | + | + | + | + | + | + | + | |||
| Total species per habitat type per month | 5 | 12 | 6 | 17 | 11 | 19 | 11 | 16 | 12 | 16 | 14 | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobrosavljević, J.; Marković, Č.; Marjanović, M.; Milanović, S. Pedunculate Oak Leaf Miners’ Community: Urban vs. Rural Habitat. Forests 2020, 11, 1300. https://doi.org/10.3390/f11121300
Dobrosavljević J, Marković Č, Marjanović M, Milanović S. Pedunculate Oak Leaf Miners’ Community: Urban vs. Rural Habitat. Forests. 2020; 11(12):1300. https://doi.org/10.3390/f11121300
Chicago/Turabian StyleDobrosavljević, Jovan, Čedomir Marković, Marija Marjanović, and Slobodan Milanović. 2020. "Pedunculate Oak Leaf Miners’ Community: Urban vs. Rural Habitat" Forests 11, no. 12: 1300. https://doi.org/10.3390/f11121300
APA StyleDobrosavljević, J., Marković, Č., Marjanović, M., & Milanović, S. (2020). Pedunculate Oak Leaf Miners’ Community: Urban vs. Rural Habitat. Forests, 11(12), 1300. https://doi.org/10.3390/f11121300