The Chloroplast Genome of Carya illinoinensis: Genome Structure, Adaptive Evolution, and Phylogenetic Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Chloroplast Genome Sequencing
2.2. Chloroplast Genome Assembly and Annotation
2.3. Repeat Sequence Analysis and Codon Usage Analysis
2.4. SNP and Indel Detection
2.5. Chloroplast Genome Comparison
2.6. Molecular Evolution and Phylogenetic Analysis
3. Results and Discussion
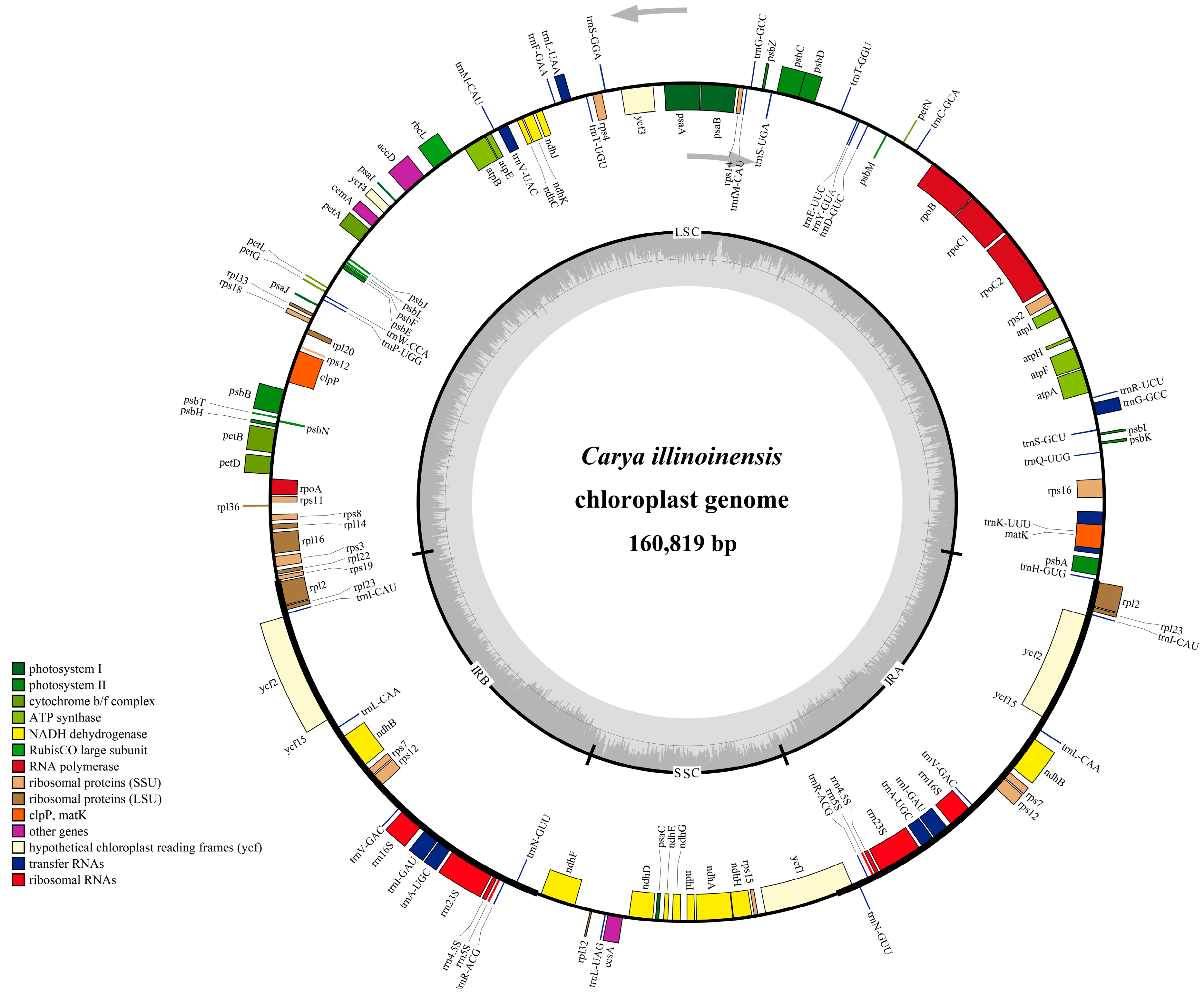
3.1. Chloroplast Genome Features of C. illinoinensis
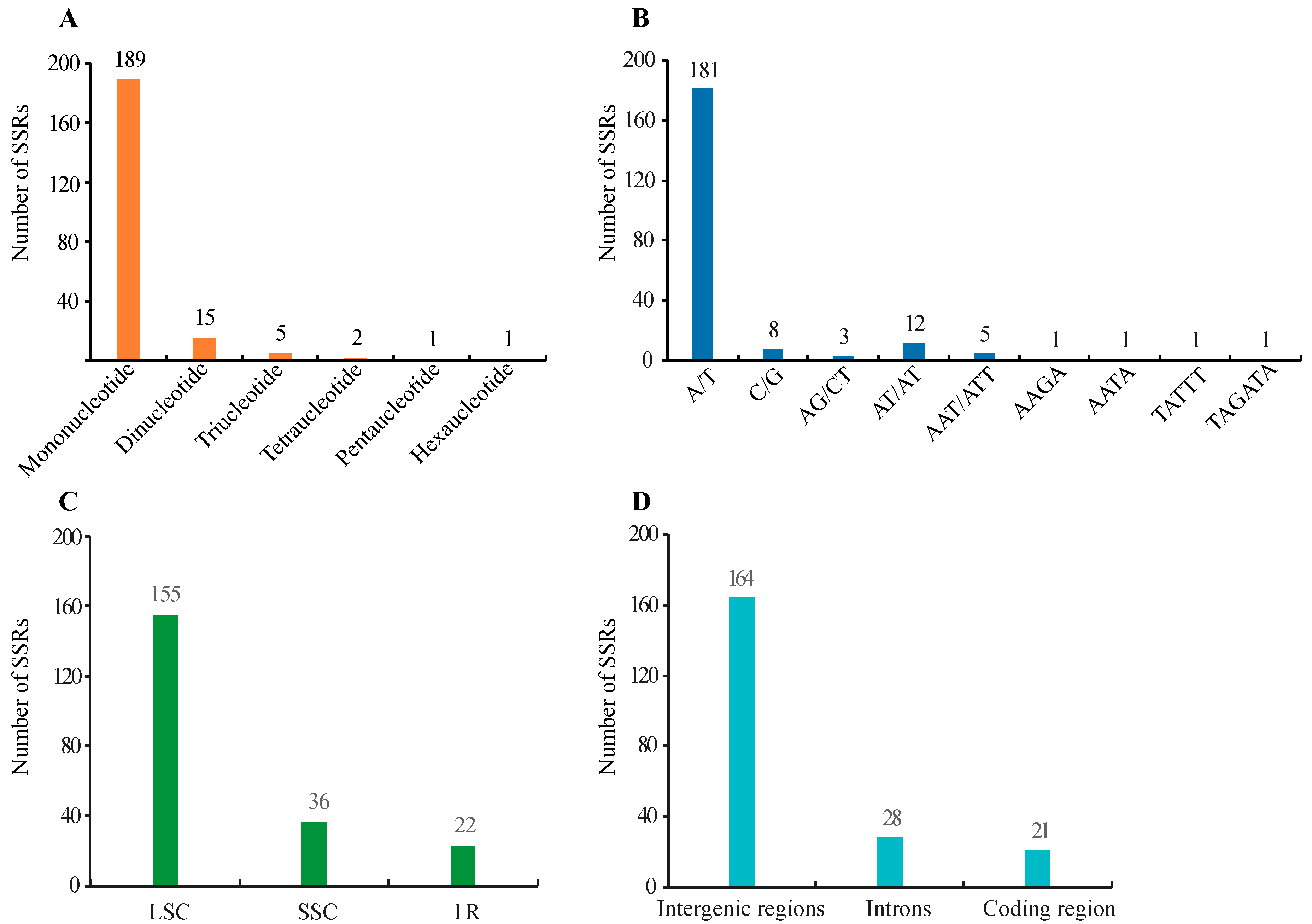
3.2. SSR and Long Repeats Identification
3.3. Codon Preference Analysis
3.4. SNP and Indel Detection
3.5. Comparative Chloroplast Genome Analysis
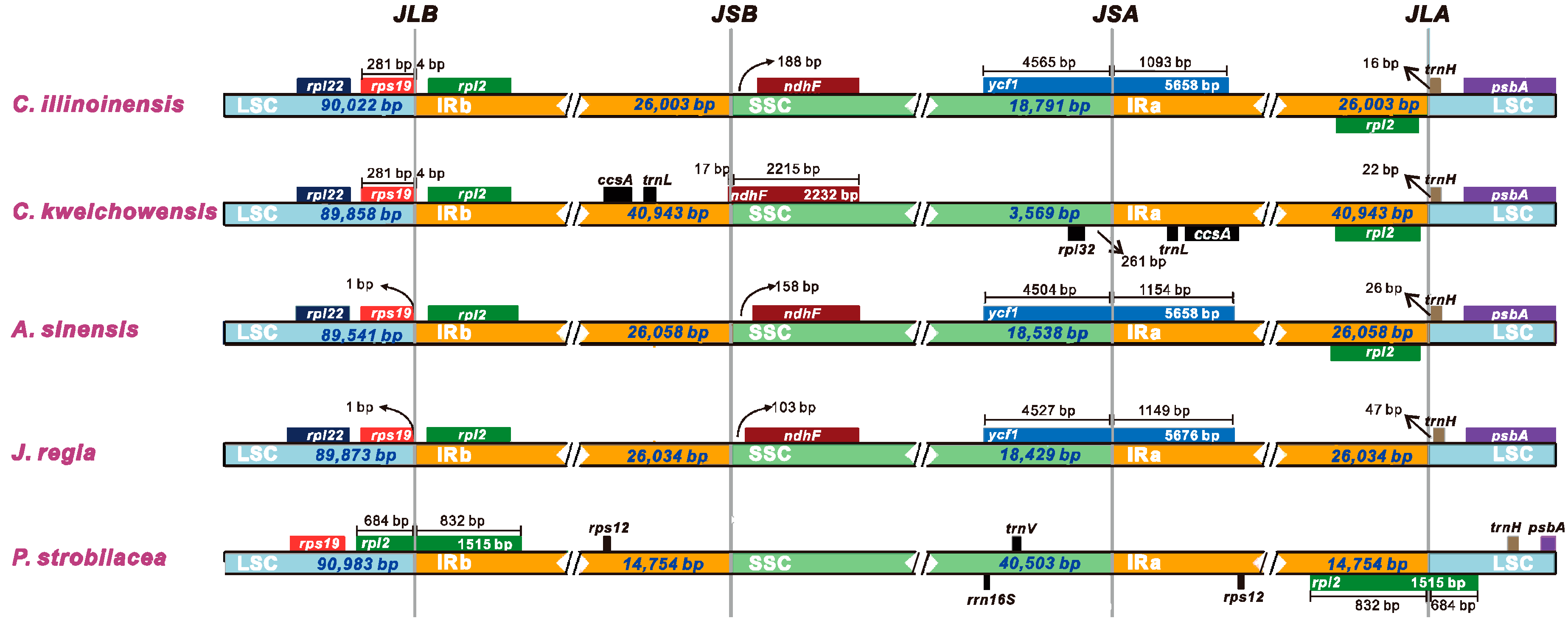
3.6. IR Contraction and Expansion
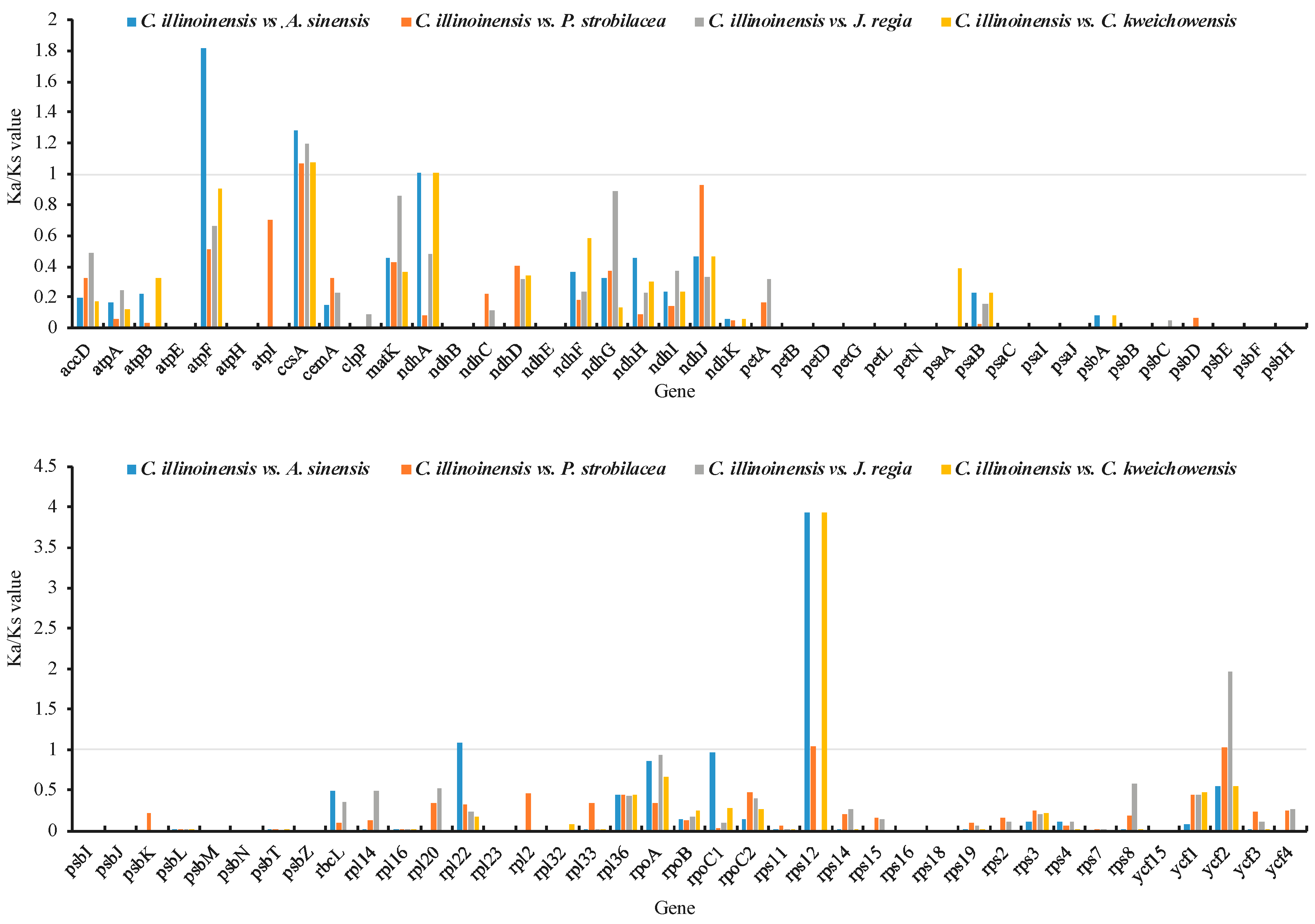
3.7. Selective Pressure in the Evolution of C. illinoinensis
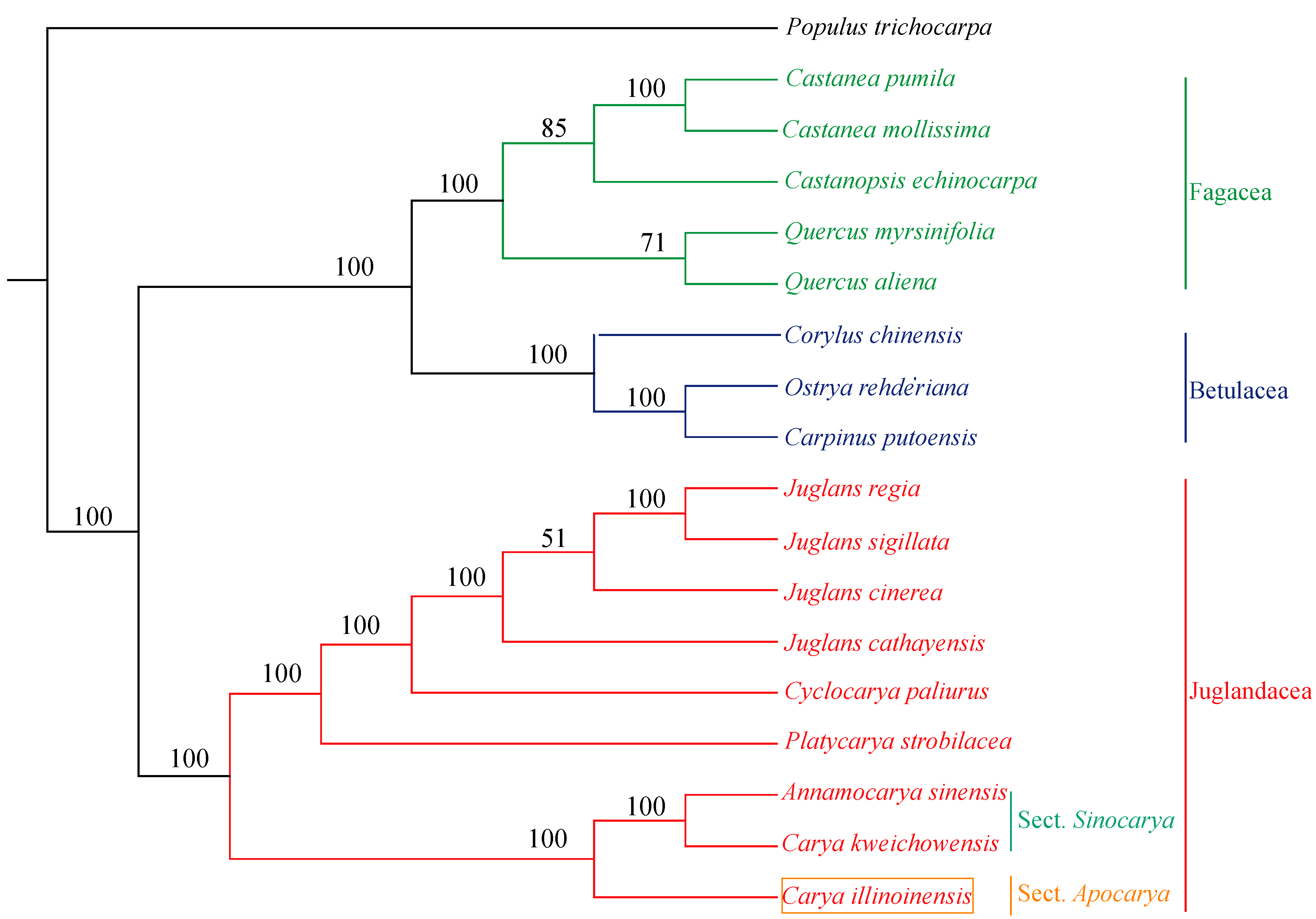
3.8. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drouin, G.; Daoud, H.; Xia, J. Relative rates of synonymous substitutions in the mitochondrial, chloroplast and nuclear genomes of seed plants. Mol. Phylogenetics Evol. 2008, 49, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.B.; Wu, Y.; Yang, J.; Shahzad, K.; Li, Z.H. Comparative chloroplast genomics of Dipsacales species: Insights into sequence variation, adaptive evolution, and phylogenetic relationships. Front. Plant Sci. 2018, 9, 689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Shi, C.; Liu, Y.; Mao, S.Y.; Gao, L.Z. Thirteen Camellia chloroplast genome sequences determined by high-throughput sequencing: Genome structure and phylogenetic relationships. BMC Evol. Biol. 2014, 14, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.C.; Lin, B.Y.; Lin, J.Y.; Wu, W.L.; Chang, C.C. Evaluation of chloroplast DNA markers for intraspecific identification of Phalaenopsis equestris cultivars. Sci. Hortic. 2016, 203, 86–94. [Google Scholar] [CrossRef]
- Mo, Z.; Feng, G.; Su, W.; Liu, Z.; Peng, F. Transcriptomic analysis provides insights into grafting union development in pecan (Carya illinoinensis). Genes 2018, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Lin, H.; Meng, C.; Jiang, P.; Fu, W. Effects of intercropping grasses on soil organic carbon and microbial community functional diversity under Chinese hickory (Carya cathayensis Sarg.) stands. Soil Res. 2014, 52, 575–583. [Google Scholar] [CrossRef]
- Mo, Z.; He, H.; Su, W.; Peng, F. Analysis of differentially accumulated proteins associated with graft union formation in pecan (Carya illinoensis). Sci. Hortic. 2017, 224, 126–134. [Google Scholar] [CrossRef]
- Mo, Z.; Feng, G.; Su, W.; Liu, Z.; Peng, F. Identification of miRNAs Associated with Graft Union Development in Pecan [Carya illinoinensis (Wangenh.) K. Koch]. Forests 2018, 9, 472. [Google Scholar] [CrossRef] [Green Version]
- Manos, P.S.; Stone, D.E. Evolution, phylogeny, and systematics of the Juglandaceae. Ann. Mo. Bot. Gard. 2001, 88, 231–269. [Google Scholar] [CrossRef]
- Lu, A.M.; Zhang, Z.Y. The Differentiation, Evolution and Systematic Relationship of Juglandales. J. Syst. Evol. 1990, 28, 96–102. [Google Scholar]
- Zhang, J.B.; Li, R.Q.; Xiang, X.G.; Manchester, S.R.; Lin, L.; Wang, W.; Wen, J.; Chen, Z.D. Integrated fossil and molecular data reveal the biogeographic diversification of the eastern Asian-eastern North American disjunct hickory genus (Carya Nutt.). PLoS ONE 2013, 8, e70449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcpherson, H.; Merwe, M.V.D.; Delaney, S.K.; Edwards, M.A.; Henry, R.J.; Mcintosh, E.; Rymer, P.D.; Milner, M.L.; Siow, J.; Rossetto, M. Capturing chloroplast variation for molecular ecology studies: A simple next generation sequencing approach applied to a rainforest tree. BMC Ecol. 2013, 13, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 1, 2047. [Google Scholar] [CrossRef]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [Green Version]
- Peter, S.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33, W686–W689. [Google Scholar]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Marçais, G.; Delcher, A.L.; Phillippy, A.M.; Coston, R.; Zimin, A. MUMmer4: A fast and versatile genome alignment system. PLoS Comput. Biol. 2018, 14, e1005944. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Frazer, K.A.; Lior, P.; Alexander, P.; Rubin, E.M.; Inna, D. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Woeste, K.E.; Dang, M.; Zhou, T.; Feng, X.; Zhao, G.; Liu, Z.; Li, Z.; Zhao, P. The complete chloroplast genome of common walnut (Juglans regia). Mitochondrial DNA 2016, 1, 189–190. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, X.; Feng, X.; Woeste, K.E.; Zhao, P. Characterization of the complete chloroplast genome of the endangered species Carya sinensis (Juglandaceae). Conserv. Genet. Resour. 2016, 8, 467–470. [Google Scholar] [CrossRef]
- Zhai, D.C.; Yao, Q.; Cao, X.F.; Hao, Q.Q.; Ma, M.T.; Pan, J.; Bai, X.H. Complete chloroplast genome of the wild-type Hickory Carya cathayensis. Mitochondrial DNA 2019, 4, 1457–1458. [Google Scholar] [CrossRef]
- Yan, C.; Du, J.; Gao, L.; Li, Y.; Hou, X. The complete chloroplast genome sequence of watercress (Nasturtium officinale R. Br.): Genome organization, adaptive evolution and phylogenetic relationships in Cardamineae. Gene 2019, 699, 24–36. [Google Scholar] [CrossRef]
- Redwan, R.; Saidin, A.; Kumar, S. Complete chloroplast genome sequence of MD-2 pineapple and its comparative analysis among nine other plants from the subclass Commelinidae. BMC Plant Biol. 2015, 15, 196. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.F.; Zhu, G.F.; Li, D.M.; Wang, X.J. Complete chloroplast genome sequence and phylogenetic analysis of Spathiphyllum’Parrish’. PLoS ONE 2019, 14, e0224038. [Google Scholar] [CrossRef]
- He, Y.; Xiao, H.; Deng, C.; Xiong, L.; Yang, J.; Peng, C. The complete chloroplast genome sequences of the medicinal plant Pogostemon cablin. Int. J. Mol. Sci. 2016, 17, 820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Woeste, K.E.; Zhao, P. Completion of the chloroplast genomes of five Chinese Juglans and their contribution to chloroplast phylogeny. Front. Plant Sci. 2017, 7, 1955. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.B.; Tang, M.; Li, H.T.; Zhang, Z.R.; Li, D.Z. Complete chloroplast genome of the genus Cymbidium: Lights into the species identification, phylogenetic implications and population genetic analyses. BMC Evol. Biol. 2013, 13, 84. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yu, H.; Wang, J.; Lei, W.; Gao, J.; Qiu, X.; Wang, J. The complete chloroplast genome sequences of the medicinal plant Forsythia suspensa (Oleaceae). Int. J. Mol. Sci. 2017, 18, 2288. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Zhang, X.; Liu, G.; Yin, Y.; Chen, K.; Yun, Q.; Zhao, D.; Al-Mssallem, I.S.; Yu, J. The complete chloroplast genome sequence of date palm (Phoenix dactylifera L.). PLoS ONE 2010, 5, e12762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, G.; Zhou, J.; Zhou, X.; Li, P.; Wang, Z. The first complete chloroplast genome sequence of Pteroceltis tatarinowii (Ulmaceae), an endangered tertiary relict tree endemic to China. Mitochondrial DNA 2019, 4, 487–488. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.H.; Lee, S.J.; Cho, B.; Lee, Y. Differential promoter usage of infA in response to cold shock in Escherichia coli. FEBS Lett. 2006, 580, 539–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millen, R.S.; Olmstead, R.G.; Adams, K.L.; Palmer, J.D.; Lao, N.T.; Heggie, L.; Kavanagh, T.A.; Hibberd, J.M.; Gray, J.C.; Morden, C.W. Many parallel losses of infA from chloroplast DNA during angiosperm evolution with multiple independent transfers to the nucleus. Plant Cell 2001, 13, 645–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaf, S.; Khan, A.L.; Khan, A.R.; Waqas, M.; Kang, S.; Khan, M.A.; Lee, S.; Lee, I. Complete Chloroplast Genome of Nicotiana otophora and its Comparison with Related Species. Front. Plant Sci. 2016, 7, 843. [Google Scholar] [CrossRef] [Green Version]
- Vera, A.; Sugiura, M. A novel RNA gene in the tobacco plastid genome: Its possible role in the maturation of 16S rRNA. EMBO J. 1994, 13, 2211–2217. [Google Scholar] [CrossRef]
- Sugita, M.; Svab, Z.; Maliga, P.; Sugiura, M. Targeted deletion of sprA from the tobacco plastid genome indicates that the encoded small RNA is not essential for pre-16S rRNA maturation in plastids. Mol. Gen. Genet. MGG 1997, 257, 23–27. [Google Scholar] [CrossRef]
- Ibrahim, R.I.H.; Azuma, J.-I.; Sakamoto, M. Complete nucleotide sequence of the cotton (Gossypium barbadense L.) chloroplast genome with a comparative analysis of sequences among 9 dicot plants. Genes Genet. Syst. 2006, 81, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Vu, H.T.; Tran, N.; Nguyen, T.D.; Vu, Q.L.; Bui, M.H.; Le, M.T.; Le, L. Complete chloroplast genome of Paphiopedilum delenatii and phylogenetic relationships among Orchidaceae. Plants 2020, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.L.; Wang, R.N.; Zhang, N.Y.; Fan, W.B.; Fang, M.F.; Li, Z.H. Molecular evolution of chloroplast genomes of orchid species: Insights into phylogenetic relationship and adaptive evolution. Int. J. Mol. Sci. 2018, 19, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, D.Y.; Wu, H.; Wang, Y.L.; Gao, L.M.; Zhang, S.Z.; Lu, L. Complete chloroplast genome sequence of Magnolia kwangsiensis (Magnoliaceae): Implication for DNA barcoding and population genetics. Genome 2011, 54, 663–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.M.; Zhao, C.Y.; Liu, X.F. Complete Chloroplast Genome Sequences of Kaempferia galanga and Kaempferia elegans: Molecular Structures and Comparative Analysis. Molecules 2019, 24, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef]
- Mengoni, A.; Barabesi, C.; Gonnelli, C.; Galardi, F.; Gabbrielli, R.; Bazzicalupo, M. Genetic diversity of heavy metal-tolerant populations in Silene paradoxa L.(Caryophyllaceae): A chloroplast microsatellite analysis. Mol. Ecol. 2001, 10, 1909–1916. [Google Scholar] [CrossRef]
- Provan, J.; Russell, J.; Booth, A.; Powell, W. Polymorphic chloroplast simple sequence repeat primers for systematic and population studies in the genus Hordeum. Mol. Ecol. 1999, 8, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Bausher, M.G.; Singh, N.D.; Lee, S.-B.; Jansen, R.K.; Daniell, H. The complete chloroplast genome sequence of Citrus sinensis (L.) Osbeck var’Ridge Pineapple’: Organization and phylogenetic relationships to other angiosperms. BMC Plant Biol. 2006, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.H.; Shang, A.Q.; Zhang, S.; Yu, X.Y.; Ren, Y.C.; Yang, M.S.; Wang, J.M. The first complete chloroplast genome sequences of Ulmus species by de novo sequencing: Genome comparative and taxonomic position analysis. PLoS ONE 2017, 12, e0171264. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Xue, Q. Comparative studies on codon usage pattern of chloroplasts and their host nuclear genes in four plant species. J. Genet. 2005, 84, 55–62. [Google Scholar] [CrossRef]
- Sharp, P.M.; Li, W.H. The codon adaptation index-a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res. 1987, 15, 1281–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasmahapatra, K.; Mallet, J. DNA barcodes: Recent successes and future prospects. Heredity 2006, 97, 254–255. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plant DNA barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, Z.; Sablok, G.; Daskalova, E.; Zahmanova, G.; Apostolova, E.; Yahubyan, G.; Baev, V. Chloroplast genome analysis of resurrection tertiary relict Haberlea rhodopensis highlights genes important for desiccation stress response. Front. Plant Sci. 2017, 8, 204. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zuo, L.; Lu, D.; Lu, B.; Yang, M.; Wang, J. Comparative analysis of chloroplast genomes of five Robinia species: Genome comparative and evolution analysis. Gene 2019, 689, 141–151. [Google Scholar] [CrossRef]
- Kim, K.J.; Lee, H.L. Complete chloroplast genome sequences from Korean ginseng (Panax schinseng Nees) and comparative analysis of sequence evolution among 17 vascular plants. DNA Res. 2004, 11, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Brunet, F.D.R.G.; Crollius, H.R.; Paris, M.; Aury, J.-M.; Gibert, P.; Jaillon, O.; Laudet, V.; Robinson-Rechavi, M. Gene loss and evolutionary rates following whole-genome duplication in teleost fishes. Mol. Biol. Evol. 2006, 23, 1808–1816. [Google Scholar] [CrossRef]
- Yang, Z.; Nielsen, R. Estimating synonymous and nonsynonymous substitution rates under realistic evolutionary models. Mol. Biol. Evol. 2000, 17, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Sablok, G.; Wang, B.; Qu, D.; Barbaro, E.; Viola, R.; Li, M.; Varotto, C. Plastome organization and evolution of chloroplast genes in Cardamine species adapted to contrasting habitats. BMC Genom. 2015, 16, 306. [Google Scholar] [CrossRef] [Green Version]
- Grauke, L.; Mendoza-Herrera, M.; Binzel, M. Plastid microsatellite markers in Carya. In Proceedings of the International Symposium on Molecular Markers in Horticulture 859, Mexico; 2009; pp. 237–246. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Features | C. illinoinensis |
|---|---|
| Genome size (bp)/GC content (%) | 160,819/36.15 |
| LSC size (bp)/GC content (%) | 90,022/33.74 |
| SSC size (bp)/GC content (%) | 18,791/29.89 |
| IR size (bp)/GC content (%) | 26,003/42.58 |
| Total gene number | 131 |
| Unique gene number | 112 |
| Protein-coding gene | 79 |
| tRNAs | 29 |
| rRNAs | 4 |
| Genes duplicated in IR | 19 |
| Groups of Genes | Name of Genes |
|---|---|
| Transfer RNAs | trnA-UGCab, trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCCab, trnH-GUG, trnI-CAUa, trnI-GAUab, trnK-UUUb, trnL-CAAa, trnL-UAG, trnL-UAAb, trnM-CAU, trnN-GUUa, trnP-UGG, trnQ-UUG, trnR-ACGa, trnR-UCU, trnS-GCU, trnS-UGA, trnS-GGA, trnT-GGU, trnT-UGU, trnV-GACa, trnV-UACb, trnW-CCA, trnY-GUA |
| ribosomal RNAs | rrn4.5Sa, rrn5Sa, rrn16Sa, rrn23Sa |
| Ribosomal protein small subunit | rps2, rps3, rps4, rps7a, rps8, rps11, rps12abc, rps14, rps15, rps16b, rps18, rps19 |
| Ribosomal protein large subunit | rpl2ab, rpl14, rpl16b, rpl20, rpl22, rpl23a, rpl32, rpl33, rpl36 |
| Subunits of RNA polymerase | rpoB, rpoA, rpoC1b, rpoC2 |
| Photosystem I | psaA, psaB, psaC, psaI, psaJ, ycf3b, ycf4 |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ |
| Cythochrome b/f complex | petA, petBb, petDb, petG, petL, petN |
| ATP synthase | atpA, atpB, atpE, atpFb, atpH, atpI |
| NADH-dehydrogenase | ndhAb, ndhBab, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK |
| Large subunit Rubisco | rbcL |
| Acetyl-CoA carboxylase | accD |
| Maturase | matK |
| Inner membrane protein | cemA |
| ATP-dependent protease | clpPb |
| Cytochrome c biogenesis | ccsA |
| Conserved open reading frames | ycf1, ycf2a, ycf15ad |
| Amino Acids | Codon | NO. | RSCU | Amino Acids | Codon | NO. | RSCU |
|---|---|---|---|---|---|---|---|
| Ala | GCA | 393 | 1.1293 | Pro | CCA | 303 | 1.1211 |
| Ala | GCC | 201 | 0.5775 | Pro | CCC | 209 | 0.7733 |
| Ala | GCG | 157 | 0.4511 | Pro | CCG | 154 | 0.5698 |
| Ala | GCT | 641 | 1.8419 | Pro | CCT | 415 | 1.5356 |
| Cys | TGC | 78 | 0.5182 | Gln | CAA | 734 | 1.5633 |
| Cys | TGT | 223 | 1.4817 | Gln | CAG | 205 | 0.4366 |
| Asp | GAC | 212 | 0.3962 | Arg | AGA | 488 | 1.8614 |
| Asp | GAT | 858 | 1.6037 | Arg | AGG | 167 | 0.6369 |
| Glu | GAA | 1020 | 1.4988 | Arg | CGA | 360 | 1.3731 |
| Glu | GAG | 341 | 0.5011 | Arg | CGC | 103 | 0.3928 |
| Pro | TTC | 517 | 0.6991 | Arg | CGG | 115 | 0.4386 |
| Pro | TTT | 962 | 1.3008 | Arg | CGT | 340 | 1.2968 |
| Gly | GGA | 725 | 1.6467 | Ser | AGC | 127 | 0.381 |
| Gly | GGC | 180 | 0.4088 | Ser | AGT | 403 | 1.209 |
| Gly | GGG | 270 | 0.6132 | Ser | TCA | 407 | 1.221 |
| Gly | GGT | 586 | 1.331 | Ser | TCC | 321 | 0.963 |
| His | CAC | 131 | 0.4192 | Ser | TCG | 182 | 0.546 |
| His | CAT | 494 | 1.5808 | Ser | TCT | 560 | 1.68 |
| Ile | ATA | 759 | 0.9734 | STOP | TAA | 43 | 1.4999 |
| Ile | ATC | 443 | 0.5681 | STOP | TAG | 23 | 0.8023 |
| Ile | ATT | 1137 | 1.4583 | STOP | TGA | 20 | 0.6976 |
| Lys | AAA | 1047 | 1.483 | Thr | ACA | 391 | 1.2095 |
| Lys | AAG | 365 | 0.5169 | Thr | ACC | 230 | 0.7115 |
| Leu | CTA | 374 | 0.8054 | Thr | ACG | 143 | 0.4423 |
| Leu | CTC | 187 | 0.4027 | Thr | ACT | 529 | 1.6365 |
| Leu | CTG | 177 | 0.3811 | Val | GTA | 546 | 1.5611 |
| Leu | CTT | 602 | 1.2964 | Val | GTC | 169 | 0.4832 |
| Leu | TTA | 887 | 1.9102 | Val | GTG | 180 | 0.5146 |
| Leu | TTG | 559 | 1.2038 | Val | GTT | 504 | 1.441 |
| Met | ATG | 607 | 1 | Trp | TGG | 463 | 1 |
| Asn | AAC | 297 | 0.4608 | Tyr | TAC | 202 | 0.4064 |
| Asn | AAT | 992 | 1.5391 | Tyr | TAT | 792 | 1.5935 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, Z.; Lou, W.; Chen, Y.; Jia, X.; Zhai, M.; Guo, Z.; Xuan, J. The Chloroplast Genome of Carya illinoinensis: Genome Structure, Adaptive Evolution, and Phylogenetic Analysis. Forests 2020, 11, 207. https://doi.org/10.3390/f11020207
Mo Z, Lou W, Chen Y, Jia X, Zhai M, Guo Z, Xuan J. The Chloroplast Genome of Carya illinoinensis: Genome Structure, Adaptive Evolution, and Phylogenetic Analysis. Forests. 2020; 11(2):207. https://doi.org/10.3390/f11020207
Chicago/Turabian StyleMo, Zhenghai, Wenrui Lou, Yaqi Chen, Xiaodong Jia, Min Zhai, Zhongren Guo, and Jiping Xuan. 2020. "The Chloroplast Genome of Carya illinoinensis: Genome Structure, Adaptive Evolution, and Phylogenetic Analysis" Forests 11, no. 2: 207. https://doi.org/10.3390/f11020207