
Exploring the Effects of Thinning on Cunninghamia lanceolata Lamb. Carbon Allocation in Southwestern China Using a Process-Based Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Collection and Sampling Methods
2.3. Model and Statistical Analysis
2.3.1. CROBAS-CL Model
2.3.2. Localization Parameters
2.3.3. Error Test
2.3.4. Flowchart of the Process for Exploring the Effects of Thinning on C. Lanceolata Carbon Allocation
3. Results
3.1. Optimum Parameters
3.2. Verification of Accuracy and Reliability of CROBAS-CL
3.3. Carbon Storage and Distribution in Trees
3.4. Dynamic Variation of Soil Carbon
3.5. Relationship between Carbon Content in the Stands, Understory, and Soil
4. Discussion
4.1. Effects of Thinning on Stands and Soil
4.2. Model Evaluation and Uncertainty Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Methods and References |
|---|---|---|
| φs | 1 | Theoretical value based on the pipe model assumption [18] |
| φc | 0.75 | Based on the conical form [18] |
| φb | 1 | As suggested by Sun and Sheng [52] |
| φt | 1 | No heartwood in coarse roots [18] |
| cb | 0.23 | From data used in this study (unpublished data) |
| ct | 1 | As suggested by Sun and Sheng [52] |
| ρs, ρb, ρt (kg/m3) | 300 | Duan et al. 2016 [53] |
| αs (m2/kg) | 0.001 | Parameter estimation based on research plots |
| αb (m2/kg) | 0.00068 | Parameter estimation based on research plots |
| αt (m2/kg) | 0.00035 | Parameter estimation based on research plots |
| αr | 0.0175 | Tian et al., 2011 [54] |
| 2z | 2.499 | Theoretical value based on Mäkelä and Sievänen, 1992 [55] |
| ξ | 0.0492 | Calculated from data in Mäkelä and Albrektson, 1992 [56] to give reasonable foliage weight when 2z = 2.499 |
| Y (kg C/kg DWa) | 0.65 | Y = fc + rg where fc = carbon content of DW = 0.45 [57] and rg = specific growth respiration rate = 0.20 [58] |
| r1 (kg C/kg DW yrb) | 0.2 | As suggested by Mäkelä [18] |
| r2 (kg C/kg DW yrb) | 0.02 | As suggested by Mäkelä [18] |
| sf | 0.25 | Based on needle lifetime of 4a |
| sr | 1 | Based on fine-root lifetime of 1a |
| ds0, db0, dt0 | 1 | By definition as presented in Mäkelä [18] |
| ds1, db1, dt1 | 0.01 | As estimated by Mäkelä [18] |
| Ψs | 1 | Theoretical value according to pipe model |
| Ψc | 0.5 | As estimated by Mäkelä [18] |
| Ψb | 0.9 | As estimated by Mäkelä [18] |
| Ψt | 0 | No heartwood in transport roots |
| an (m2/kg) | 4 | Barclay and Trofymow, 2000 [59] |
| P0 | 2.134 | X(1), parameter optimization within the range 2–4 |
| aσ | 0.02 | Yoder et al., 1994 [60] |
| k | 0.2 | Oker-Blom 1986 [61] |
| q | 1 | Trial and error |
| p | 9.7914 | X(2), parameter optimization within the range 0–10 |
| aq | 0.03912 | X(3), parameter optimization within the range 0–1 |
| m0 | 0.001 | As suggested by Mäkelä [18] and confirmed through trial and error |
| m1 | 0.01 | As suggested by Mäkelä [18] and confirmed through trial and error |
References
- Trabucco, A.; Zomer, R.J.; Bossio, D.A.; Straaten, O.V.; Verchot, L.V. Climate change mitigation through afforestation/reforestation: A global analysis of hydrologic impacts with four case studies. Agric. Ecosyst. Environ. 2008, 126, 81–97. [Google Scholar] [CrossRef]
- Rodriguez, L.C.E.; Pasalodos-Tato, M.; Diaz-Balteiro, L.; McTague, J.P. The Importance of Industrial Forest Plantations. In The Management of Industrial Forest Plantations; Managing Forest Ecosystems; Borges, J., Diaz-Balteiro, L., McDill, M., Rodriguez, L., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 33. [Google Scholar]
- Naudts, K.; Ryder, J.; McGrath, M.; Otto, J.; Chen, Y.; Valade, A.; Bellasen, V.; Berhongaray, G.; Bönisch, G.; Campioli, M.; et al. A vertically discretised canopy description for ORCHIDEE (SVN r2290) and the modifications to the energy, water and carbon fluxes. Geosci. Model Dev. 2015, 8, 2035–2065. [Google Scholar] [CrossRef] [Green Version]
- Bellassen, V.; Luyssaert, S. Managing forests in uncertain times. Nature 2014, 506, 153–155. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.V.; Ning, H.S.; Wang, R.H.; Lu, X.L.; Ji, X.M. Carbon sequestration of artificial forecast: Taking forest ecosystem of Moyu, Xinjiang as an example. Environ. Sci. Technol. 2010, 33, 27–30. [Google Scholar]
- Tong, X.; Brandt, M.; Yue, Y.; Ciais, P.; Jepsen, M.R.; Penuelas, J.; Wigneron, J.P.; Xiao, X.M.; Song, X.P.; Horion, S.; et al. Forest management in southern China generates short term extensive carbon sequestration. Nat. Commun. 2020, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Del, G.I.; Six, J.; Peressotti, A.; Cotrufo, M.F. Assessing the impact of land-use change on soil C sequestration in agricultural soils by means of organic matter fraction and stable C isotopes. Glob. Chang. Biol. 2003, 9, 1204–1213. [Google Scholar]
- Fang, J.Y.; Yang, Y.H.; Ma, W.H.; Anwar, M.; Shen, H.H. Ecosystem carbon stocks and their changes in China’s grasslands. Sci. China Life Sci. 2010, 53, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Sitzia, T.; Campagnaro, T.; Kotze, D.J.; Nardi, S.; Ertani, A. The invasion of abandoned fields by a major alien tree filters understory plant traits in novel forest ecosystems. Sci. Rep. 2018, 8, 8410. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Cui, L.L.; Tian, Z. Impact of site management on changes in soil carbon after afforestation: A review. For. Ecosyst. 2010, 12, 158–165. [Google Scholar] [CrossRef]
- Lv, H.; Liang, Z. Dynamics of soil organic carbon and dissolved organic carbon in Robina pseudoacacia forests. J. Soil Sci. Plant Nutr. 2012, 12, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Blanco, J.A. Significant increase in ecosystem C can be achieved with sustainable forest management in subtropical plantation forests. PLoS ONE 2014, 9, e89688. [Google Scholar] [CrossRef]
- Scott, N.A.; Tate, K.R.; Ross, D.J.; Parshotam, A. Processes influencing soil carbon storage following afforestation of pasture with Pinus radiata at different stocking rates in New Zealand. Soil Res. 2006, 44, 85–96. [Google Scholar] [CrossRef]
- Smith, W.N.; Grant, B.B.; Campbell, C.A.; Mcconkey, B.G.; Desjardins, R.L.; Kröbel, R.; Malhi, S.S. Crop residue removal effects on soil carbon: Measured and inter-model comparisons. Agric. Ecosyst. Environ. 2012, 161, 27–38. [Google Scholar] [CrossRef]
- Girardin, M.P.; Raulier, F.; Bernier, P.Y.; Tardif, J.C. Response of tree growth to a changing climate in boreal central Canada: A comparison of empirical, process-based, and hybrid modelling approaches. Ecol. Model. 2008, 213, 209–228. [Google Scholar] [CrossRef]
- Kirschbaum, M.; Watt, M.; Tait, A.; Ausseil, A.G.E. Future wood productivity of Pinus radiata in New Zealand under expected climatic changes. Glob. Chang. Biol. 2012, 18, 1342–1356. [Google Scholar] [CrossRef]
- Xue, H.L.; Mäkelä, A.; Valsta, L.; Vanclay, J.K.; Cao, T.J. Comparison of population-based algorithms for optimizing thinnings and rotation using a process-based growth model. Scand. J. For. Res. 2019, 34, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Mäkelä, A. A carbon balance model of growth and self-pruning in trees based on structural relationships. For. Sci. 1997, 43, 7–24. [Google Scholar]
- Kantola, A.; Mäkelä, A. Crown development in Norway spruce [Picea abies (L.) Karst.]. Trees 2004, 18, 408–421. [Google Scholar] [CrossRef]
- Liao, Z.Y.; Tian, X.L.; Xue, H.L.; Wang, B.; Sun, S.C.; Cao, T.J.; Chen, S.J.; Hou, L. Comparison of empirical and process-based methods on mortality predictions for Pinus tabulaeformis stands. J. Northeast For. Univ. 2017, 45, 51–57. (In Chinese) [Google Scholar]
- Storn, R.; Price, K. Differential evolution—A simple and efficient heuristic for global optimization over continuous spaces. J. Glob. Optim. 1997, 11, 341–359. [Google Scholar] [CrossRef]
- Sun, S.C.; Cao, Q.V.; Cao, T.J. Characterizing diameter distributions for uneven-aged pine-oak mixed forests in the Qinling Mountains of China. Forests 2019, 10, 596. [Google Scholar] [CrossRef] [Green Version]
- Wilnhammer, M.; Rothe, A.; Weis, W.; Wittkopf, S. Estimating forest biomass supply from private forest owners: A case study from Southern Germany. Biomass Bioenergy 2012, 47, 177–187. [Google Scholar] [CrossRef]
- Fichtner, A.; Forrester, D.I.; Härdtle, W.; Sturm, K.; Von, O.G. Facilitative-competitive interactions in an old-growth forest: The importance of large-diameter trees as benefactors and stimulators for forest community assembly. PLoS ONE 2015, 10, e0120335. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.W.; Huang, Y.L.; Chen, J.C.; Chen, C.T. Habitat environment data and potential habitat interpolation of Cyathea lepifera at the Tajen experimental forest station in Taiwan. Trop. Conserv. Sci. 2016, 9, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Russell, T.G.; Alan, E.H.; Theresa, B.J.; Jonalea, R.T. The Effects of Thinning and Similar Stand Treatments on Fire Behavior in Western Forests; General Technical Report PNW-GTR-463; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1999. [Google Scholar]
- Ming, A.G.; Yang, Y.J.; Liu, S.R.; Wang, H.; Li, Y.F.; Li, H.; Nong, Y.; Cai, D.X.; Jia, H.Y.; Tao, Y.; et al. Effects of near natural forest management on soil greenhouse gas flux in Pinus massoniana (lamb.) and Cunninghamia lanceolata (lamb.) hook. plantations. Forests 2018, 9, 229. [Google Scholar] [CrossRef] [Green Version]
- Penman, J.; Gytarsky, M.; Hiraishi, T.; Krug, T.; Kruger, D.; Pipatti, R.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K.; et al. Good Practice Guidance for Land Use, Land-Use Change and Forestry; The Institute for Global Environmental Strategies, The Intergovernmental Panel on Climate Change (IPCC): Hayama Kanagawa, Japan, 2003; pp. 1.4–1.10. [Google Scholar]
- Johann, F.; Schaich, H. Land ownership affects diversity and abundance of tree microhabitats in deciduous temperate forests. For. Ecol. Manag. 2016, 380, 70–81. [Google Scholar] [CrossRef]
- Yu, D.; Wang, X.; Yin, Y.; Zhan, J.; Lewis, B.J.; Tian, J.; Bao, Y.; Zhou, W.; Zhou, L.; Dai, L. Estimates of forest biomass carbon storage in Liaoning Province of Northeast China: A review and assessment. PLoS ONE 2014, 9, e89572. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Miao, N.; Li, S.C.; Ma, R.; Liao, Z.Y.; Wang, W.P.; Sun, H.L. Relationship between stand characteristics and soil properties of two typical forest plantations in the mountainous area of western sichuan, China. J. Mt. Sci. 2019, 16, 1816–1832. [Google Scholar] [CrossRef]
- Wilke, B.M.; Margesin, R.; Schinner, F. Determination of Chemical and Physical Soil Properties: Monitoring and Assessing Soil Bioremediation; Biology, S., Margesin, R., Schinner, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Schlesinger, W.H.; Lichter, J. Limited carbon storage in soil and litter of experimental forest plots under increased atmospheric CO2. Nature 2001, 411, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.K.; Astley, H.; Smith, P.; Ghoshal, N. Soil CO2–C flux and carbon storage in the dry tropics: Impact of land-use change involving bioenergy crop plantation. Biomass Bioenergy 2015, 83, 123–130. [Google Scholar] [CrossRef]
- Shipunov, A.; Balandin, P.A.; Volkova, A.I.; Korobejnikov, S.A.; Nazarova, S.V.P.; Sufijanov, V.G. Visual Statistics. Use R; DMK Press: Moscow, Russia, 2012; p. 429. Available online: http://ashipunov.info/shipunov/school/biol_240/en/ (accessed on 17 March 2019).
- Lin, S.Z.; Cao, G.Q.; Du, L.; Wang, A.P. Effect of allelochemicals of Chinese-fir root extracted by supercritical CO2 extraction on Chinese fir. J. Appl. Ecol. 2003, 14, 122–126. [Google Scholar]
- Huang, J.X.; Lin, T.C.; Xiong, D.C.; Yang, Z.J.; Liu, X.F.; Chen, G.S.; Xie, J.S.; Li, Y.Q.; Yang, Y.S. Organic carbon mineralization in soils of a natural forest and a forest plantation of southeastern China. Geoderma 2019, 344, 119–126. [Google Scholar] [CrossRef]
- Chatskikh, D.; Olesen, J.E. Soil tillage enhanced CO and NO emissions from loamy sand soil under spring barley. Soil Tillage Res. 2007, 97, 5–18. [Google Scholar] [CrossRef]
- Cao, J.; Wang, X.; Yun, T.; Wen, Z.; Zha, T. Pattern of carbon allocation across three different stages of stand development of a Chinese pine (Pinus tabulaeformis) forest. Ecol. Res. 2012, 27, 883–892. [Google Scholar] [CrossRef]
- Verma, A.; Kaushal, R.; Alam, N.M.; Mehta, H.; Chaturvedi, O.P.; Mandal, D.; Tomar, J.M.S.; Rathore, A.C.; Singh, C. Predictive models for biomass and carbon stock estimation in Grewia optiva on degraded lands in western Himalaya. Agrofor. Syst. 2014, 88, 895–905. [Google Scholar] [CrossRef]
- Zhao, J.; Kang, F.; Wang, L.; Yu, X.; Zhao, W.; Song, X.; Zhang, Y.; Chen, F.; Sun, Y.; He, T. Patterns of biomass and carbon distribution across a chronosequence of Chinese pine (Pinus tabulaeformis) forests. PLoS ONE 2014, 9, e94966. [Google Scholar] [CrossRef] [PubMed]
- Angélico, T.d.S.; Marcati, C.R.; Rossi, S.; Silva, M.R.; Sonsin-Oliveira, J. Soil effects on stem growth and wood anatomy of tamboril are mediated by tree age. Forests 2021, 12, 1058. [Google Scholar] [CrossRef]
- Cook, R.L.; Dan, B.; Mendes, J.C.T.; Stape, J.L. Soil carbon stocks and forest biomass following conversion of pasture to broadleaf and conifer plantations in southeastern Brazil. For. Ecol. Manag. 2014, 324, 37–45. [Google Scholar] [CrossRef]
- Davis, J.P.; Haines, B.; Coleman, D.; Hendrick, R. Fine root dynamics along an elevational gradient in the southern Appalachian Mountains, USA. For. Ecol. Manag. 2004, 187, 19–33. [Google Scholar] [CrossRef]
- Medeiros, R.A.; Paiva, H.N.D.; Soares, A.; Cruz, J.P.D.; Leite, H.G. Thinning from below: Effects on height of dominant trees and diameter distribution in Eucalyptus stands. J. Trop. For. Sci. 2017, 29, 238–247. [Google Scholar]
- Minunno, F.; Peltoniemi, M.; Hrknen, S.; Kalliokoski, T.; Mkel, A. Bayesian calibration of a carbon balance model PREBAS using data from permanent growth experiments and national forest inventory. For. Ecol. Manag. 2019, 440, 208–257. [Google Scholar] [CrossRef]
- Blanco, J.A.; Seely, B.; Welham, C.; Kimmins, J.P.H.; Seebacher, T.M. Testing the performance of a forest ecosystem model (FORECAST) against 29 years of field data in a Pseudotsuga menziesii plantation. Can. J. For. Res. 2007, 37, 1808–1820. [Google Scholar] [CrossRef]
- Jyske, M.; Kalliokoski, N. Intra-annual tracheid production of Norway spruce and Scots pine across a latitudinal gradient in Finland. Agric. For. Meteorol. 2014, 194, 241–254. [Google Scholar] [CrossRef]
- Tian, X.; Minunno, F.; Cao, T.; Peltoniemi, M.; Kalliokoski, T.; Mkel, A. Extending the range of applicability of the semi-empirical ecosystem flux model PRELES for varying forest types and climate. Glob. Chang. Biol. 2020, 26, 2923–2943. [Google Scholar] [CrossRef]
- Valentine, H.T.; Annikki, M. Bridging process-based and empirical approaches to modeling tree growth. Tree Physiol. 2005, 25, 769–779. [Google Scholar] [CrossRef]
- Stage, A.R.; Wykoff, W.R. Adapting distance-independent forest growth models to represent spatial variability: Effects of sampling design on model coefficients. For. Sci. 1998, 44, 224–238. [Google Scholar]
- Sun, C.Z.; Sheng, G.F. Study on the present condition and the potentialities of the productivity of main tree species forest plantation of China I.Study on the forest plantation productivities of Cunninghamia lanceolata and Pinus massoniana. For. Res. 2000, 13, 613–621. [Google Scholar]
- Duan, H.; Cao, S.; Zheng, H.; Hu, D.; Lin, J.; Lin, H.; Hu, R.; Sun, Y.; Li, Y. Variation in the growth traits and wood properties of chinese fir from six provinces of southern China. Forests 2016, 7, 192. [Google Scholar] [CrossRef] [Green Version]
- Tian, D.; Xiang, W.; Chen, X.; Yan, W.; Peng, Y. A long-term evaluation of biomass production in first and second rotations of Chinese fir plantations at the same site. Forestry 2011, 84, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Mäkelä, A.; Sievanen, R. Height growth strategies in open-grown trees. J. Theor. Biol. 1992, 159, 443–467. [Google Scholar] [CrossRef]
- Mäkelä, A.; Albrektson, A. An analysis of the relationship between foliage biomass and crown surface area in Pinus sylvestris in Sweden. Scand. J. For. Res. 1992, 7, 297–307. [Google Scholar] [CrossRef]
- Tao, Y.H.; Feng, J.C.; Cao, S.G.; Guo, Q.; Xiang, D.Y. Study on carbon storage of Pinus massoniana, Cunninghamia lanceolata plantations at Shatang, Guangxi Province. J. Northwest Agric. For. Univ. (Nat. Sci. Ed.) 2013, 40, 38–44. (In Chinese) [Google Scholar]
- Lavigne, M.B.; Franklin, S.E.; Hunt, E.R. Estimating stem maintenance respiration rates of dissimilar balsam fir stands. Tree Physiol. 1996, 16, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Barclay, H.J.; Trofymow, J.A. Relationship of readings from the LI-COR canopy analyzer to total one-sided leaf area index and stand structure in immature Douglas-fir. For. Ecol. Manag. 2000, 132, 121–126. [Google Scholar] [CrossRef]
- Yoder, B.; Ryan, M.; Waring, R.; Schoettle, A.; Kaufmann, M. Evidence of reduced photosynthetic rates in old trees. For. Sci. 1994, 40, 513–527. [Google Scholar]
- Oker-Blom, P. Photosynthetic radiation regime and canopy structure in modeled forest stands. Acta For. Fenn. 1986, 197, 1–44. [Google Scholar] [CrossRef]


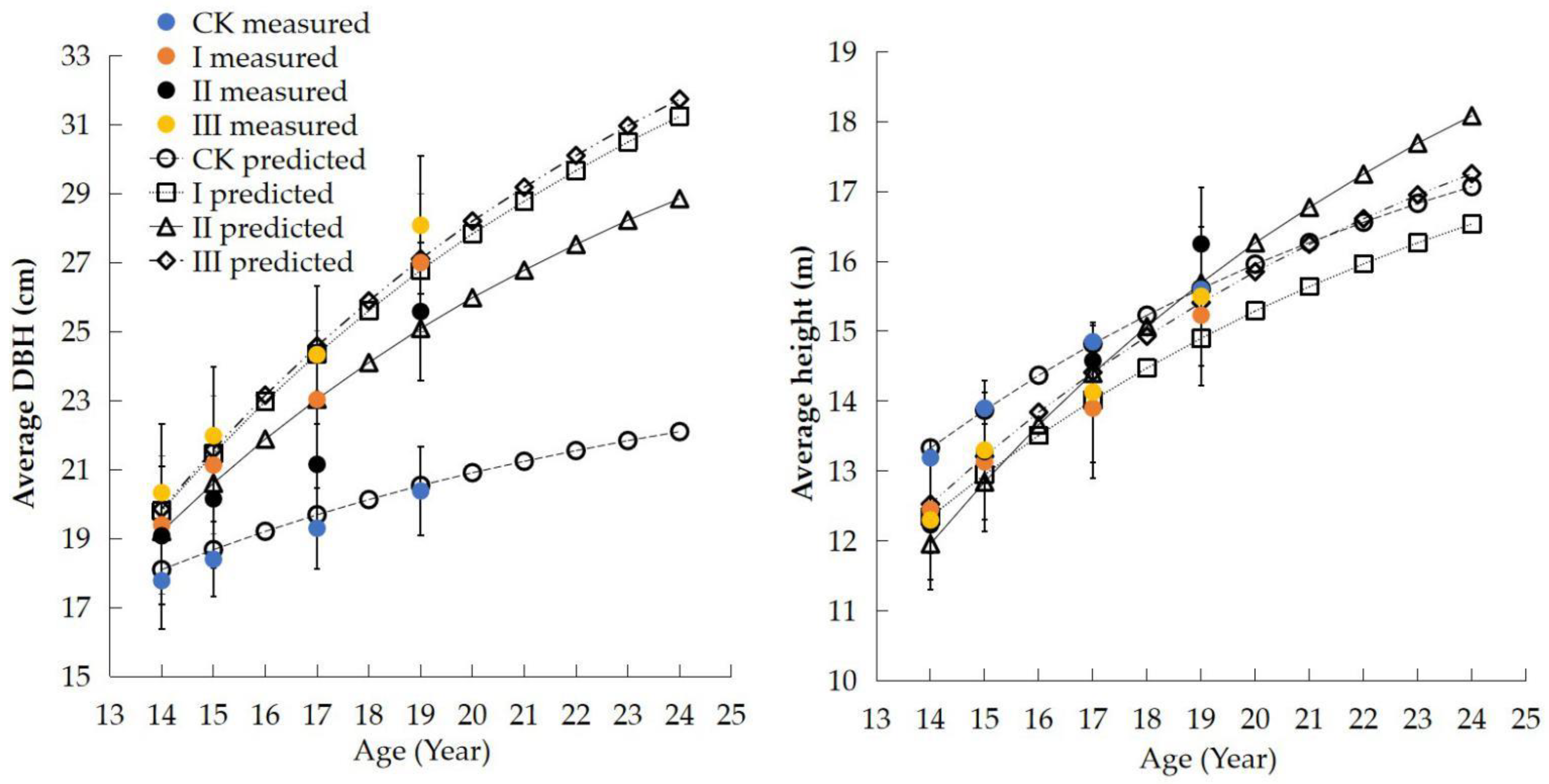

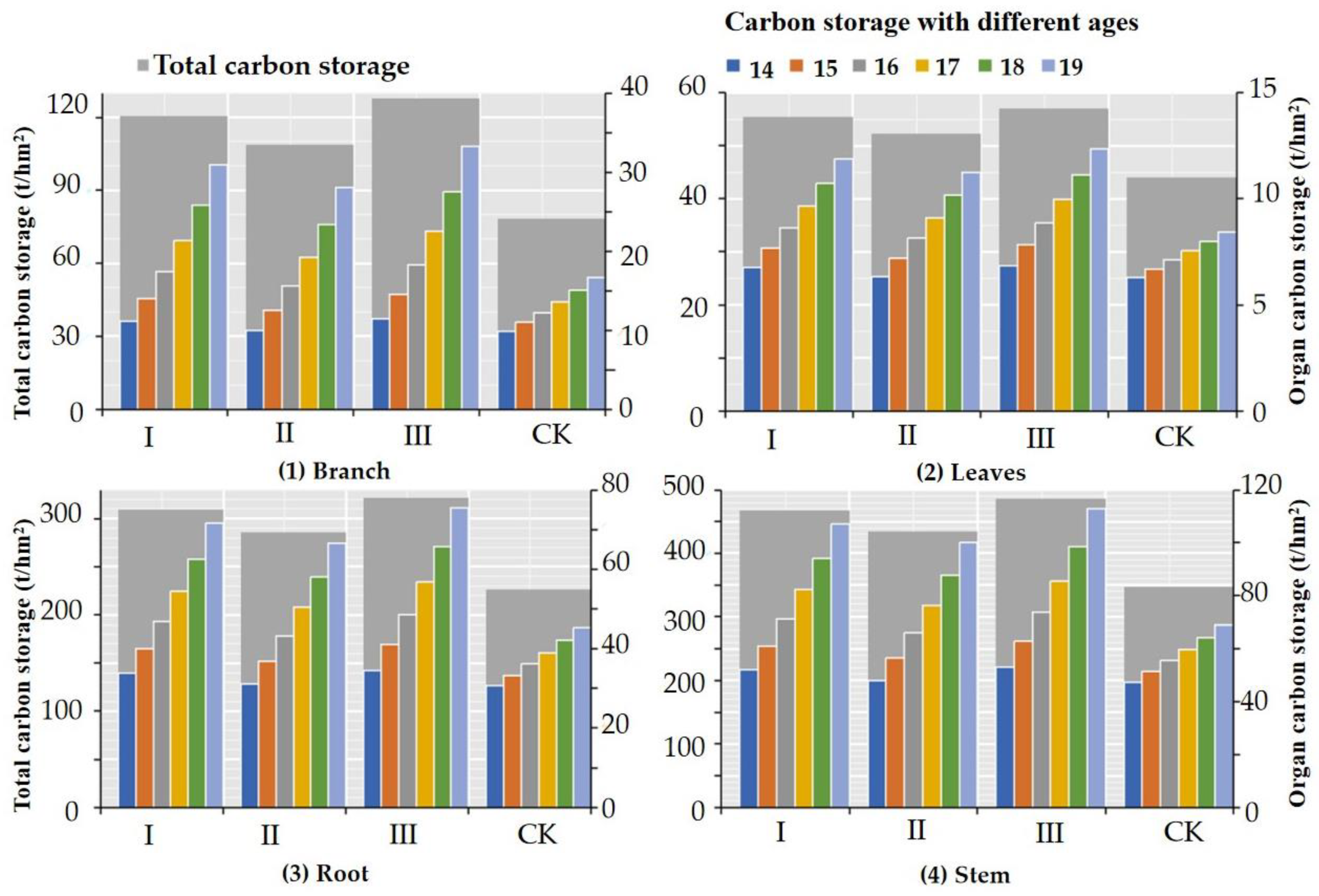

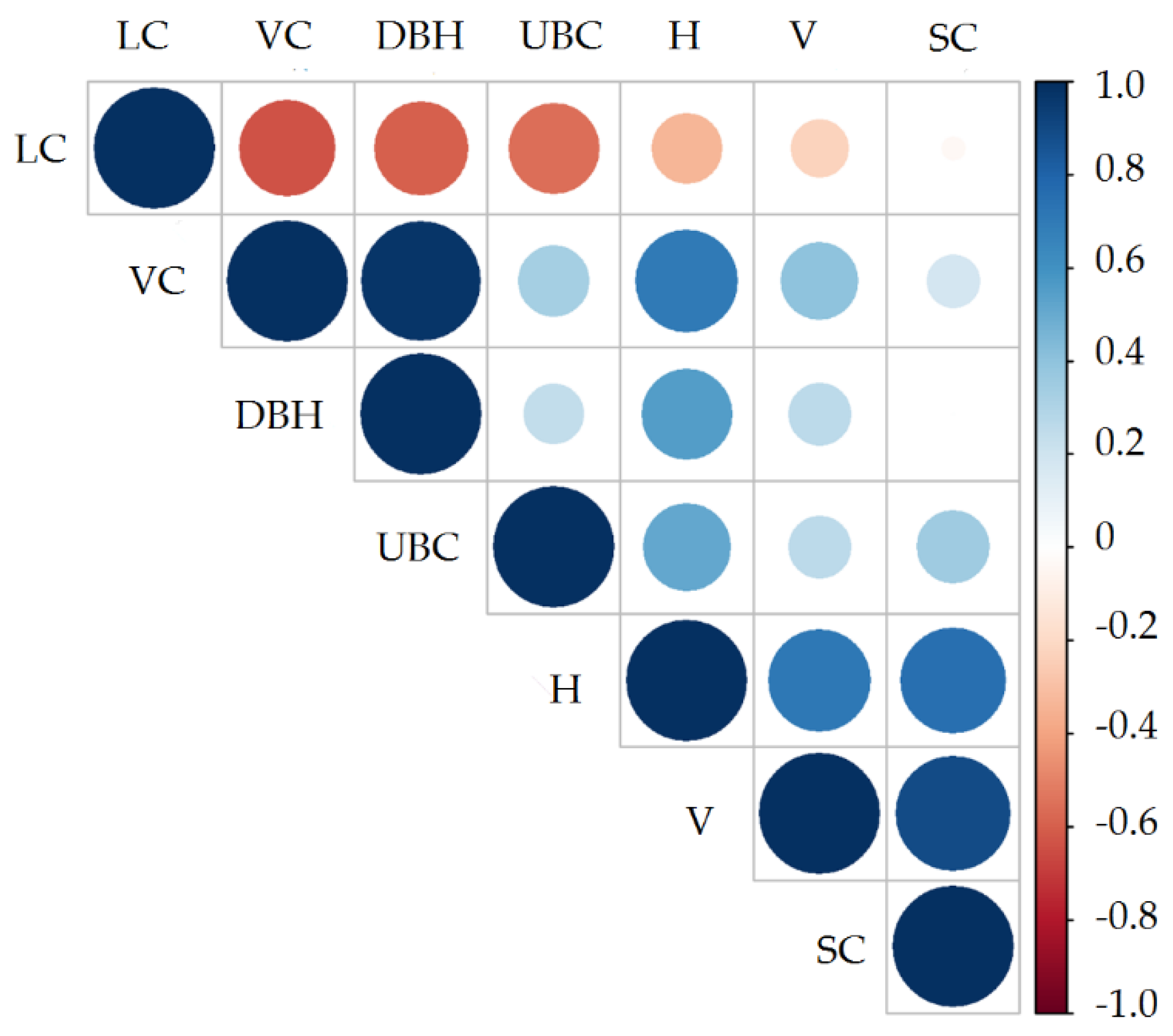
| Item | Age (Year) | Thinning-I | Thinning-II | Thinning-III | CK Treatment |
|---|---|---|---|---|---|
| DBH (cm) | 14 | 19.40 ± 0.59 ab | 19.08 ± 1.80 ab | 20.33 ± 0.67 a | 17.78 ± 1.39 a |
| 15 | 21.13 ± 0.43 bc | 20.15 ± 1.65 ab | 21.98 ± 0.66 b | 18.40 ± 1.09 b | |
| 17 | 23.03 ± 1.79 bc | 21.15 ± 2.44 bc | 24.33 ± 1.15 c | 19.30 ± 1.17 b | |
| 19 | 27 ± 1.64 abc | 25.58 ± 2.55 c | 28.08 ± 1.08 d | 20.38 ± 1.29 b | |
| Height (m) | 14 | 12.45 ± 1.02 a | 12.23 ± 0.40 a | 12.30 ± 0.59 a | 13.19 ± 1.06 a |
| 15 | 13.13 ± 0.70 a | 13.28 ± 0.40 b | 13.30 ± 0.47 b | 13.90 ± 0.75 ab | |
| 17 | 13.90 ± 0.69 ab | 14.58 ± 0.51 c | 14.13 ± 0.41 b | 14.85 ± 0.70 bc | |
| 19 | 15.23 ± 0.67 c | 16.25 ± 0.81 d | 15.50 ± 0.91 c | 15.6 ± 0.79 cd | |
| Single-tree biomass (kg per tree) | 14 | 113.05 ± 11.44 a | 145.60 ± 9.80 a | 188.72 ± 31.04 ab | 100.02 ± 21.28 a |
| 15 | 107.80 ± 22.77 a | 133.88 ± 26.58 a | 165.97 ± 38.83 a | 113.78 ± 14.22 ab | |
| 17 | 123.95 ± 10.97 a | 160.96 ± 8.50 a | 216.02 ± 18.04 ab | 136.42 ± 18.53 bc | |
| 19 | 134.74 ± 35.74 a | 161.90 ± 42.14 a | 223.72 ± 42.75 b | 162.75 ± 19.32 c |
| Fitting Results | Parameters | Object Value | ||
|---|---|---|---|---|
| X(1) | X(2) | X(3) | ||
| CK | 2.134 | 9.791 | 0.039 | 0.320 |
| I | 3.787 | 8.295 | 0.089 | 0.261 |
| II | 2.671 | 8.998 | 0.098 | 0.422 |
| III | 2.671 | 8.999 | 0.099 | 0.413 |
| Thinning Type | MAD | RMSE (%) | ||
|---|---|---|---|---|
| H | DBH | H | DBH | |
| CK | 0.053 | 0.056 | 0.004 | 0.003 |
| I | 0.059 | 0.159 | 0.005 | 0.008 |
| II | 0.087 | 0.134 | 0.007 | 0.007 |
| III | 0.067 | 0.165 | 0.006 | 0.008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Liao, Z.; Ming, A.; Miao, N. Exploring the Effects of Thinning on Cunninghamia lanceolata Lamb. Carbon Allocation in Southwestern China Using a Process-Based Model. Forests 2021, 12, 1590. https://doi.org/10.3390/f12111590
Yang H, Liao Z, Ming A, Miao N. Exploring the Effects of Thinning on Cunninghamia lanceolata Lamb. Carbon Allocation in Southwestern China Using a Process-Based Model. Forests. 2021; 12(11):1590. https://doi.org/10.3390/f12111590
Chicago/Turabian StyleYang, Hao, Ziyan Liao, Angang Ming, and Ning Miao. 2021. "Exploring the Effects of Thinning on Cunninghamia lanceolata Lamb. Carbon Allocation in Southwestern China Using a Process-Based Model" Forests 12, no. 11: 1590. https://doi.org/10.3390/f12111590