Agents Affecting the Productivity of Pine Plantations on the Loess Plateau in China: A Study Based on Structural Equation Modeling

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection and Analysis
2.3. Selection of Variables
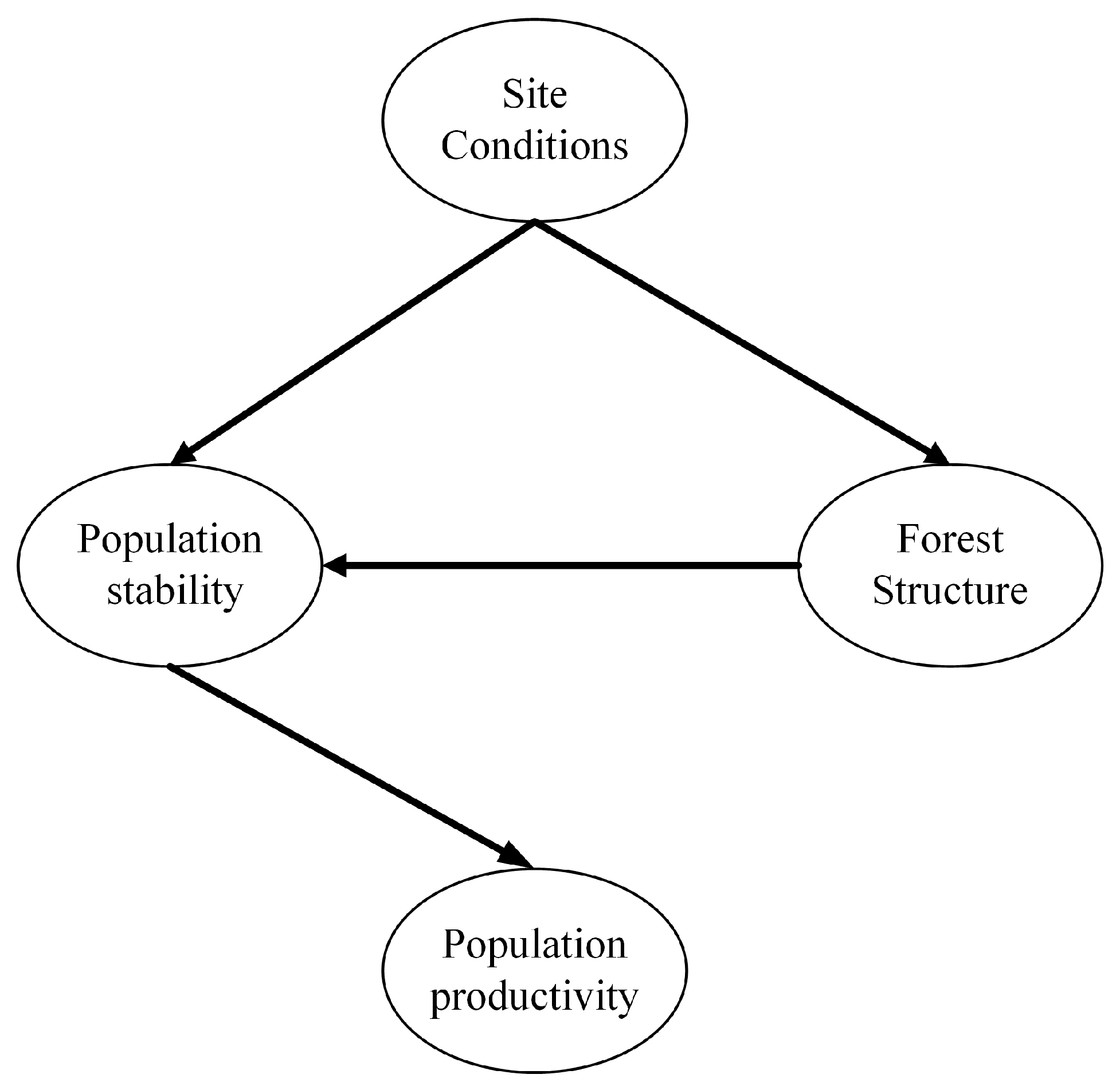
2.4. Construction of the Initial Model
2.5. Canonical Correlation Analysis
2.6. Model Suitability Test
3. Results
3.1. Canonical Correlation Analysis
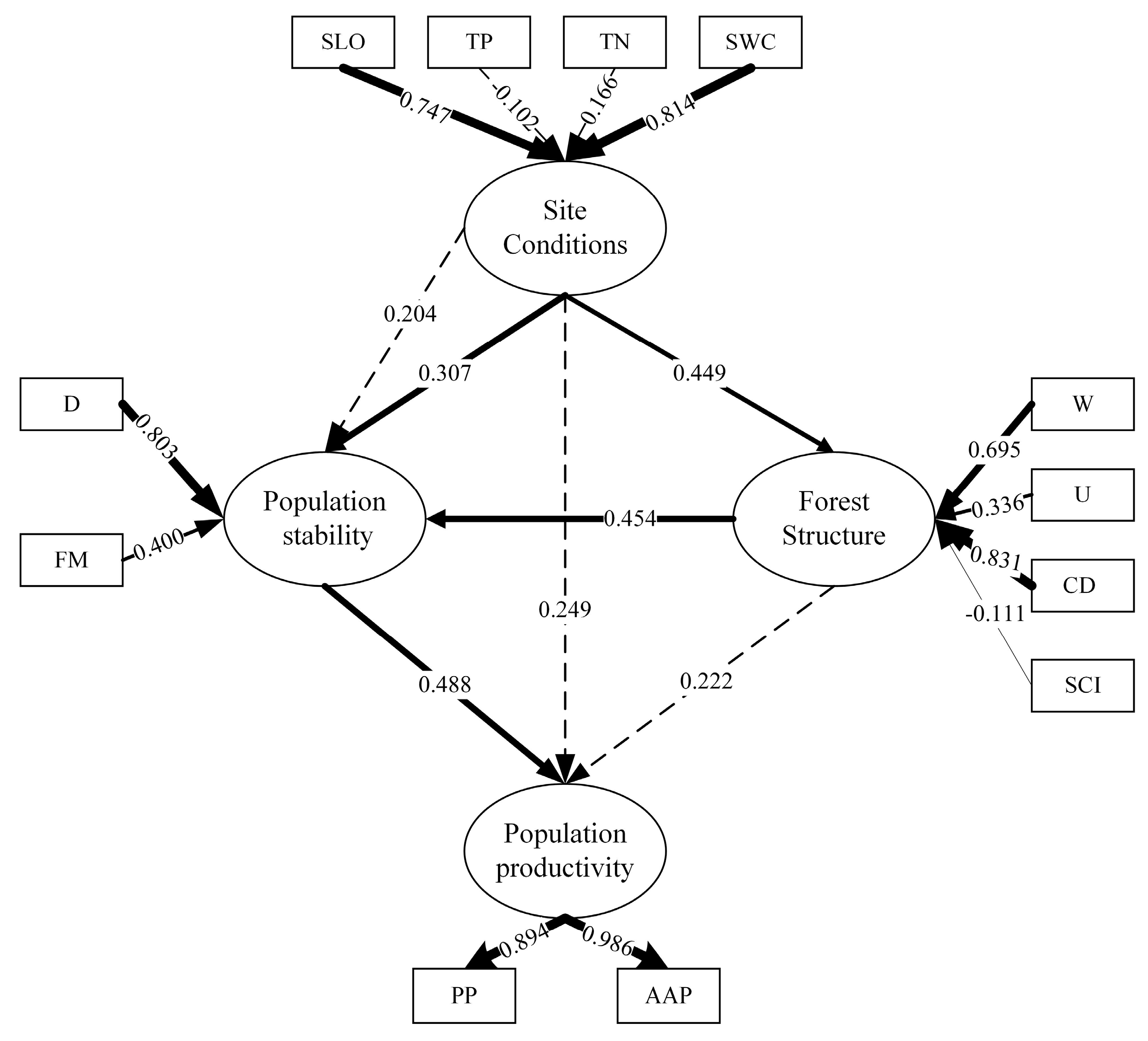
3.2. Modification and Interpretation of the Model
3.3. Model Fit Test
3.4. Relationships between Latent Variables
4. Discussion
4.1. Direct Effect of Population Stability on Population Productivity
4.2. Indirect Effect of Site Conditions on Population Productivity
4.3. Population Productivity and Stability are Differently Affected by Forest Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- National Forestry and Grassland Administration. 2014–2018 China Forest Resources Report; China Forestry Publishing House: Beijing, China, 2019; pp. 2–3. [Google Scholar]
- Wang, Q.; Wang, S.; Yu, X. Decline of soil fertility during forest conversion of secondary forest to Chinese fir plantations in Subtropical China. Land Degrad. Dev. 2011, 22, 444–452. [Google Scholar] [CrossRef]
- Liu, S.; Li, X.; Niu, L. The degradation of soil fertility in pure larch plantations in the northeastern part of China. Ecol. Eng. 1998, 10, 75–86. [Google Scholar] [CrossRef]
- Changshun, Z. Advance in Research on Soil Degradation and Soil Improvement of Timber Plantations. World For. Res. 2005, 18, 17–21. [Google Scholar]
- Chen, K.; Abbott, R.; Milne, R.; Tian, X.-M.; Liu, J. Phylogeography of Pinus tabulaeformis Carr. (Pinaceae), a dominant species of coniferous forest in northern China. Mol. Ecol. 2008, 17, 4276–4288. [Google Scholar] [CrossRef]
- Tao, W.U.; Hui, Z.W.; Chang, L.Y.; Hui, F.S. Effects of Different Forest Practices on Pinus tabulaeformis Population Numbers and Species Diversity in the Forest Region of Huanglongshan Mountain. Acta Bot. Boreali-Occident. Sin. 2006, 26, 1007–1013. [Google Scholar]
- Steffen, W.; Noble, I.; Canadell, J.; Apps, M.; Schulze, E.D.; Jarvis, P.; Baldocchi, D.; Ciais, P.; Cramer, W.; Ehleringer, J.; et al. The Terrestrial Carbon Cycle: Implications for the Kyoto Protocol. Science 1998, 280, 1393–1394. [Google Scholar]
- Norris, M.; Blair, J.; Johnson, L.; McKane, R. Assessing changes in biomass, productivity, and C and N stores following Juniperus virginiana forest expansion into tallgrass prairie. Can. J. For. Res. 2001, 31, 1940–1946. [Google Scholar] [CrossRef] [Green Version]
- Cousens, J.; Newbould, P. Methods of Estimating the Primary Production of Forests. J. Appl. Ecol. 1968, 5, 745. [Google Scholar] [CrossRef] [Green Version]
- Glatthorn, J.; Feldmann, E.; Pichler, V.; Hauck, M.; Leuschner, C. Biomass Stock and Productivity of Primeval and Production Beech Forests: Greater Canopy Structural Diversity Promotes Productivity. Ecosystems 2017. [Google Scholar] [CrossRef]
- Grimm, V.; Schmidt, E.; Wissel, C. On the application of stability concepts in ecology. Ecol. Model. 1992, 63, 143–161. [Google Scholar] [CrossRef]
- Hardiman, B.; Bohrer, G.; Gough, C.; Vogel, C.; Curtisi, P. The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest. Ecology 2011, 92, 1818–1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, C.; Falk, D.; Lynch, A.; Swetnam, T.; Wilcox, C. Disturbance and productivity interactions mediate stability of forest composition and structure. Ecol. Appl. 2016, 27. [Google Scholar] [CrossRef]
- Bohn, F.; Huth, A. The importance of forest structure to biodiversity–productivity relationships. R. Soc. Open Sci. 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dănescu, A.; Albrecht, A.; Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 2016, 182, 319–333. [Google Scholar] [CrossRef]
- Díaz-Yáñez, O.; Mola-Yudego, B.; Olabarria, J.; Pukkala, T. How does forest composition and structure affect the stability against wind and snow? For. Ecol. Manag. 2017, 401, 215–222. [Google Scholar] [CrossRef]
- Bourdier, T.; Cordonnier, T.; Kunstler, G.; Piedallu, C.; Lagarrigues, G.; Courbaud, B. Tree Size Inequality Reduces Forest Productivity: An Analysis Combining Inventory Data for Ten European Species and a Light Competition Model. PLoS ONE 2016, 11, e0151852. [Google Scholar] [CrossRef]
- Carnaval, A.; Hickerson, M.; Haddad, C.; Rodrigues, M.; Moritz, C. Stability Predicts Genetic Diversity in the Brazilian Atlantic Forest Hotspot. Science 2009, 323, 785–789. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Fernández, B.; Campos Arce, J.; Finegan, B.; Alvarado-Hernandez, A. Factors affecting site productivity of a Costa Rican secondary rain forest in relation to Vochysia ferruginea, a commercially valuable canopy tree species. For. Ecol. Manag. 1999, 118, 73–81. [Google Scholar] [CrossRef]
- Yadav, V.; Mueller, K.L.; Dragoni, D.; Michalak, A. A geostatistical synthesis study of factors affecting gross primary productivity in various ecosystems of North America. Biogeosci. Discuss. 2010, 7. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef]
- Bouriaud, O.; Marin, G.; Bouriaud, L.; Hessenmöller, D.; Ernst Detlef, S. Romanian legal management rules limit wood production in Norway spruce and beech forests. For. Ecosyst. 2016, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bueis, T.; Bravo, F.; Pando, V.; Turrión, M.B. Relationship between environmental parameters and Pinus sylvestris L. site index in forest plantations in northern Spain acidic plateau. IForest Biogeosci. For. 2016, 9, e1–e8. [Google Scholar] [CrossRef] [Green Version]
- Doncaster, C. Structural Equation Modeling and Natural Systems. Fish Fish. 2007, 8, 368–369. [Google Scholar] [CrossRef]
- Laughlin, D.; Abella, S.; Covington, W.; Grace, J. Species richness and soil properties in Pinus ponderosa forests: A structural equation modeling analysis. J. Veg. Sci. 2007, 18. [Google Scholar] [CrossRef]
- Watt, M.; Kimberley, M.; Dash, J.; Harrison, D. Spatial prediction of optimal final stand density for even age plantation forests using productivity indices. Can. J. For. Res. 2016, 47. [Google Scholar] [CrossRef]
- Aakala, T.; Fraver, S.; D’Amato, A.; Palik, B. Influence of competition and age on tree growth in structurally complex old-growth forests in northern Minnesota, USA. For. Ecol. Manag. 2013, 308, 128–135. [Google Scholar] [CrossRef]
- Kershaw, J.A., Jr.; Ducey, M.J.; Beers, T.W.; Husch, B. Forest Mensuration; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Aj, W.; Black, I. An Examination of the Degtjareff Method for Determining Soil Organic Matter, and A Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis, Part 3; American Society of Agronomy-Soil Science Society of America: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Parkinson, J.; Allen, S. A Wet Oxidation Procedure for Determination of Nitrogen and Mineral Nutrients in Biological Material. Communications in Soil Science and Plant Analysis. Commun. Soil Sci. Plant Anal. 1975, 6, 1–11. [Google Scholar] [CrossRef]
- Hui, G.; Gadow, K.; Hu, Y.B. The optimum standard angle of the uniform angle index. For. Res. 2004, 17, 687–692. [Google Scholar]
- Hui, G.; Gadow, K.; Albert, M. A new parameter for stand spatial structure—Neighbourhood comparison. Forest Research. 1999, 12, 1–6. [Google Scholar]
- Hui, G.; Albert, M.; Gadow, K. The measure of neighbourhood dimensions as a parameter to reproduce stand structures. Forstwissensch. Cent. 1998, 117, 258–266. [Google Scholar] [CrossRef]
- Hu, Y.B.; Hui, G.Y. How to describe the crowding degree of trees based on the relationship of neighboring trees. J. Beijing For. Univ. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- Hegyi, F. A simulation model for managing jack-pine standssimulation. R. Res. Notes 1974, 30, 74–90. [Google Scholar]
- Hui, G.; Wang, Y.; Zhang, G.; Zhao, Z.; Bai, C.; Liu, W. A novel approach for assessing the neighborhood competition in two different aged forests. For. Ecol. Manag. 2018, 422, 49–58. [Google Scholar] [CrossRef]
- Gadow, K. Characterizing Forest Spatial Structure and Diversity; University of Lund: Lund, Sweden, 2002; pp. 20–30. [Google Scholar]
- Putz, F.E.; Chan, H.T. Tree growth, dynamics, and productivity in a mature mangrove forest in Malaysia. For. Ecol. Manag. 1986, 17, 211–230. [Google Scholar] [CrossRef]
- Wong, K.K. Partial Least Squares Structural Equation Modeling (PLS-SEM) Techniques Using SmartPLS. Mark. Bull. 2013, 24, 1–32. [Google Scholar]
- Wong, K. Handling small survey sample size and skewed dataset with partial least square path modelling. Vue Mag. Mark. Res. Intell. Assoc. 2010, 20, 20–23. [Google Scholar]
- Bacon, L.D. Using LISREL and PLS to measure customer satisfaction. In Proceedings of the Sawtooth Software Conference Proceedings, La Jolla, CA, USA, 2–5 February 1999; pp. 305–306. [Google Scholar]
- Wang, B.; Yuankun, B.; Li, Y.; Li, W.; Zhao, P.; Yang, Y.; Qi, N.; Gou, R. Quantifying the Relationship among Impact Factors of Shrub Layer Diversity in Chinese Pine Plantation Forest Ecosystems. Forests 2019, 10, 781. [Google Scholar] [CrossRef] [Green Version]
- Hotelling, H. Relations between Two Sets of Variates. Biometrika 1935, 28, 321–377. [Google Scholar] [CrossRef]
- Xiangdong, L. Indicators on Structural Diversity Within-Stand: A Review. Sci. Silv. Sin. 2002, 38, 140–146. [Google Scholar]
- Anderson, J.; Gerbing, D. Structural Equation Modeling in Practice: A Review of Recommended Two-Step Approach. Psychological Bulletin Psychol. Bull. 1988, 103, 411–423. [Google Scholar] [CrossRef]
- Hair, J.F.; Ringle, C.; Sarstedt, M. PLS-SEM: Indeed a silver bullet. J. Mark. Theory Pract. 2011, 19, 139–151. [Google Scholar] [CrossRef]
- Hair, J.; Anderson, R.; Tatham, R.; Black, W. Multivariate Data Analysis, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1998. [Google Scholar]
- Fornell, C.; Larcker, D. Evaluating Structural Equation Models with Unobservable Variables and Measurement Error. J. Mark. Res. 1981, 24, 337–346. [Google Scholar] [CrossRef]
- Fonseca, M. Principles and Practice of Structural Equation Modeling, Third Edition by Rex B. Kline. Int. Stat. Rev. 2013, 81. [Google Scholar] [CrossRef]
- Chen, F. Sensitivity of Goodness of Fit Indexes to Lack of Measurement Invariance. Struct. Equ. Model. 2007, 14, 464–504. [Google Scholar] [CrossRef]
- Hu, L.T.; Bentler, P.M. Cutoff criteria for fit indexes in covariance structure analysis: Conventional criteria versus new alternatives. Struct. Equ. Model. 1998, 3, 55–61. [Google Scholar] [CrossRef]
- Gutsch, M.; Lasch, P.; Suckow, F.; Reyer, C. Evaluating the productivity of four main tree species in Germany under climate change with static reduced models. Ann. For. Sci. 2016, 73, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Xian, W.; Wang, X.; Tang, Z.; Shen, Z.; Zheng, C.; Xia, X.; Fang, J. The relationship between species richness and biomass changes from boreal to subtropical forests in China. Ecography 2014, 38. [Google Scholar] [CrossRef]
- Jiao-jun, Z.; Zhi-ping, F.; De-hui, Z.; Feng-qi, J.; Takeshi, M. Comparison of stand structure and growth between artificial and natural forests ofPinus sylvestiris var.mongolica on sandy land. J. For. Res. 2003, 14, 103–111. [Google Scholar] [CrossRef]
- Peng, H.; Tague, C.; Jia, Y. Evaluating the eco-hydrologic impacts of reforestation in the Loess Plateau, China, using an eco-hydrologic model. Ecohydrology 2015, 9. [Google Scholar] [CrossRef]
- Chen, W.; Jia, X.; Zha, T.-S.; Wu, B.; Zhang, Y.-Q.; Li, C.; Wang, X.; He, G.; Yu, H.; Guopeng, C. Soil respiration in a mixed urban forest in China in relation to soil temperature and water content. Eur. J. Soil Biol. 2013, 54, 63–68. [Google Scholar] [CrossRef]
- Sharma, C.M.; Baduni, N.; Gairola, S.; Ghildiyal, S.K.; Suyal, S. Effects of slope aspects on forest compositions, community structures and soil properties in natural temperate forests of Garhwal Himalaya. J. For. Res. 2010, 21, 331–337. [Google Scholar] [CrossRef]
- Måren, I.; Karki, S.; Prajapati, C.; Yadav, R.K.; Shrestha, B. Facing north or south: Does slope aspect impact forest stand characteristics and soil properties in a semiarid trans-Himalayan valley? J. Arid Environ. 2015, 121, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Stage, A.; Salas-Eljatib, C. Interactions of Elevation, Aspect, and Slope in Models of Forest Species Composition and Productivity. For. Sci. 2007, 53, 486–492. [Google Scholar]
- Cantlon, J. Vegetation and Microclimates on North and South Slopes of Cushetunk Mountain, New Jersey. Ecol. Monogr. 1953, 23, 241. [Google Scholar] [CrossRef]
- Socha, J. Effect of topography and geology on the site index of Picea abies in the West Carpathian, Poland. Scand. J. For. Res. 2008, 23, 203–213. [Google Scholar] [CrossRef]
- Hansson, K.; Laclau, J.-P.; Saint-Andre, L.; Mareschal, L.; Van der Heijden, G.; Nys, C.; Nicolas, M.; Ranger, J.; Legout, A. Chemical fertility of forest ecosystems. Part 1: Common soil chemical analyses were poor predictors of stand productivity across a wide range of acidic forest soils. For. Ecol. Manag. 2020, 461, 117843. [Google Scholar] [CrossRef]
- Braun, S.; Thomas, V.; Quiring, R.; Flückiger, W. Does nitrogen deposition increase forest production? The role of phosphorus. Environ. Pollut. 2009, 158, 2043–2052. [Google Scholar] [CrossRef]
- Belote, T.; Prisley, S.; Jones, R.; Fitzpatrick, M.; de Beurs, K. Forest productivity and tree diversity relationships depend on ecological context within mid-Atlantic and Appalachian forests (USA). For. Ecol. Manag. 2011, 261, 1315–1324. [Google Scholar] [CrossRef]
- Zeller, L.; Liang, J.; Pretzsch, H. Tree species richness enhances stand productivity while stand structure can have opposite effects, based on forest inventory data from Germany and the United States of America. For. Ecosyst. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Pommerening, A. Evaluating structural indices by reversing forest structural analysis. For. Ecol. Manag. 2006, 224, 266–277. [Google Scholar] [CrossRef]
- Newton, P.; Jolliffe, P. Aboveground modular component responses to intraspecific competition within density-stressed black spruce stands. Can. J. For. Res. 2011, 28, 1587–1610. [Google Scholar] [CrossRef]
- Zhou, W.; Cheng, X.; Wu, R.; Han, H.; Kang, F.; Zhu, J.; Tian, P. Effect of intraspecific competition on biomass partitioning of Larix principis-rupprechtii. J. Plant Interact. 2018, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yaussy, D.; Iverson, L.; Matthews, S. Competition and Climate Affects US Hardwood-Forest Tree Mortality. For. Sci. Wash. 2012, 59. [Google Scholar] [CrossRef] [Green Version]
- Hülsmann, L.; Bugmann, H.; Brang, P. How to predict tree death from inventory data—Lessons from a systematic assessment of European tree mortality models. Can. J. For. Res. 2017, 47. [Google Scholar] [CrossRef] [Green Version]
- Yücesan, Z.; Özçelik, S.; Oktan, E. Effects of thinning on stand structure and tree stability in an afforested oriental beech (Fagus orientalis Lipsky) stand in northeast Turkey. J. For. Res. 2015, 26. [Google Scholar] [CrossRef]
- Ishii, H.; Tanabe, S.-I.; Hiura, T. Exploring the Relationships among Canopy Structure, Stand Productivity, and Biodiversity of Temperate Forest Ecosystems. For. Sci. 2004, 50, 342–355. [Google Scholar]
- Zeller, L.; Pretzsch, H. Effect of forest structure on stand productivity in Central European forests depends on developmental stage and tree species diversity. For. Ecol. Manag. 2019, 434, 193–204. [Google Scholar] [CrossRef]
- Park, J.; Kim, H.S.; Jo, H.K.; Jung, I.I. The Influence of Tree Structural and Species Diversity on Temperate Forest Productivity and Stability in Korea. Forests 2019, 10, 1113. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Strata | Area (ha) | Wh | Ch | Ch2 | WhCh2 | nh = nWh |
|---|---|---|---|---|---|---|
| MAPM | 984.10 | 0.1063 | 0.3953 | 0.1563 | 0.0166 | 6 |
| MAPH | 5868.25 | 0.6338 | 0.3821 | 0.1460 | 0.0925 | 36 |
| MPH | 2344.84 | 0.2532 | 0.3882 | 0.1507 | 0.0382 | 15 |
| MPM | 62.29 | 0.0067 | 0.3036 | 0.0922 | 0.0006 | 0 |
| SUM | 9259.48 | 1 | - | - | 0.3053 | 57 |
| Variable | Min | Max | Mean | SD |
|---|---|---|---|---|
| Age (year) | 29 | 60 | 39 | 8.41 |
| Average DBH (cm) | 8.29 | 32.02 | 15.34 | 5.32 |
| Average height (m) | 8.05 | 18.87 | 13.12 | 2.27 |
| Elevation (m) | 1114 | 1519 | 1370.08 | 113.82 |
| Slope (°) | 3 | 40 | 18.67 | 8.55 |
| Litter thickness (cm) | 3.41 | 16.1 | 7.32 | 2.82 |
| Number of stems (ha) | 500 | 4533 | 2346 | 1131.19 |
| Canopy density | 0.6 | 0.98 | 0.87 | 0.06 |
| Basal area (m2/ha) | 15.64 | 58.26 | 34.69 | 8.49 |
| Latent Variable | Observed Variable | Abbreviation | Description | |
|---|---|---|---|---|
| Site Conditions | Litter thickness | LT | Mean thickness of litter in a stand | |
| Total carbon content | TC | Content of total carbon in soil | ||
| Total nitrogen content | TN | Content of total nitrogen in soil | ||
| Total phosphorus content | TP | Content of total phosphorus in soil | ||
| Soil water content | SWC | Content of water in soil | ||
| Soil organic matter | SOM | Content of organic matter in soil | ||
| Altitude | ALT | Altitude of Chinese pine population stand | ||
| Slope | SLO | Slope where Chinese pine population occurs | ||
| Forest Structure | Dominance | U | Proportion of the n nearest neighbors of a given reference tree that are smaller than the reference tree | [33,34] |
| Uniform angle index | W | Characterization of the spatial distribution of a forest community or of individual tree species within that community by gradually comparing the four included angles with the standard angle | [32] | |
| Crowding | C | Crowding degree of a neighborhood unit according to the overlapping of the crown in the spatial microenvironment, which clearly defines the crowding degree for a reference tree and its four nearest neighbors | [35] | |
| Forest Structure | Hegyi index | Hegyi | [36] | |
| SCI competition index | SCI | The distance-dependent competition index The structure-based competition index | [37] | |
| Mean diameter at breast height | MDBH | Average diameter of the Chinese pine population | ||
| Average height Crown density | AH CD | Average height of the Chinese pine population Aggregate of all vertically projected tree crowns onto the ground surface | ||
| Population stability | Density | D | Chinese pine population density | |
| Regeneration of undergrowth seedlings | RUS | The ratio of Chinese pine seedlings (DBH less than 3 cm) to the total number of plants in each plot | ||
| Forest mortality | FM | Proportion of dead wood in the forest plot | ||
| Average age | AGE | Average age of the Chinese pine population | ||
| Population productivity | Plot productivity | PP | Total productivity of trees in plot | |
| Average annual productivity | AAP | Average annual productivity of the plot | ||
| Individual productivity | IP | Average productivity of individual trees in the plot |
| Latent Variable Group | Canonical Correlations 1 | p-Value | Wilk’s Statistic | Chi-Square | DF |
|---|---|---|---|---|---|
| Site conditions and forest structure | 0.749 ** | 0.001 | 0.850 | 115.79 | 72 |
| Site conditions and population stability | 0.676 ** | 0.004 | 0.251 | 67.717 | 40 |
| Site conditions and population productivity | 0.633 | 0.058 | 0.470 | 37.722 | 24 |
| Forest structure and population stability | 0.875 *** | 0.000 | 0.111 | 106.691 | 45 |
| Forest structure and population productivity | 0.742 | 0.064 | 0.156 | 92.959 | 24 |
| Population stability and population productivity | 0.738 *** | 0.000 | 0.150 | 94.734 | 24 |
| Indicators | Cronbach’s Alpha | CR | AVE | SRME |
|---|---|---|---|---|
| Value | 0.893 | 0.839 | 0.885 | 0.099 |
| Reliability test standards | >0.6 | >0.7 | >0.5 | <0.1 |
| Latent Variable Group | Direct Effect | T-Value | Indirect Effect | T-Value | Total Effect | T-Value |
|---|---|---|---|---|---|---|
| Site conditions -> Forest structure | 0.449 *** | 3.801 | _ | _ | 0.449 *** | 3.801 |
| Site conditions -> Population stability | 0.307 * | 2.548 | 0.204 ** | 2.716 | 0.511 *** | 5.683 |
| Site conditions -> Population productivity | _ | _ | 0.249 *** | 3.921 | 0.249 *** | 3.921 |
| Forest structure -> Population stability | 0.454 *** | 4.046 | _ | _ | 0.454 *** | 4.046 |
| Forest structure -> Population productivity | _ | _ | 0.222 ** | 3.209 | 0.222 ** | 3.209 |
| Population stability -> Population productivity | 0.488 *** | 6.657 | _ | _ | 0.488 *** | 6.657 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Li, Y.; Song, H.; Jia, Y.; Liu, J. Agents Affecting the Productivity of Pine Plantations on the Loess Plateau in China: A Study Based on Structural Equation Modeling. Forests 2020, 11, 1328. https://doi.org/10.3390/f11121328
Zhao X, Li Y, Song H, Jia Y, Liu J. Agents Affecting the Productivity of Pine Plantations on the Loess Plateau in China: A Study Based on Structural Equation Modeling. Forests. 2020; 11(12):1328. https://doi.org/10.3390/f11121328
Chicago/Turabian StyleZhao, Xuan, Yanjie Li, Hao Song, Yuhuan Jia, and Jianjun Liu. 2020. "Agents Affecting the Productivity of Pine Plantations on the Loess Plateau in China: A Study Based on Structural Equation Modeling" Forests 11, no. 12: 1328. https://doi.org/10.3390/f11121328