
Effects of Afforestation on Plant Diversity and Soil Quality in Semiarid SE Spain



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Statistical Analyses
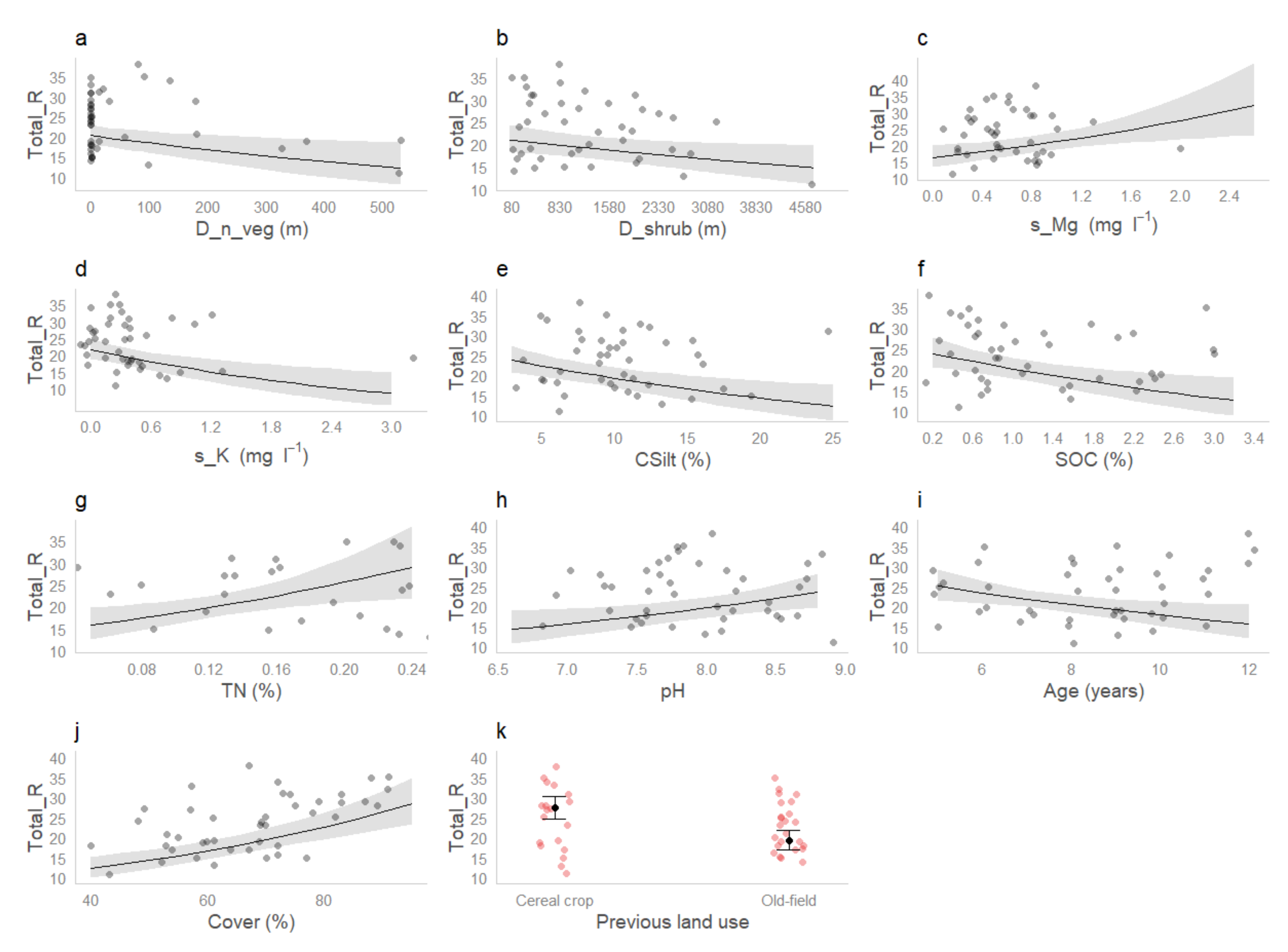
3. Results
3.1. Environmental Characteristics and Plant Species in Afforested Sites
3.2. Effect of the Previous Land Use on Plant Diversity and Environmental Variables
3.3. Effects of Environmental Variables and Soil Properties on Plant Diversity
3.4. Diversity and Soil Properties Comparison between Afforestation and Cereal Crops
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seddon, N.; Smith, A.; Smith, P.; Key, I.; Chausson, A.; Girardin, C.; House, J.; Srivastava, S.; Turner, B. Getting the message right on nature-based solutions to climate change. Glob. Chang. Biol. 2021, 27, 1518–1546. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organisation of the United Nations. Global Forest Resources Assessment 2020—Key Findings; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Castro, J.; Morales-Rueda, F.; Navarro, F.B.; Löf, M.; Vacchiano, G.; Alcaraz-Segura, D. Precision restoration: A necessary approach to foster forest recovery in the 21st century. Restor. Ecol. 2021, 29, e13421. [Google Scholar] [CrossRef]
- Keesstra, S.D.; Bouma, J.; Wallinga, J.; Tittonell, P.; Smith, P.; Cerdà, A.; Montanarella, L.; Quinton, J.N.; Pachepsky, Y.; van der Putten, W.H.; et al. The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. Soil 2016, 2, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Meli, P.; Holl, K.D.; Benayas, J.M.R.; Jones, H.P.; Jones, P.C.; Montoya, D.; Mateos, D.M. A global review of past land use, climate, and active vs. passive restoration effects on forest recovery. PLoS ONE 2017, 12, e0171368. [Google Scholar] [CrossRef]
- Bremer, L.L.; Farley, K.A. Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodivers. Conserv. 2010, 19, 3893–3915. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.M.; Navarro, L.M. Rewilding European Landscapes; Springer: Cham, Switzerland, 2015; ISBN 9783319120393. [Google Scholar]
- Jones, H.; Jones, P.C.; Barbier, E.B.; Blackburn, R.C.; Rey Benayas, J.M.; Holl, K.D.; McCrakin, M.; Meli, P.; Montoya, D.; Moreno Mateos, D. Restoration and repair of Earth’s damaged ecosystems. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172577. [Google Scholar] [CrossRef]
- Derak, M.; Cortina, J. Multi-criteria participative evaluation of Pinus halepensis plantations in a semiarid area of southeast Spain. Ecol. Indic. 2014, 43, 56–68. [Google Scholar] [CrossRef]
- Fuss, S.; Lamb, W.F.; Callaghan, M.W.; Hilaire, J.; Creutzig, F.; Amann, T.; Beringer, T.; De Oliveira Garcia, W.; Hartmann, J.; Khanna, T.; et al. Negative emissions—Part 2: Costs, potentials and side effects. Environ. Res. Lett. 2018, 13, 63002. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Qiao, W.; Gao, D.; Dai, Y.; Deng, J.; Yang, G.; Han, X.; Ren, G. Relationship between soil nutrient properties and biological activities along a restoration chronosequence of Pinus tabulaeformis plantation forests in the Ziwuling Mountains, China. Catena 2018, 161, 85–95. [Google Scholar] [CrossRef]
- Andrés, C.; Ojeda, F. Effects of afforestation with pines on woody plant diversity of mediterranean heathlands in southern Spain. Biodivers. Conserv. 2002, 11, 1511–1520. [Google Scholar] [CrossRef]
- Laganiere, J.; Angers, D.A.; Pare, D. Carbon accumulation in agricultural soils after afforestation: A meta-analysis. Glob. Chang. Biol. 2010, 16, 439–453. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Curran, M.; Ferreira, M.S.; Lindenmayer, D.B.; Grelle, C.E.V.; Rey Benayas, J.M. A global meta-Analysis on the ecological drivers of forest restoration success. Nat. Commun. 2016, 7, 11666. [Google Scholar] [CrossRef]
- Huang, C.; Zeng, Y.; Wang, L.; Wang, S. Responses of soil nutrients to vegetation restoration in China. Reg. Environ. Chang. 2020, 20, 82. [Google Scholar] [CrossRef]
- Plaza, C.; Zaccone, C.; Sawicka, K.; Méndez, A.M.; Tarquis, A.; Gascó, G.; Heuvelink, G.B.M.; Schuur, E.A.G.; Maestre, F.T. Soil resources and element stocks in drylands to face global issues. Sci. Rep. 2018, 8, 13788. [Google Scholar] [CrossRef]
- Kouba, Y.; Martínez-García, F.; De Frutos, Á.; Alados, C.L. Effects of previous land-use on plant species composition and diversity in Mediterranean forests. PLoS ONE 2015, 10, e0139031. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J. Are Pinus halepensis plantations useful as a restoration tool in semiarid Mediterranean areas? For. Ecol. Manag. 2004, 198, 303–317. [Google Scholar] [CrossRef]
- Chen, L.F.; He, Z.B.; Zhu, X.; Du, J.; Yang, J.J.; Li, J. Impacts of afforestation on plant diversity, soil properties, and soil organic carbon storage in a semi-arid grassland of northwestern China. Catena 2016, 147, 300–307. [Google Scholar] [CrossRef]
- Jiao, J.; Zhang, Z.; Bai, W.; Jia, Y.; Wang, N. Assessing the Ecological Success of Restoration by Afforestation on the Chinese Loess Plateau. Restor. Ecol. 2012, 20, 240–249. [Google Scholar] [CrossRef]
- Zhao, F.; Kang, D.; Han, X.; Yang, G.; Yang, G.; Feng, Y.; Ren, G. Soil stoichiometry and carbon storage in long-term afforestation soil affected by understory vegetation diversity. Ecol. Eng. 2015, 74, 415–422. [Google Scholar] [CrossRef]
- Solomou, A.D.; Skoufogianni, E.; Danalatos, N.G. Exploitation of soil properties for controlling herbaceous plant communities in an organic cultivation of lippia citriodora in the mediterranean landscape. Bulg. J. Agric. Sci. 2020, 26, 79–83. [Google Scholar]
- Cuesta, B.; Rey Benayas, J.M.; Gallardo, A.; Villar-Salvador, P.; González-Espinosa, M. Soil chemical properties in abandoned Mediterranean cropland after succession and oak reforestation. Acta Oecol. 2012, 38, 58–65. [Google Scholar] [CrossRef]
- Segura, C.; Navarro, F.B.; Jiménez, M.N.; Fernández-Ondoño, E. Implications of afforestation vs. secondary succession for soil properties under a semiarid climate. Sci. Total Environ. 2020, 704, 135393. [Google Scholar] [CrossRef]
- Heydari, M.; Zeynali, N.; Bazgir, M.; Omidipour, R.; Kohzadian, M.; Sagar, R.; Prevosto, B. Rapid recovery of the vegetation diversity and soil fertility after cropland abandonment in a semiarid oak ecosystem: An approach based on plant functional groups. Ecol. Eng. 2020, 155, 105963. [Google Scholar] [CrossRef]
- Hulshof, C.M.; Spasojevic, M.J. The edaphic control of plant diversity. Glob. Ecol. Biogeogr. 2020, 29, 1634–1650. [Google Scholar] [CrossRef]
- Kleijn, D.; Sutherland, W.J. How effective are European agri-environment schemes in conserving and promoting biodiversity? J. Appl. Ecol. 2003, 40, 947–969. [Google Scholar] [CrossRef]
- Kleijn, D.; Baquero, R.A.; Clough, Y.; Díaz, M.; De Esteban, J.; Fernández, F.; Gabriel, D.; Herzog, F.; Holzschuh, A.; Jöhl, R.; et al. Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecol. Lett. 2006, 9, 243–254. [Google Scholar] [CrossRef]
- Batáry, P.; Dicks, L.V.; Kleijn, D.; Sutherland, W.J. The role of agri-environment schemes in conservation and environmental management. Conserv. Biol. 2015, 29, 1006–1016. [Google Scholar] [CrossRef] [Green Version]
- Zanchi, A.G.; Thiel, D.; Green, T.; Lindner, M. Afforestation in Europe; European Forest Institute: Joensuu, Finland, 2007. [Google Scholar]
- Fuchs, R.; Herold, M.; Verburg, P.H.; Clevers, J.G.P.W. A high-resolution and harmonized model approach for reconstructing and analysing historic land changes in Europe. Biogeosciences 2013, 10, 1543–1559. [Google Scholar] [CrossRef] [Green Version]
- Vadell, E.; De Miguel, S.; Fernández Centeno, G.; Robla, E.; Lerner, M.; Pemán García, J. Afforestation of agricultural lands: Balance of a forest policy instrument for land-use change. Cuadernos de la Sociedad Española de Ciencias Forestales 2019, 45, 1–20. [Google Scholar] [CrossRef]
- Madsen, L.M. The Danish afforestation programme and spatial planning: New challenges. Landsc. Urban Plan. 2002, 58, 241–254. [Google Scholar] [CrossRef]
- Molina, C.M. The Restoration of Forest Landscapes. In The Conservation of Cultural Landscapes; Agnoletti, M., Ed.; CAB International: Wallingford, UK, 2006; pp. 195–208. [Google Scholar]
- Horák, J.; Brestovanská, T.; Mladenović, S.; Kout, J.; Bogusch, P.; Halda, J.P.; Zasadil, P. Green desert?: Biodiversity patterns in forest plantations. For. Ecol. Manag. 2019, 433, 343–348. [Google Scholar] [CrossRef]
- Nunes, A.; Oliveira, G.; Mexia, T.; Valdecantos, A.; Zucca, C.; Costantini, E.A.C.; Abraham, E.M.; Kyriazopoulos, A.P.; Salah, A.; Prasse, R.; et al. Ecological restoration across the Mediterranean Basin as viewed by practitioners. Sci. Total Environ. 2016, 566–567, 722–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantini, E.A.C.; Branquinho, C.; Nunes, A.; Schwilch, G.; Stavi, I.; Valdecantos, A.; Zucca, C. Soil indicators to assess the effectiveness of restoration strategies in dryland ecosystems. Solid Earth 2016, 7, 397–414. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Carreras, J. The effect of bedrock type, temperature and moisture on species richness of Pyrenean Scots pine (Pinus sylvestris L.) forests. Vegetatio 1995, 116, 85–92. [Google Scholar] [CrossRef]
- Gleixner, G.; Kramer, C.; Hahn, V.; Sachse, D. The effect of biodiversity on carbon storage in soils. In Forest Diversity and Function: Temperate and Boreal Systems; Scherer-Lorenzen, M., Korner, C., Schulze, E.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 165–183. [Google Scholar]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Ochoa, V.; Delgado-Baquerizo, M.; García-Gómez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; et al. Plant species richness and ecosystem multifunctionality in global drylands. Science 2012, 335, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Borja, M.E.; Delgado-Baquerizo, M. Plant diversity and soil stoichiometry regulates the changes in multifunctionality during pine temperate forest secondary succession. Sci. Total Environ. 2019, 697, 134204. [Google Scholar] [CrossRef]
- Rivas-Martinez, S.; Loidi, J. Bioclimatology of the Iberian Peninsula. Itinera Geobot. 1999, 13, 41–47. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 1998; Report Num. 84; IUSS Working Group WRB: Roma, Italy, 1998. [Google Scholar]
- Chalmers, N.; Parker, P. The OU Project Guide: Fieldwork and Statistics for Ecological Projects; The Dorset Press: Dorchester, UK, 1989. [Google Scholar]
- Cabezudo, B.; Talavera, S.; Blanca, G.; Salazar, C.; Cueto, M.; Valdés, B.; Hernández-Bermejo, J.E.; Herrera, C.M.; Hiraldo, C.R.; Navas, D. Lista Roja de la Flora Vascular Amenazada de Andalucía; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2005. [Google Scholar]
- Soil Conservation Service. Soil Survey Laboratory Methods and Procedures for Collecting Soils Samples; Soil Conservation Service: Washington, DC, USA, 1972.
- Cassel, D.K.; Nielsen, D.R. Fields capacity and available water capacity. In Methods of Soil Analysis; Part. 1: Physical and Mineralogical Methods; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; pp. 901–926. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of Degtjareff method for determining soil organic matter and a proposed modification of the cromic titration method. Soil Sci. 1934, 34, 29–38. [Google Scholar] [CrossRef]
- Tyurin, I.V. Analytical procedure for a comparature study of soil humus. Tr. Pochr. Inst. Dokuchaeva 1951, 33, 5–21. [Google Scholar]
- Bremner, J.M. Nitrogen availability indexes. In Methods of Soil Analysis; Part. 2. Chemical and Microbiological Properties; Black, C.A., Evans, D.D., Esminger, T.E., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1324–1345. [Google Scholar]
- Williams, D.E. A rapid manometric method for the determination of carbonate in soils. Soil Sci. Soc. Am. Proc. 1948, 13, 27–129. [Google Scholar] [CrossRef] [Green Version]
- US Salinity Laboratory. Diagnosis and Improvement of Saline and Alkali Soils; US Department of Agriculture: Washington, DC, USA, 1954.
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis: Chemical and Microbiological Properties; Page, D.L., Ed.; American Society of Agronomy; Soil Science Society of America: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Consejería de Medio. Ambiente Mapa digital de Usos y Coberturas Vegetales del Suelo de Andalucía a Escala 1:25.000; Junta de Andalucía: Sevilla, Spain, 2003.
- Sánchez-Palomares, O.; Sánchez Serrano, F.; Carretero-Carrero, M.P. Modelos y Cartografía de Estimaciones Climáticas Termopluviométricas Para la España Peninsular; INIA: Madrid, Spain, 1999. [Google Scholar]
- IGME. Mapa Geológico de España a Escala 1:50.000; Instituto Geológico y Minero de España: Madrid, Spain, 1980.
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002; ISBN 0387-95457-0. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; pp. 27–41. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84); 2017. Available online: https://github.com/taiyun/corrplot (accessed on 7 December 2021).
- Lüdecke, D. ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. J. Open Source Softw. 2018, 3, 772. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage Publications: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: http://www.r-project.org (accessed on 29 September 2021).
- Blanca, G.; Cabezudo, B.; Cueto, M.; Salazar, C.; Morales Torres, C. Flora Vascular de Andalucía Oriental, 2nd ed.; Junta de Andalucía: Sevilla, Spain, 2011; Volume 4.
- Kleijn, D.; Joenje, W.; Le Coeur, D.; Marshall, E.J.P. Similarities in vegetation development of newly established herbaceous strips along contrasting European field boundaries. Agric. Ecosyst. Environ. 1998, 68, 13–26. [Google Scholar] [CrossRef]
- Sanz Herraiz, C.; López Estébanez, N.; Molina Holgado, P. Influencia de las repoblaciones forestales en la evolución de las comunidades vegetales y orníticas de la Sierra de los Filabres (Almería). Ería 2002, 58, 157–176. [Google Scholar] [CrossRef]
- Médail, F.; Quezél, P. Linked references are available on JSTOR for this article: Biodiversity Hotspots in the Mediterranean Basin: Setting Global Conservation Priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.; Deng, J.; Wang, B. Afforestation affects soil seed banks by altering soil properties and understory plants on the eastern Loess Plateau, China. Ecol. Indic. 2021, 126, 107670. [Google Scholar] [CrossRef]
- Wulf, M. Plant species richness of afforestations with different former use and habitat continuity. For. Ecol. Manag. 2004, 195, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Janssens, F.; Peeters, A.; Tallowin, J.R.B.; Bakker, J.P.; Bekker, R.M.; Fillat, F. Relationship between soil chemical factors and grassland diversity. Plant Soil 1998, 202, 69–78. [Google Scholar] [CrossRef]
- Liang, W.; Wei, X. Relationships between ecosystems above and below ground including forest structure, herb diversity and soil properties in the mountainous area of Northern China. Glob. Ecol. Conserv. 2020, 24, e01228. [Google Scholar] [CrossRef]
- Hrivnák, R.; Slezák, M.; Jarcuška, B.; Jarolímek, I.; Kochjarová, J. Native and alien plant species richness response to soil nitrogen and phosphorus in temperate floodplain and swamp forests. Forests 2015, 6, 3501–3513. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.; Austin, M. Patterns of plant species richness in relation to different environments: An appraisal. J. Veg. Sci. 2001, 12, 153–166. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, T.; Guo, J.; Tan, Z.; Dong, W.; Wang, H.H. Changes in the understory diversity of secondary Pinus tabulaeformis forests are the result of stand density and soil properties. Glob. Ecol. Conserv. 2021, 28, e01628. [Google Scholar] [CrossRef]
- Basnou, C.; Vicente, P.; Espelta, J.M.; Pino, J. Of niche differentiation, dispersal ability and historical legacies: What drives woody community assembly in recent Mediterranean forests? Oikos 2016, 125, 107–116. [Google Scholar] [CrossRef]
- Abadie, J.; Avon, C.; Dupouey, J.-L.; Lopez, J.-M.; Tatoni, T.; Bergès, L. Land use legacies on forest understory vegetation and soils in the Mediterranean region: Should we use historical maps or in situ land use remnants? For. Ecol. Manag. 2018, 427, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Amici, V.; Santi, E.; Filibeck, G.; Diekmann, M.; Geri, F.; Landi, S.; Scoppola, A.; Chiarucci, A. Influence of secondary forest succession on plant diversity patterns in a Mediterranean landscape. J. Biogeogr. 2013, 40, 2335–2347. [Google Scholar] [CrossRef]
- Cortina, J.; Amat, B.; Castillo, V.; Fuentes, D.; Maestre, F.T.; Padilla, F.M.; Rojo, L. The restoration of vegetation cover in the semi-arid Iberian southeast. J. Arid Environ. 2011, 75, 1377–1384. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yang, T.; Wang, Q.; Huang, F.; Li, L. Dynamics of soil carbon and nitrogen stocks after afforestation in arid and semi-arid regions: A meta-analysis. Sci. Total Environ. 2018, 618, 1658–1664. [Google Scholar] [CrossRef]
- Guo, L.B.; Gifford, R.M. Soil carbon stocks and land use change: A meta analysis. Glob. Chang. Biol. 2002, 8, 345–360. [Google Scholar] [CrossRef]
- Cunningham, S.C.; Cavagnaro, T.R.; Mac Nally, R.; Paul, K.I.; Baker, P.J.; Beringer, J.; Thomson, J.R.; Thompson, R.M. Reforestation with native mixed-species plantings in a temperate continental climate effectively sequesters and stabilizes carbon within decades. Glob. Chang. Biol. 2015, 21, 1552–1566. [Google Scholar] [CrossRef]
- Guo, Y.; Abdalla, M.; Espenberg, M.; Hastings, A.; Hallett, P.; Smith, P. A systematic analysis and review of the impacts of afforestation on soil quality indicators as modified by climate zone, forest type and age. Sci. Total Environ. 2021, 757, 143824. [Google Scholar] [CrossRef]
- Deng, Q.; McMahon, D.E.; Xiang, Y.; Yu, C.L.; Jackson, R.B.; Hui, D. A global meta-analysis of soil phosphorus dynamics after afforestation. New Phytol. 2017, 213, 181–192. [Google Scholar] [CrossRef]
- Yang, S.; Zhao, W.; Pereira, P. Determinations of environmental factors on interactive soil properties across different land-use types on the Loess Plateau, China. Sci. Total Environ. 2020, 738, 140270. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable Type | Variable | Unit | Mean (SD) | Minimum | Maximum |
|---|---|---|---|---|---|
| Geographic | Elevation above sea level (Elevation) | m | 1237.0 (304.2) | 881.0 | 1939.0 |
| Distance to crops (D_crops) | m | 188.5 (308.0) | 0.0 | 1488.0 | |
| Distance to natural vegetation (D_n_veg) | m | 61.9 (133.7) | 0.0 | 531.5 | |
| Distance to shrub patches (D_shrub) | m | 1241.0 (1022.0) | 90.5 | 4675.0 | |
| Distance to Quercus woodland (D_Qil) | m | 1727.0 (2219.0) | 0.0 | 9834.0 | |
| Climatic | Annual Precipitation (Pp) | mm | 394.0 (65.2) | 311.0 | 547.6 |
| Mean Temperature (T) | °C | 12.68 (0.98) | 10.90 | 14.10 | |
| Edaphic | Available water (AW) | % | 6.82 (2.90) | 2.22 | 12.74 |
| Soil moisture at field capacity (33 kPa) (SM33) | % | 16.24 (5.78) | 5.09 | 28.58 | |
| Soil moisture of the wilting point (1500 kPa) (SM1500) | % | 9.42 (3.81) | 2.87 | 19.02 | |
| Water saturation (WS) | % | 38.39 (7.59) | 27.63 | 56.42 | |
| Gravel (>2 mm) | % | 37.24 (18.02) | 0.00 | 73.42 | |
| Sand (2–0.05 mm) | % | 45.28 (16.75) | 3.23 | 75.65 | |
| Coarse silt (0.05–0.02 mm) (CSilt) | % | 10.27 (4.35) | 3.07 | 24.67 | |
| Fine silt (0.02–0.002 mm) (FSilt) | % | 22.15 (9.66) | 8.37 | 57.25 | |
| Clay (<0.002 mm) | % | 22.30 (8.99) | 6.12 | 38.47 | |
| Cation exchange capacity (CEC) | Cmol(+)kg−1 | 11.28 (6.13) | 3.27 | 25.39 | |
| Electrical conductivity (EC) | mS cm−1 | 0.89 (0.40) | 0.51 | 2.21 | |
| Exchangeable sodium (Na) | Cmol(+)kg−1 | 0.06 (0.07) | 0 | 0.29 | |
| Exchangeable magnesium (Mg) | Cmol(+)kg−1 | 1.86 (1.28) | 0.30 | 5.98 | |
| Exchangeable potassium (K) | Cmol(+)kg−1 | 0.36 (0.27) | 0.06 | 1.28 | |
| Soluble calcium (s_Ca) | mg L−1 | 3.71 (2.41) | 0.67 | 9.81 | |
| Soluble magnesium (s_Mg) | mg L−1 | 0.65 (0.47) | 0.16 | 2.59 | |
| Soluble potassium (s_K) | mg L−1 | 0.40 (0.51) | 0.02 | 3.05 | |
| Soluble sodium (s_Na) | mg L−1 | 1.61 (0.98) | 0.13 | 4.78 | |
| CaCO3 (CaCO3) | % | 24.36 (23.24) | 0.10 | 73.78 | |
| pH | 7.90 (0.55) | 6.72 | 8.83 | ||
| Soil organic carbon (SOC) | % | 1.24 (0.82) | 0.30 | 3.19 | |
| Total nitrogen (TN) | % | 0.11 (0.05) | 0.05 | 0.24 | |
| NO2− (NO2) | mg L−1 | 3.29 (8.65) | 0.00 | 41.60 | |
| NO3− (NO3) | mg L−1 | 11.10 (44.41) | 0.00 | 233.40 | |
| SO42− (SO4) | mg L−1 | 46.60 (64.58) | 6.81 | 301.50 | |
| Cl− (Cl) | mg L−1 | 30.25 (22.01) | 10.53 | 117.2 | |
| Geological | Lithology | * | |||
| Land use | Age of afforestation | years | 8.4 (2.1) | 5.0 | 12.0 |
| Previous land use (cereal crop or old-field) | * | ||||
| Plant cover (Cover) | % | 67.49 (13.15) | 40.00 | 91.00 | |
| Cover of Pinus halepensis (CovPha) | % | 5.93 (6.44) | 0.00 | 32.00 | |
| Cover of Quercus ilex subsp. ballota (CovQil) | % | 0.26 (0.62) | 0.00 | 3.00 |
| Estimate | SE | z Value | Pr(>|z|) | |
|---|---|---|---|---|
| Intercept | 1.39000 | 0.73410 | 1.894 | 0.058 |
| D_n_veg | −0.00096 | 0.00037 | −2.566 | 0.010 |
| D_shrub | −0.00008 | 0.00004 | −2.091 | 0.037 |
| s_Mg | 0.26380 | 0.09270 | 2.846 | 0.004 |
| s_K | −0.30620 | 0.09801 | −3.124 | 0.002 |
| CSilt | −0.02948 | 0.00944 | −3.121 | 0.002 |
| SOC | −0.21200 | 0.07265 | −2.919 | 0.004 |
| pH | 0.22720 | 0.08642 | 2.629 | 0.009 |
| TN | 3.18600 | 1.17900 | 2.703 | 0.007 |
| Age | −0.06633 | 0.02660 | −2.494 | 0.013 |
| Previous land use Old-field | −0.35190 | 0.09792 | −3.593 | <0.001 |
| Cover | 0.01508 | 0.00289 | 5.219 | <0.001 |
| Factor | Estimate | SE | t Value | Pr(>|t|) |
|---|---|---|---|---|
| (Intercept) | −0.92220 | 0.91590 | −1.007 | 0.322 |
| D_n_veg | −0.00098 | 0.00039 | −2.531 | 0.017 |
| D_shrub | −0.00018 | 0.00004 | −4.285 | <0.001 |
| D_Qil | −0.00005 | 0.00003 | −1.969 | 0.058 |
| s_Mg | 0.50330 | 0.13530 | 3.720 | 0.001 |
| s_K | −0.26050 | 0.11160 | −2.335 | 0.026 |
| CSilt | −0.01716 | 0.01152 | −1.490 | 0.147 |
| CaCO3 | −0.00672 | 0.00261 | −2.576 | 0.015 |
| pH | 0.47630 | 0.11550 | 4.123 | <0.001 |
| Age | −0.06244 | 0.03068 | −2.036 | 0.051 |
| Old-field previous land use | −0.35910 | 0.10910 | −3.292 | 0.003 |
| CovQil | 0.13640 | 0.06451 | 2.115 | 0.043 |
| Cover | 0.01312 | 0.00405 | 3.241 | 0.003 |
| Paired Sites | Paired t-Test | |||||
|---|---|---|---|---|---|---|
| Cereal Crop | Afforestation | t | df | p-Value | ||
| Plant diversity | Abundance | 102.50 (18.21) | 97.33 (23.42) | 0.36183 | 5 | >0.05 |
| Total_R | 12.00 (3.03) | 21.00 (6.20) | −4.0249 | 5 | <0.05 | |
| Total_H | 1.50 (0.31) | 2.34 (0.63) | −4.9691 | 5 | <0.01 | |
| Edaphic properties 0–5 cm | Gravels (%) | 45.31 (15.61) | 46.12 (18.30) | −0.28349 | 17 | >0.05 |
| SM33 (%) | 19.89 (4.75) | 21.86 (4.84) | −2.4497 | 17 | <0.05 | |
| SM1500 (%) | 12.13 (2.87) | 13.75 (3.30) | −4.1787 | 17 | <0.001 | |
| AW (%) | 7.75 (2.88) | 8.11 (2.39) | −0.71374 | 17 | >0.05 | |
| BD (g cm−3) | 0.84 (0.11) | 0.79 (0.14) | 1.9476 | 17 | >0.05 | |
| pH | 8.55 (0.18) | 8.41 (0.27) | 2.4947 | 17 | >0.05 | |
| CaCO3 (%) | 38.84 (17.13) | 34.68 (15.44) | 1.1000 | 17 | >0.05 | |
| Mg (Cmol(+) kg−1) | 1.68 (0.68) | 1.75 (0.94) | −0.28188 | 17 | >0.05 | |
| K (Cmol(+) kg−1) | 0.41 (0.24) | 0.57 (0.23) | −4.1418 | 17 | <0.001 | |
| Na (Cmol(+) kg−1) | 0.33 (0.54) | 0.15 (0.20) | 1.2212 | 17 | >0.05 | |
| CEC (Cmol(+) kg−1) | 12.83 (5.22) | 14.96 (7.70) | −0.9949 | 17 | >0.05 | |
| P (mg kg−1) | 11.55 (6.81) | 15.70 (12.38) | −1.6987 | 17 | >0.05 | |
| SOC (%) | 1.93 (0.96) | 2.75 (1.13) | −3.5696 | 17 | <0.01 | |
| SOC (Mg C ha−1) | 7.91 (3.57) | 10.30 (3.56) | −2.9043 | 17 | <0.01 | |
| TN (%) | 0.14 (0.04) | 0.18 (0.05) | −4.539 | 17 | <0.001 | |
| C:N | 13.14 (4.70) | 15.24 (5.07) | −2.0917 | 17 | 0.0518 | |
| 5–10 cm | Gravels (%) | 44.92 (15.52) | 44.00 (17.95) | 0.45947 | 17 | >0.05 |
| SM33 (%) | 20.43 (4.51) | 20.40 (5.06) | 0.02957 | 17 | >0.05 | |
| SM1500 (%) | 12.38 (2.97) | 12.90 (3.17) | −1.2572 | 17 | >0.05 | |
| AW (%) | 8.05 (2.75) | 7.50 (2.49) | 1.0489 | 17 | >0.05 | |
| BD (g cm−3) | 0.88 (0.20) | 0.90 (0.17) | −0.90351 | 17 | >0.05 | |
| pH | 8.59 (0.23) | 8.54 (0.24) | 0.77857 | 17 | >0.05 | |
| CaCO3 (%) | 39.29 (17.09) | 33.80 (15.18) | 1.4698 | 17 | >0.05 | |
| Mg (Cmol(+) kg−1) | 1.89 (1.09) | 1.54 (0.98) | 1.0955 | 17 | >0.05 | |
| K (Cmol(+) kg−1) | 0.39 (0.24) | 0.39 (0.16) | 0.0191 | 17 | >0.05 | |
| Na (Cmol(+) kg−1) | 0.14 (0.15) | 0.33 (0.55) | 0.0191 | 17 | >0.05 | |
| CEC (Cmol(+) kg−1) | 12.31 (6.11) | 13.03 (6.49) | −0.4215 | 17 | >0.05 | |
| P (mg kg−1) | 10.88 (6.59) | 7.19 (5.26) | 2.9986 | 17 | <0.01 | |
| SOC (%) | 1.77 (0.85) | 1.91 (0.91) | −0.74067 | 17 | >0.05 | |
| SOC (Mg C ha−1) | 7.42 (3.12) | 7.95 (2.99) | −0.06191 | 17 | >0.05 | |
| TN (%) | 0.14 (0.04) | 0.15 (0.05) | −0.92473 | 17 | >0.05 | |
| C:N | 12.47 (5.47) | 12.82 (4.93) | −0.28237 | 17 | >0.05 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segura, C.; Jiménez, M.N.; Fernández-Ondoño, E.; Navarro, F.B. Effects of Afforestation on Plant Diversity and Soil Quality in Semiarid SE Spain. Forests 2021, 12, 1730. https://doi.org/10.3390/f12121730
Segura C, Jiménez MN, Fernández-Ondoño E, Navarro FB. Effects of Afforestation on Plant Diversity and Soil Quality in Semiarid SE Spain. Forests. 2021; 12(12):1730. https://doi.org/10.3390/f12121730
Chicago/Turabian StyleSegura, Carmen, María N. Jiménez, Emilia Fernández-Ondoño, and Francisco B. Navarro. 2021. "Effects of Afforestation on Plant Diversity and Soil Quality in Semiarid SE Spain" Forests 12, no. 12: 1730. https://doi.org/10.3390/f12121730
APA StyleSegura, C., Jiménez, M. N., Fernández-Ondoño, E., & Navarro, F. B. (2021). Effects of Afforestation on Plant Diversity and Soil Quality in Semiarid SE Spain. Forests, 12(12), 1730. https://doi.org/10.3390/f12121730