Net Primary Productivity of Pinus massoniana Dependence on Climate, Soil and Forest Characteristics


Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection and Treatments
2.2. Influencing Factors
2.2.1. Soil Data and Stand Characteristics
2.2.2. Climatic Variables
2.3. Statistical Analysis
3. Results
3.1. Variability of P. mansoniana Distribution and NPP
3.2. Factors Influencing NPP of Different P. massoniana Components
3.3. Relationship between Site Conditions and Environmental Factors
3.4. Relationship between Temperatures and P. massoniana NPP
4. Discussion
4.1. Factors Influencing NPP of P. massoniana Forests
4.1.1. Climate Effects
4.1.2. Soil Effects
4.1.3. Stand Characteristics Effects
4.2. Latitudinal Effects on NPP of P. massoniana Forests
4.3. Uncertainty Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef]
- Hamilton, S.E.; Friess, D.A. Global carbon stocks and potential emissions due to mangrove deforestation from 2000 to 2012. Nat. Clim. Chang. 2018, 8, 240–244. [Google Scholar] [CrossRef]
- Keeling, H.C.; Phillips, O.L. The global relationship between forest productivity and biomass. Glob. Ecol. Biogeogr. 2007, 16, 618–631. [Google Scholar] [CrossRef]
- Malhi, Y.; Doughty, C.; Galbraith, D. The allocation of ecosystem net primary productivity in tropical forests. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3225–3245. [Google Scholar] [CrossRef]
- Cloern, J.E.; Abreu, P.C.; Carstensen, J.; Chauvaud, L.; Elmgren, R.; Grall, J.; Greening, H.; Johansson, J.O.R.; Kahru, M.; Sherwood, E.T.; et al. Human activities and climate variability drive fast-paced change across the world’s estuarine-coastal ecosystems. Glob. Chang. Biol. 2016, 22, 513–529. [Google Scholar] [CrossRef]
- Drake, J.E.; Gallet-Budynek, A.; Hofmockel, K.S.; Bernhardt, E.S.; Billings, S.A.; Jackson, R.B.; Johnsen, K.S.; Lichter, J.; Mccarthy, H.R.; Mccormack, M.L.; et al. Increases in the flux of carbon belowground stimulate nitrogen uptake and sustain the long-term enhancement of forest productivity under elevated CO2. Ecol. Lett. 2011, 14, 349–357. [Google Scholar] [CrossRef]
- Malhi, Y.; Girardin, C.A.J.; Goldsmith, G.R.; Doughty, C.E.; Salinas, N.; Metcalfe, D.B.; Huaraca Huasco, W.; Silva-Espejo, J.E.; del Aguilla-Pasquell, J.; Farfán Amézquita, F.; et al. The variation of productivity and its allocation along a tropical elevation gradient: A whole carbon budget perspective. New Phytol. 2017, 214, 1019–1032. [Google Scholar] [CrossRef]
- Girardin, C.A.J.; Malhi, Y.; Aragão, L.E.O.C.; Mamani, M.; Huaraca Huasco, W.; Durand, L.; Feeley, K.J.; Rapp, J.; Silva-Espejo, J.E.; Silman, M.; et al. Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the Peruvian Andes. Glob. Chang. Biol. 2010, 16, 3176–3192. [Google Scholar] [CrossRef]
- Malhi, Y. The productivity, metabolism and carbon cycle of tropical forest vegetation. J. Ecol. 2012, 100, 65–75. [Google Scholar] [CrossRef]
- Ni, J.; Zhang, X.S.; Scurlock, J.M.O. Synthesis and analysis of biomass and net primary productivity in Chinese forests. Ann. For. Sci. 2001, 58, 351–384. [Google Scholar] [CrossRef]
- Ni, J. Net primary productivity in forests of China: Scaling-up of national inventory data and comparison with model predictions. For. Ecol. Manag. 2003, 176, 485–495. [Google Scholar] [CrossRef]
- Yang, J.; Ji, X.; Deane, D.C.; Wu, L.; Chen, S. Spatiotemporal distribution and driving factors of forest biomass carbon storage in China: 1977–2013. Forests 2017, 8, 263. [Google Scholar] [CrossRef]
- Xu, X.; Li, K. Biomass carbon sequestration by planted forests in China. Chin. Geogr. Sci. 2010, 20, 289–297. [Google Scholar] [CrossRef]
- Silva, L.N.; Freer-Smith, P.; Madsen, P. Production, restoration, mitigation: A new generation of plantations. New For. 2019, 50, 153–168. [Google Scholar] [CrossRef]
- Zhao, M.; Zhou, G.S. Estimation of biomass and net primary productivity of major planted forests in China based on forest inventory data. For. Ecol. Manag. 2005, 207, 295–313. [Google Scholar] [CrossRef]
- Payn, T.; Carnus, J.M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. Changes in planted forests and future global implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef]
- Xi, W.; Wang, F.; Shi, P.; Dai, E.; Anoruo, A.O.; Bi, H.; Rahmlow, A.; He, B.; Li, W. Challenges to sustainable development in China: A review of six large-scale forest restoration and land conservation programs. J. Sustain. For. 2014, 33, 435–453. [Google Scholar] [CrossRef]
- Zhou, Z.X. Masson Pine in China; China Forestry Publishing House: Beijing, China, 2001. (In Chinese) [Google Scholar]
- Feng, Z.W.; Chen, C.Y.; Zhang, J.W.; Wang, K.P.; Zhao, J.L.; Gao, H. Determination of biomass of Pinus massoniana stand in Huitong county, Hunan province. Sci. Silvae Sin. 1982, 18, 127–134. (In Chinese) [Google Scholar]
- Ali, A.; Ahmad, A.; Akhtar, K.; Teng, M.; Zeng, W.; Yan, Z.; Zhou, Z. Patterns of biomass, carbon, and soil properties in masson pine (Pinus massoniana Lamb) plantations with different stand ages and management practices. Forests 2019, 10, 645. [Google Scholar] [CrossRef]
- Xiang, W.; Liu, S.; Deng, X.; Shen, A.; Lei, X.; Tian, D.; Zhao, M.; Peng, C. General allometric equations and biomass allocation of Pinus massoniana trees on a regional scale in southern China. Ecol. Res. 2011, 26, 697–711. [Google Scholar] [CrossRef]
- You, Y.; Huang, X.; Zhu, H.; Liu, S.; Liang, H.; Wen, Y.; Wang, H.; Cai, D.; Ye, D. Positive interactions between Pinus massoniana and Castanopsis hystrix species in the uneven-aged mixed plantations can produce more ecosystem carbon in subtropical China. For. Ecol. Manag. 2018, 410, 193–200. [Google Scholar] [CrossRef]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, biomass, and productivity of mangrove forests: A review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Liang, H.; Huang, J.G.; Ma, Q.; Li, J.; Wang, Z.; Guo, X.; Zhu, H.; Jiang, S.; Zhou, P.; Yu, B.; et al. Contributions of competition and climate on radial growth of Pinus massoniana in subtropics of China. Agric. For. Meteorol. 2019, 274, 7–17. [Google Scholar] [CrossRef]
- Zhang, H.; Song, T.; Wang, K.; Wang, G.; Liao, J.; Xu, G.; Zeng, F. Biogeographical patterns of forest biomass allocation vary by climate, soil and forest characteristics in China. Environ. Res. Lett. 2015, 10, 44014. [Google Scholar] [CrossRef]
- Liu, L.B.; Yang, H.M.; Xu, Y.; Guo, Y.M.; Ni, J. Forest biomass and net primary productivity in Southwestern China: A meta-analysis focusing on environmental driving factors. Forests 2016, 7, 173. [Google Scholar] [CrossRef]
- Zhan, X.; Guo, M.; Zhang, T. Joint control of net primary productivity by climate and soil nitrogen in the forests of Eastern China. Forests 2018, 9, 173. [Google Scholar]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Chen, G.; Hobbie, S.E.; Reich, P.B.; Yang, Y.; Robinson, D. Allometry of fine roots in forest ecosystems. Ecol. Lett. 2019, 22, 322–331. [Google Scholar] [CrossRef]
- Zhang, W.J.; Wang, H.M.; Yang, F.T.; Yi, Y.H.; Wen, X.F.; Sun, X.M.; Yu, G.R.; Wang, Y.D.; Ning, J.C. Underestimated effects of low temperature during early growing season on carbon sequestration of a subtropical coniferous plantation. Biogeosciences 2011, 8, 1667–1678. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, Q.; Xu, M.; Chi, Y.; Shen, R.; Li, P.; Dai, H. Responses of Pinus massoniana and Pinus taeda to freezing in temperate forests in central China. Scand. J. For. Res. 2012, 27, 520–531. [Google Scholar] [CrossRef]
- Grueber, C.E.; Nakagawa, S.; Laws, R.J.; Jamieson, I.G. Multimodel inference in ecology and evolution: Challenges and solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bartoń, K. MuMIn: Multi-Model Inference, R package version 1.43.15. 2019. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 20 December 2019).
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.J.; Yu, S.L.; Zhang, T.W. Influence of climate warming and resin collection on the growth of Masson pine (Pinus massoniana) in a subtropical forest, southern China. Trees-Struct. Funct. 2015, 29, 1423–1430. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Zhou, G.; Huang, W.; Duan, H. Warming effects on photosynthesis of subtropical tree species: A translocation experiment along an altitudinal gradient. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, G.; Liu, J. Different growth and physiological responses of six subtropical tree species to warming. Front. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Zhou, H.; Luo, Y.; Zhou, G.; Yu, J.; Shah, S.; Meng, S.; Liu, Q. Exploring the sensitivity of subtropical stand aboveground productivity to local and regional climate signals in South China. Forests 2019, 10, 71. [Google Scholar] [CrossRef]
- Lindroth, A.; Grelle, A.; More, A. Long-term measurements of boreal forest carbon balance. Glob. Chang. Biol. 1998, 4, 443–450. [Google Scholar] [CrossRef]
- Schuur, E.A.G. Productivity and global climate revisited: The sensitivity of tropical forest growth to precipitation. Ecology 2003, 84, 1165–1170. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, C.Y.; Zhang, Z.; Chen, Y.D.; Liu, C.L. Spatial and temporal variability of precipitation over China, 1951–2005. Theor. Appl. Climatol. 2009, 95, 53–68. [Google Scholar] [CrossRef]
- Liang, W.; Yang, Y.; Fan, D.; Guan, H.; Zhang, T.; Long, D.; Zhou, Y.; Bai, D. Analysis of spatial and temporal patterns of net primary production and their climate controls in China from 1982 to 2010. Agric. For. Meteorol. 2015, 204, 22–36. [Google Scholar] [CrossRef]
- Wang, F.; Xu, Y.J.; Dean, T.J. Projecting climate change effects on forest net primary productivity in subtropical Louisiana, USA. Ambio 2011, 40, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Churkina, G.; Running, S.W. Contrasting climatic controls on the estimated productivity of global terrestrial biomes. Ecosystems 1998, 1, 206–215. [Google Scholar] [CrossRef]
- Huang, W.; Liu, J.; Wang, Y.P.; Zhou, G.; Han, T.; Li, Y. Increasing phosphorus limitation along three successional forests in southern China. Plant Soil 2013, 364, 181–191. [Google Scholar] [CrossRef]
- Hu, C.; Zhao, L.; Zhou, Z.; Dong, G.; Zhang, Y. Genetic variations and correlation analysis of N and P traits in Pinus massoniana under combined conditions of N deposition and P deficiency. Trees-Struct. Funct. 2016, 30, 1341–1350. [Google Scholar] [CrossRef]
- Fang, Y.; Zou, X.; Lie, Z.; Xue, L. Variation in organ biomass with changing climate and forest characteristics across Chinese forests. Forests 2018, 9, 521. [Google Scholar] [CrossRef]
- Ryan, M.G.; Binkley, D.; Fownes, J.H. Age-related decline in forest productivity: Pattern and process. Adv. Ecol. Res. 1997, 27, 213–262. [Google Scholar]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Bormann, B.T.; Gordon, J.C. Stand density effects in young red alder plantations: Productivity, photosynthate partitioning, and nitrogen fixation. Ecology 1984, 65, 394–402. [Google Scholar] [CrossRef]
- Drew, T.J.; Flewelling, J.W. Stand density management: An alternative approach and its application to Douglas-fir plantations. For. Sci. 1979, 25, 518–532. [Google Scholar]
- Horner, G.J.; Baker, P.J.; Mac Nally, R.; Cunningham, S.C.; Thomson, J.R.; Hamilton, F. Forest structure, habitat and carbon benefits from thinning floodplain forests: Managing early stand density makes a difference. For. Ecol. Manag. 2010, 259, 286–293. [Google Scholar] [CrossRef]
- Xue, L.; Hagihara, A. Growth analysis on the C-D effect in self-thinning Masson pine (Pinus massoniana) stands. For. Ecol. Manag. 2002, 165, 249–256. [Google Scholar] [CrossRef]
- Magruder, M.; Chhin, S.; Monks, A.; O’Brien, J. Effects of initial stand density and climate on red pine productivity within Huron National Forest, Michigan, USA. Forests 2012, 3, 1086–1103. [Google Scholar] [CrossRef]
- López-Medellín, X.; Ezcurra, E. The productivity of mangroves in northwestern Mexico: A meta-analysis of current data. J. Coast. Conserv. 2012, 16, 399–403. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, K.; Xu, X.; Song, T.; Xu, Y.; Zeng, F. Biogeographical patterns of biomass allocation in leaves, stems, and roots in Chinas forests. Sci. Rep. 2015, 5, 1–12. [Google Scholar]
- Han, W.X.; Fang, J.Y.; Reich, P.B.; Ian Woodward, F.; Wang, Z.H. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. Lett. 2011, 14, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Zhao, A.; Li, Q.; Kong, X.; Ji, G. Acidity regime of the red soils in a subtropical region of southern China under field conditions. Geoderma 2003, 115, 75–84. [Google Scholar] [CrossRef]
- Sun, F.; Kuang, Y.; Wen, D.; Xu, Z.; Li, J.; Zuo, W.; Hou, E. Long-term tree growth rate, water use efficiency, and tree ring nitrogen isotope composition of Pinus massoniana L. in response to global climate change and local nitrogen deposition in Southern China. J. Soils Sediments 2010, 10, 1453–1465. [Google Scholar] [CrossRef]
- Wu, T.; Lin, W.; Li, Y.; Lie, Z.; Huang, W.; Liu, J. Nitrogen addition method affects growth and nitrogen accumulation in seedlings of four subtropical tree species: Schima superba Gardner & Champ., Pinus massoniana Lamb., Acacia mangium Willd., and Ormosia pinnata Lour. Ann. For. Sci. 2019, 76, 1–11. [Google Scholar]
- Zhang, Y.; Zhou, Z.; Yang, Q. Nitrogen (N) Deposition impacts seedling growth of Pinus massoniana via N:P ratio effects and the modulation of adaptive responses to low P (phosphorus). PLoS ONE 2013, 8, e79229. [Google Scholar] [CrossRef]
- Huang, W.; Zhou, G.; Liu, J.; Duan, H.; Liu, X.; Fang, X.; Zhang, D. Shifts in soil phosphorus fractions under elevated CO2 and N addition in model forest ecosystems in subtropical China. Plant Ecol. 2014, 215, 1373–1384. [Google Scholar] [CrossRef]
- Pan, P.; Zhao, F.; Ning, J.; Zhang, L.; Ouyang, X.; Zang, H. Impact of understory vegetation on soil carbon and nitrogen dynamic in aerially seeded Pinus massoniana plantations. PLoS ONE 2018, 13, e0191952. [Google Scholar] [CrossRef] [PubMed]






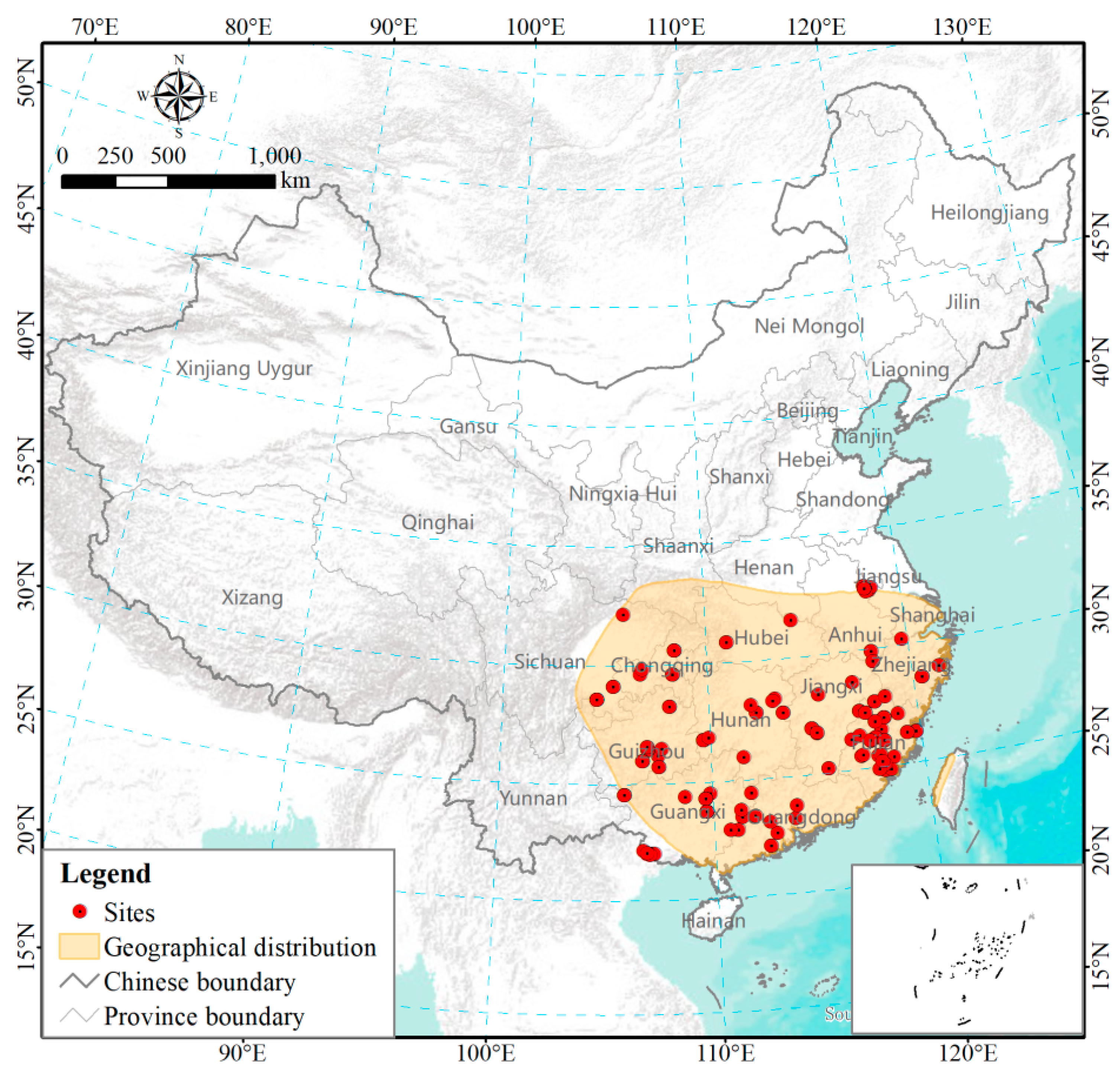{kind=link}
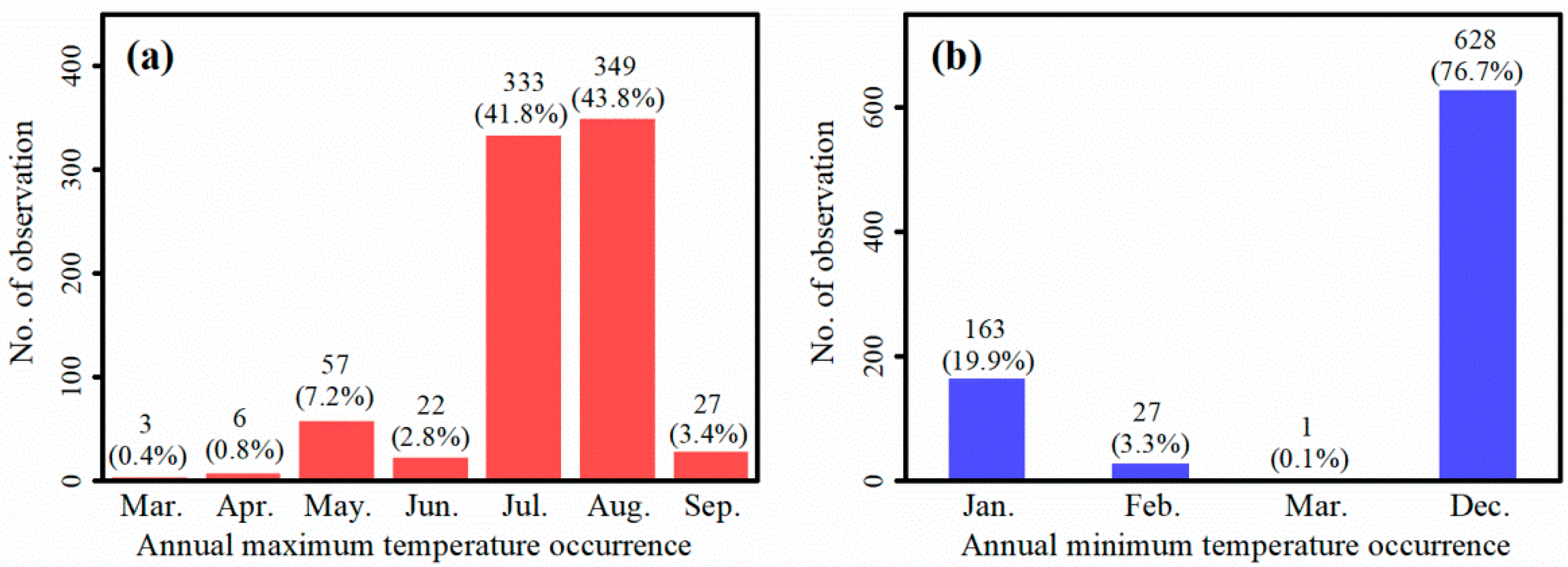{kind=link}
{kind=link}
{kind=link}
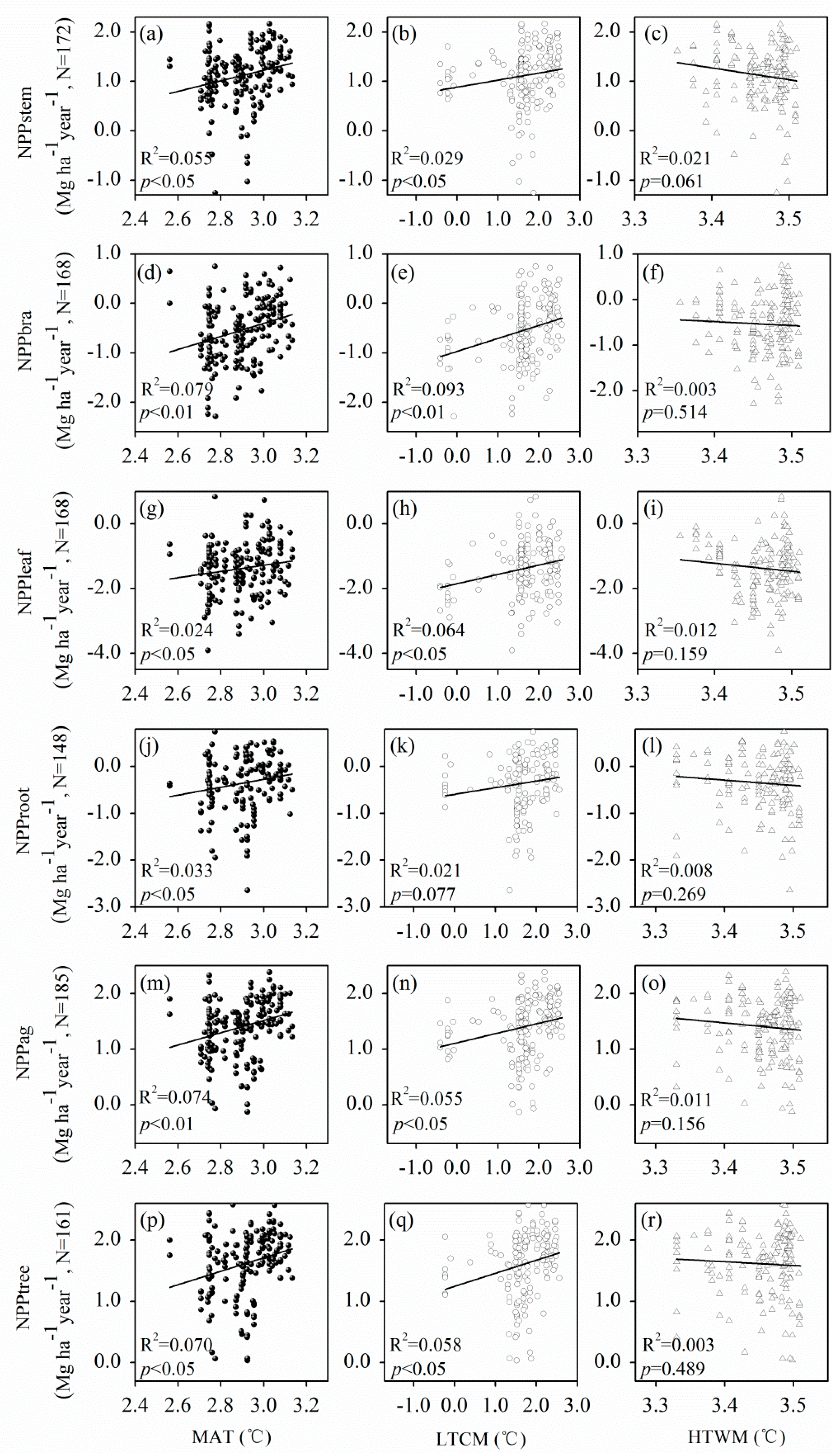{kind=link}
| Component | N | NPP (Mg·ha−1·year−1) | |||
|---|---|---|---|---|---|
| Mean | Max | Min | SE | ||
| Stem | 172 | 3.51 | 8.71 | 0.29 | 0.13 |
| Branch | 168 | 0.69 | 2.11 | 0.10 | 0.03 |
| Leaf | 168 | 0.34 | 2.31 | 0.02 | 0.02 |
| Root | 148 | 0.81 | 2.11 | 0.07 | 0.03 |
| Aboveground | 185 | 4.53 | 10.81 | 0.88 | 0.15 |
| Total tree | 161 | 5.65 | 13.13 | 1.04 | 0.20 |
| Site Conditions | NPPstem | NPPbra | NPPleaf | NPProot | NPPag | NPPtree |
|---|---|---|---|---|---|---|
| Longitude (°E) | −0.112 | −0.054 | −0.007 | −0.110 | −0.100 | −0.119 |
| Latitude (°N) | −0.285 ** | −0.369 ** | −0.208 ** | −0.251 ** | −0.338 ** | −0.344 ** |
| Elevation (m) | −0.087 | −0.019 | 0.102 | −0.069 | −0.061 | −0.094 |
| NPP Component | Parameter | Variable | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Intercept | MAT | HTWM | AGE | DENSITY | BD | MAP | AN | AP | SOM | ||
| Stem | Estimate | 8.797 | 1.575 | −5.099 | 0.562 | 0.216 | 1.170 | — | — | 0.424 | 0.370 |
| SE | 5.746 | 0.466 | 1.745 | 0.086 | 0.067 | 0.585 | — | — | 0.128 | 0.171 | |
| t-value | 1.531 | 3.378 | −2.922 | 6.520 | 3.233 | 1.999 | — | — | 3.311 | 2.160 | |
| p-value | 0.131 | <0.01 | <0.01 | <0.001 | <0.01 | <0.05 | — | — | <0.01 | <0.05 | |
| Branch | Estimate | 2.524 | — | −3.997 | — | 0.148 | 0.913 | 1.294 | — | — | — |
| SE | 7.119 | — | 2.472 | — | 0.059 | 0.654 | 0.455 | — | — | — | |
| t-value | 0.355 | — | −1.617 | — | 2.533 | 1.397 | 2.844 | — | — | — | |
| p-value | 0.724 | — | 0.111 | — | <0.05 | 0.168 | <0.01 | — | — | — | |
| Leaf | Estimate | 6.673 | — | −4.488 | −0.588 | 0.144 | 0.661 | 1.098 | — | — | — |
| SE | 7.924 | — | 2.777 | 0.097 | 0.073 | 0.725 | 0.515 | — | — | — | |
| t-value | 0.842 | — | −1.616 | −6.045 | 1.964 | 0.912 | 2.133 | — | — | — | |
| p-value | 0.403 | — | 0.111 | <0.001 | 0.052 | 0.366 | <0.05 | — | — | — | |
| Root | Estimate | −1.479 | 1.173 | −2.815 | 0.429 | 0.257 | 1.885 | — | 0.833 | — | — |
| SE | 5.436 | 0.656 | 1.589 | 0.091 | 0.070 | 0.898 | — | 0.439 | — | — | |
| t-value | −0.272 | 1.787 | −1.772 | 4.699 | 3.680 | 2.098 | — | 1.894 | — | — | |
| p-value | 0.786 | 0.079 | 0.081 | <0.001 | <0.001 | <0.05 | — | 0.063 | — | — | |
| Aboveground | Estimate | 1.494 | 1.142 | −1.666 | 0.333 | 0.182 | — | — | — | — | — |
| SE | 3.510 | 0.412 | 0.998 | 0.077 | 0.064 | — | — | — | — | — | |
| t-value | 0.426 | 2.771 | −1.669 | 4.333 | 2.852 | — | — | — | — | — | |
| p-value | 0.671 | <0.01 | 0.099 | <0.001 | <0.01 | — | — | — | — | — | |
| Total tree | Estimate | 1.920 | 0.989 | −4.158 | 0.409 | 0.258 | 1.356 | 0.681 | 0.598 | — | — |
| SE | 4.635 | 0.520 | 1.910 | 0.080 | 0.066 | 0.647 | 0.529 | 0.341 | — | — | |
| t-value | 0.414 | 1.900 | −2.177 | 5.092 | 3.911 | 2.096 | 1.287 | 1.757 | — | — | |
| p-value | 0.680 | 0.062 | <0.05 | <0.001 | <0.001 | <0.05 | 0.202 | 0.083 | — | — | |
| Variables | MAT | LTCM | HTWM | MAP |
|---|---|---|---|---|
| Longitude (°E) | −0.073 | −0.173 * | 0.352 ** | 0.242 ** |
| Latitude (°N) | −0.291 ** | −0.413 ** | −0.061 | −0.256 ** |
| Elevation (m) | −0.334 ** | 0.319 ** | −0.391 ** | −0.048 |
| Variables | AP | AN | AK | PH | SOM | BD |
|---|---|---|---|---|---|---|
| Longitude (°E) | −0.068 | −0.175 * | −0.291 ** | 0.051 | −0.341 ** | 0.152 * |
| Latitude (°N) | 0.223 ** | −0.093 | −0.074 | 0.216 ** | −0.216 ** | −0.112 |
| Elevation (m) | 0.016 | 0.417 ** | 0.292 ** | 0.153 * | 0.280 ** | −0.213 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Huang, C.; Teng, M.; Zhou, Z.; Wang, P. Net Primary Productivity of Pinus massoniana Dependence on Climate, Soil and Forest Characteristics. Forests 2020, 11, 404. https://doi.org/10.3390/f11040404
Huang X, Huang C, Teng M, Zhou Z, Wang P. Net Primary Productivity of Pinus massoniana Dependence on Climate, Soil and Forest Characteristics. Forests. 2020; 11(4):404. https://doi.org/10.3390/f11040404
Chicago/Turabian StyleHuang, Xin, Chunbo Huang, Mingjun Teng, Zhixiang Zhou, and Pengcheng Wang. 2020. "Net Primary Productivity of Pinus massoniana Dependence on Climate, Soil and Forest Characteristics" Forests 11, no. 4: 404. https://doi.org/10.3390/f11040404
APA StyleHuang, X., Huang, C., Teng, M., Zhou, Z., & Wang, P. (2020). Net Primary Productivity of Pinus massoniana Dependence on Climate, Soil and Forest Characteristics. Forests, 11(4), 404. https://doi.org/10.3390/f11040404