
Trichoderma spp. from Pine Bark and Pine Bark Extracts: Potent Biocontrol Agents against Botryosphaeriaceae

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Pinus Sylvestris Bark Extracts
2.2. Fungal Isolates
2.3. In Vitro Assessment of Antifungal Activity of Pinus Sylvestris Bark Extracts
2.4. In Vitro Assay of Trichoderma spp. Antifungal Activity
2.5. Biochemical Characterization of Trichoderma spp. Antifungal Activity
2.6. The Effect of Trichoderma VOCs on Mycelial Growth of Botryosphaeriaceae
2.7. Collection of VOCs and GC−MS Analysis
2.8. Statistical Analyses
3. Results
3.1. In Vitro Assessment of Antifungal Activity of Pinus Sylvestris Bark Extracts
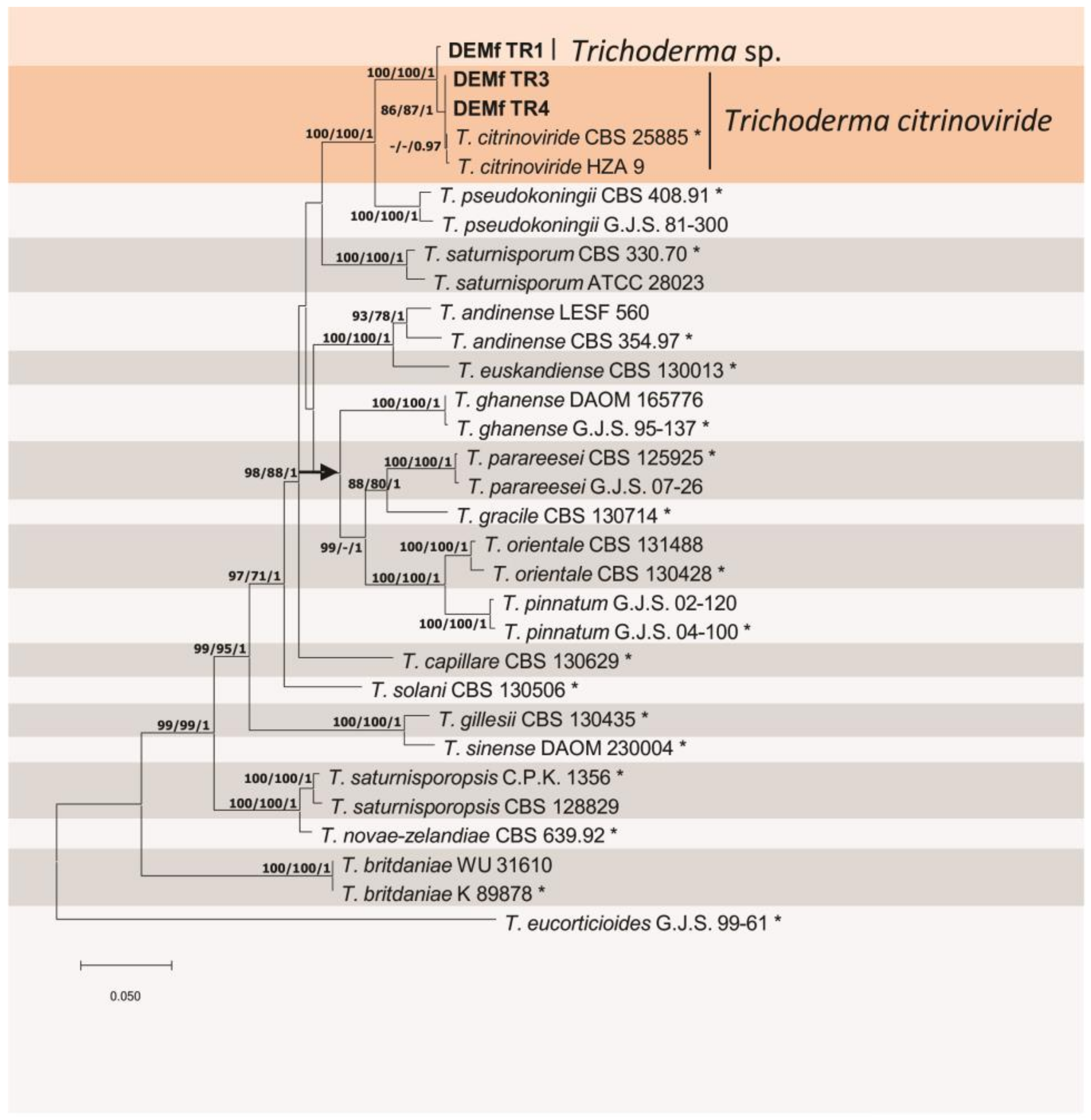
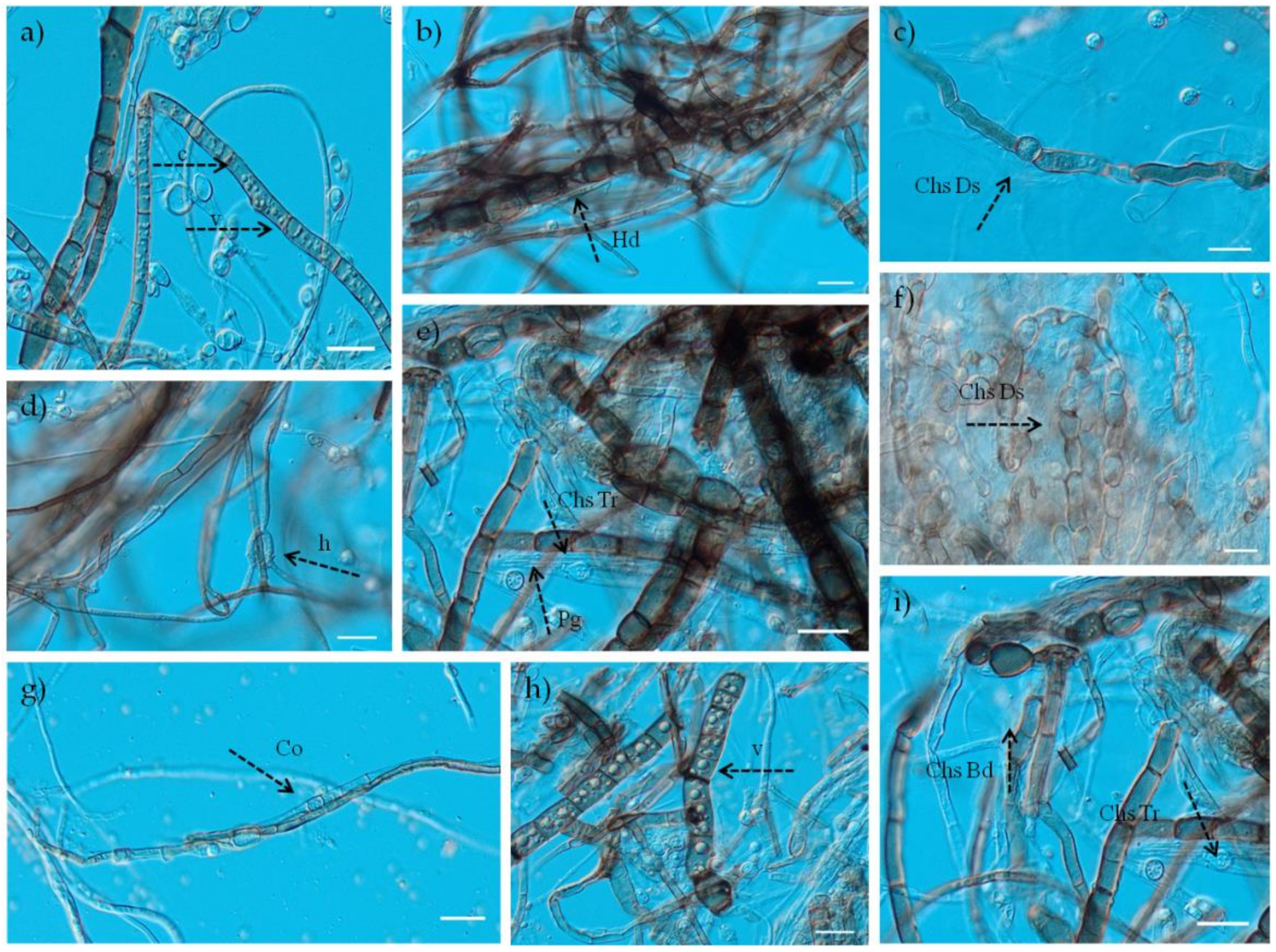
3.2. Identification of Trichoderma spp.
3.3. In Vitro Assay of Trichoderma spp. Antifungal Activity
3.4. Biochemical Characterization of Trichoderma spp. Antifungal Activity
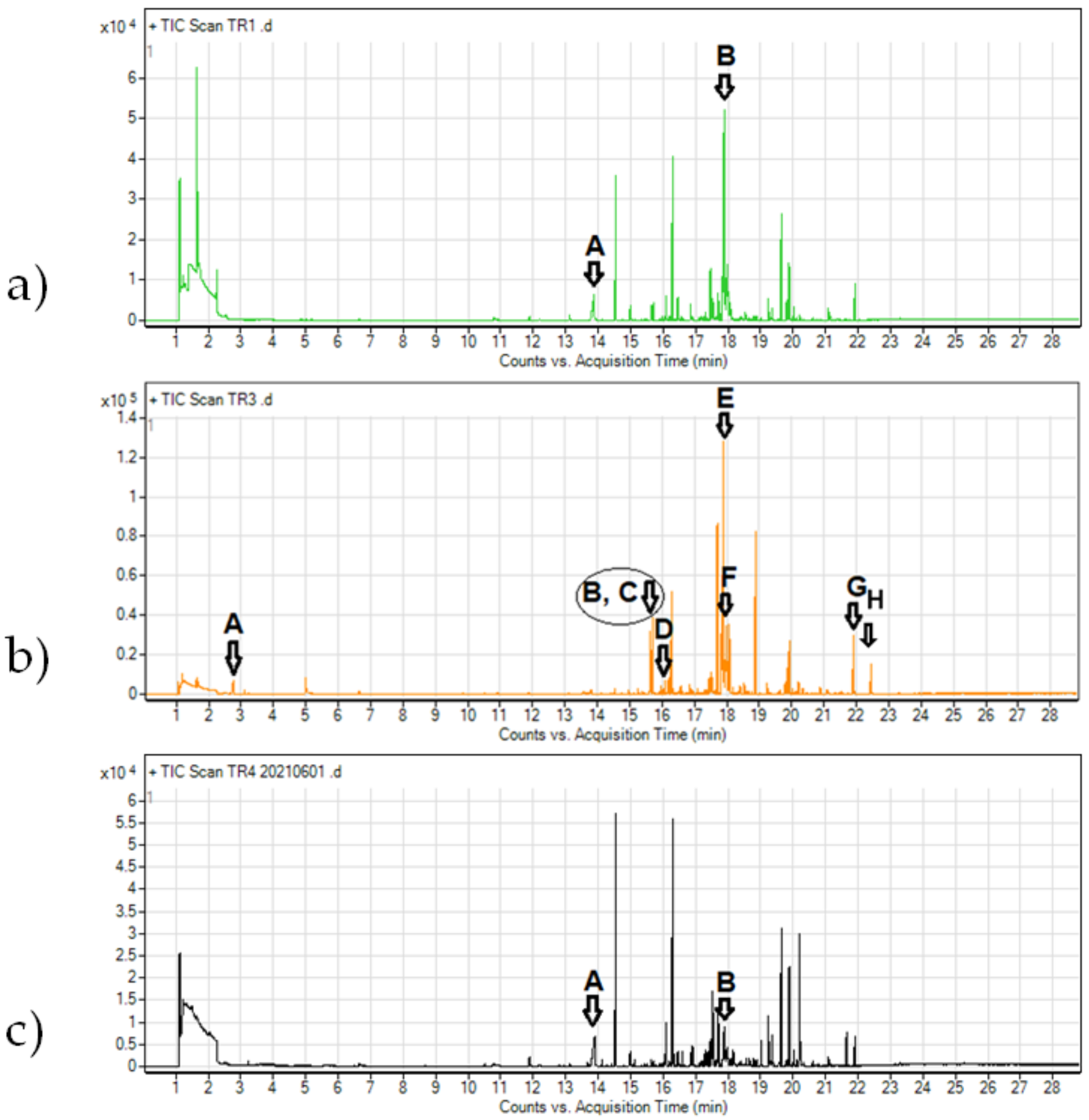
3.5. Effect of Trichoderma VOCs on Mycelial Growth of Botryosphaeriaceae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Elsherbiny, E.A.; Amin, B.H.; Aleem, B.; Kingsley, K.L.; Bennett, K.K. Trichoderma volatile organic compounds as a biofumigation tool against late blight pathogen Phytophthora infestans in post harvest potato tubers. J. Agric. Food Chem. 2020, 68, 8163–8171. [Google Scholar] [CrossRef] [PubMed]
- Aarti, T.; Meenu, S. Role of volatile metabolites from T. citrinoviride in biocontrol of phytopathogens. Int. J. Res. Chem. Environ. 2015, 5, 86–95. [Google Scholar]
- Pásztory, Z.; Mohácsiné, I.R.; Gorbacheva, G.; Börcsök, Z. The utilization of tree bark. BioResources 2016, 11, 7859–7888. [Google Scholar] [CrossRef]
- Bianchi, S. Extraction and Characterization of Bark Tannins from Domestic Softwood Species. Ph.D. Thesis, University of Hamburg, Hamburg, Germany, 13 January 2017. [Google Scholar]
- Fregoso-Madueño, J.N.; Goche-Télles, J.R.; Rutiaga-Quiñones, J.G.; González-Laredo, R.F.; Bocanegra-Salazar, M.; Chávez-Simental, J.A. Alternative uses of sawmill industry waste. Rev. Chapingo Ser. Cienc For. Ambient 2017, 23, 243–260. [Google Scholar] [CrossRef]
- Minova, S.; Seškēna, R.; Voitkâne, S.; Metla, Z.; Daugavietis, M.; Jankevica, L. Impact of pine (Pinus sylvestris L.) and spruce (Picea abies (L.) Karst.) bark extracts on important strawberry pathogens. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2015, 69, 62–67. [Google Scholar] [CrossRef]
- Hagge, J.; Bässler, C.; Gruppe, A.; Hoppe, B.; Kellner, H.; Krah, F.-S.; Müller, J.; Seibold, S.; Stengel, E.; Thorn, S. Bark coverage shifts assembly processesof microbial decomposer communitiesin dead wood. Proc. R. Soc. B 2019, 286, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zlatković, M.; Keča, N.; Wingfield, M.J.; Jami, F.; Slippers, B. Shot hole disease on Prunus laurocerasus caused by Neofusicoccum parvum in Serbia. For. Pathol. 2016, 46, 666–669. [Google Scholar] [CrossRef]
- Zlatković, M.; Keča, N.; Wingfield, M.J.; Jami, F.; Slippers, B. Botryosphaeriaceae associated with the die-back of ornamental trees in the Western Balkans. Antonie Leeuwenhoek 2016, 109, 543–564. [Google Scholar] [CrossRef]
- Zlatković, M. Botryosphaeriaceae species associated with canker and die-back disease of conifers in urban environments in Serbia. Topola/Poplar. Inst. Lowl. For. Univ. Novi Sad. 2017, 199, 55–75. (In Serbian) [Google Scholar]
- Popović, T.; Blagojević, J.; Aleksić, G.; Jelušić, A.; Krnjajić, S.; Milovanović, P. A blight disease on highbush blueberry associated with Macrophomina phaseolina in Serbia. Can. J. Plant Pathol. 2018, 40, 121–127. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fung. Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Zlatković, M.; Wingfield, M.J.; Jami, F.; Slippers, B. Host specificity of co-infecting Botryosphaeriaceae on ornamental and forest trees in the Western Balkans. For. Pathol. 2018, 48, e12410. [Google Scholar] [CrossRef]
- Zlatković, M.; Wingfield, M.J.; Jami, F.; Slippers, B. Genetic uniformity characterizes the invasive spread of Neofusicoccum parvum and Diplodia sapinea in the Western Balkans. For. Pathol. 2019, 49, e12491. [Google Scholar] [CrossRef]
- Moya, P.; Girotti, J.R.; Toledo, A.V.; Sisterna, M.N. Antifungal activity of Trichoderma VOCs against Pyrenophora teres, the causal agent of barley net blotch. J. Plant Prot. Res. 2018, 58, 45–53. [Google Scholar] [CrossRef]
- Shah, M.M.; Afiya, H. Introductory Chapter: Identification and Isolation of Trichoderma spp.—Their Significance in Agriculture, Human Health, Industrial and Environmental Application. Trichoderma—The Most Widely Used Fungicide; Shah, M.M., Sharif, U., Buhari, T.R., Eds.; IntechOpen: London, UK, 2019; pp. 1–13. [Google Scholar] [CrossRef]
- Stracquadanio, C.; Quiles, J.M.; Meca, G.; Cacciola, S.O. Antifungal activity of bioactive metabolites produced by Trichoderma asperellum and Trichoderma atroviride in liquid medium. J. Fungi 2020, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ghirardo, A.; Weber, B.; Schnitzler, J.-P.; Benz, J.P.; Rosenkranz, M. Trichoderma species differ in their volatile profiles and in antagonism toward ectomycorrhiza Laccaria bicolor. Front. Microbiol. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Zhang, F.; Yang, X.; Ran, W.; Shen, Q. Fusarium oxysporum induces the production of proteins and volatile organic compounds by Trichoderma harzianum T-E5. FEMS Microbiol. Lett. 2014, 359, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Özgenç, Ö.; Durmaz, S.; Yildiz, Ü.C.; Erişir, E. A comparison between some wood bark extracts: Antifungal activity. Kastamonu Univ. Orman. Derg. 2017, 17, 502–508. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Tomaselli, E.; Pollard-Flamand, J.; Boulé, J.; Gerin, D.; Pollastro, S. Characterization of Trichoderma isolates from southern Italy, and their potential biocontrol activity against grapevine trunk disease fungi. Phytopathol. Mediterr. 2020, 59, 425–439. [Google Scholar] [CrossRef]
- Karličić, V.; Jovičić-Petrović, J.; Marojević, V.; Zlatković, M.; Orlović, S.; Raičević, V. Potential of Trichoderma spp. and Pinus sylvestris bark extracts as biocontrol agents against fungal pathogens residing in the Botryosphaeriales. Environ. Sci. Proc. 2021, 3, 99. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.A.; Gelfand, D.H.; Sninsky, J.J.; White, T.J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evolut. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Kovač, M.; Diminić, D.; Orlović, S.; Zlatković, M. Botryosphaeria dothidea and Neofusicoccum yunnanense causing canker and die-back of Sequoiadendron giganteum in Croatia. Forests 2021, 12, 695. [Google Scholar] [CrossRef]
- Zlatković, M.; Tenorio-Baigorria, I.; Lakatos, T.; Tóth, T.; Koltay, A.; Pap, P.; Marković, M.; Orlović, S. Bacterial canker disease on Populus x euramericana caused by Lonsdalea populi in Serbia. Forests 2020, 11, 1080. [Google Scholar] [CrossRef]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef]
- Romsaiyud, A.; Singkasiri, W.; Nopharatana, A.; Chaiprasert, P. Combination effect of pH and acetate on enzymatic cellulose hydrolysis. J. Environ. Sci. 2009, 21, 965–970. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fert. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Dennis, C.; Webster, J. Antagonistic properties of species groups of Trichoderma-II. Production of volatile antibiotics. Trans. Br. Mycol Soc. 1971, 57, 47–48. [Google Scholar] [CrossRef]
- Mohareb, A.S.; Kherallah, I.E.A.; Badawy, M.E.; Salem, M.Z.M.; Yousef, H.A. Chemical composition and activity of bark and leaf extracts of Pinus halepensis and Olea europaea grown in AL-Jabel AL-Akhdar region, Libya against some plant phytopathogens. J. Appl. Biotechnol. Bioeng. 2017, 3, 331–342. [Google Scholar] [CrossRef]
- Ruiz-Gómez, F.J.; Miguel-Rojas, C. Antagonistic potential of native Trichoderma spp. against Phytophthora cinnamomiin the control of Holm Oak decline in Dehesas ecosystems. Forests 2021, 12, 945. [Google Scholar] [CrossRef]
- Samuels, G.; Petrini, O.; Kuhls, K.; Lieckfeldt, E.; Kubicek, C.P. The Hypocrea schweinitzii complex and Trichoderma sect. Longibrachiatum. Stud. Mycol. 1998, 41, 1–54. [Google Scholar]
- Krause, K.; Jung, E.-M.; Lindner, J.; Hardiman, I.; Poetschner, J.; Madhavan, S.; Mtthäus, C.; Kai, M.; Menezes, R.C.; Popp, J.; et al. Responseof the wood-decay fungus Schizophyllum commune to co-occurring microorganisms. PLoS ONE 2020, 15, e0232145. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Aoyama, M.; Doi, S.; Kanetoshi, A.; Hayashi, T. Antifungal activity of bark extracts of conifers. Holz Roh Werkst 1995, 53, 81–82. [Google Scholar] [CrossRef]
- Alfredsen, G.; Solheim, H.; Slimestad, R. Antifungal effect of bark extracts from some European tree species. Eur. J. Forest Res. 2008, 127, 387–393. [Google Scholar] [CrossRef]
- Vek, V.; Poljanšek, I.; Humar, M.; Willför, S.; Oven, P. In vitro inhibition of extractives from knotwood of P. sylvestris (Pinus sylvestris) and black pine (Pinus nigra) on growth of Schizophyllum commune, Trametes versicolor, Gloeophyllum trabeum and Fibroporia vaillantii. Wood Sci. Technol. 2020, 54, 1645–1662. [Google Scholar] [CrossRef]
- Lomeli-Ramírez, M.G.; Dávila-Soto, H.; Silva-Guzmán, J.A.; Ruiz, H.G.O.; García-Enriquez, S. Fungitoxic potential of extracts of four Pinus spp. bark to inhibit fungus Trametes versicolor (L.ex. Fr.) Pilát. BioResources 2016, 11, 10575–10584. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [PubMed]
- Marraschi, R.; Ferreira, A.B.M.; da Silva Bueno, R.N.; Leite, J.A.B.P.; Lucon, C.M.M.; Harakava, R.; Leite, L.G.; Padovani, C.R.; Bueno, C.J. A protocol for selection of Trichoderma spp. to protect grapevine pruning wounds against Lasiodiplodia theobromae. Braz. J. Microbiol. 2019, 50, 213–221. [Google Scholar] [CrossRef]
- Park, Y.-H.; Mishra, R.C.; Yoon, S.; Kim, H.; Park, C.; Seo, S.-T.; Bae, H. Endophytic Trichoderma citrinoviride isolated from mountain-cultivated ginseng (Panax ginseng) has great potential as a biocontrol agent against ginseng pathogens. J. Ginseng Res. 2019, 43, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Kuzmanovska, B.; Rusevski, R.; Jankulovska, M.; Oreshkovikj, K.B. Antagonistic activity of Trichoderma asperellum and Trichoderma harzianum against genetically diverse Botrytis cinerea isolates. Chil. J. Agric. Res. 2018, 78, 391–399. [Google Scholar] [CrossRef]
- Kotze, C.; Van Niekerk, J.; Mostert, L.; Halleen, F.; Fourie, P. Evaluation of biocontrol agents for grapevine pruning wound protection against trunk pathogen infection. Phytopathol. Mediterr. 2011, 50, S247–S263. [Google Scholar]
- Pellan, L.; Durand, N.; Martinez, V.; Fontana, A.; Schorr-Galindo, S.; Strub, C. Commercial biocontrol agents reveal contrasting comportments against two mycotoxigenic fungi in cereals: Fusarium graminearum and Fusarium verticillioides. Toxins 2020, 12, 152. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, D.K.; Dubey, R.C.; Sharma, V.K. Biocontrol effects of Trichoderma virens on Macrophomina phaseolina causing charcoal rot of peanut. Ind. J. Microbiol. 2001, 41, 251–256. [Google Scholar]
- Nidhina, K.; Sharadraj, K.M.; Prathibha, V.H.; Hegde, V.; Gangaraj, K.P. Antagonistic activity of Trichoderma spp. to Phytophthora infecting plantation crops and its beneficial effect on germination and plant growth promotion. Vegetos 2016, 29, 19–26. [Google Scholar] [CrossRef]
- Danilović, G.; Radić, D.; Raičević, V.; Jovanović, L.J.; Kredics, L.; Panković, D. Extracellular enzyme activity of Trichoderma strains isolated from different soil types. In Proceedings of the 2nd International Symposium for Agriculture and Food, Ohrid, North Macedonia, 7–9 October 2015. [Google Scholar]
- Lorito, M.; Hayes, C.K.; Di Pietro, A.; Woo, S.L.; Harman, G.E. Prification, characterization, andsynergistic activity of glucan 1,3-β-glucosidse and an N-cetyl-β-glucosaminidase from Trichoderma harzianum. Mol. Plant Pathol. 1994, 84, 398–405. [Google Scholar]
- Chen, D.; Hou, Q.; Jia, L.; Sun, K. Combined use of two Trichoderma strains to promote growth of pakchoi (Brassica chinensis L.). Agronomy 2021, 11, 726. [Google Scholar] [CrossRef]
- Chen, J.-L.; Liu, K.; Miao, C.-P.; Sun, S.-Z.; Chen, Y.-W.; Xu, L.-H.; Guan, H.-L.; Zhao, L.-X. Salt tolerance of endophytic Trichoderma koningiopsis YIM PH30002 and its volatile organic compounds (VOCs) allelopathicactivity against phytopathogens associated with Panax notoginseng. Ann. Microbiol. 2016, 66, 981–990. [Google Scholar] [CrossRef]
- Aneja, M.; Gianfagna, T.J.; Hebbar, P.K. Trichoderma harzianum produces nonanoic acid, an inhibitor of spore germination and mycelial growth of two cacao pathogens. Physiol. Mol. Plant Pathol. 2005, 67, 304–307. [Google Scholar] [CrossRef]
- Shareef, H.K.; Muhammed, H.J.; Hussein, H.M.; Hameed, I.H. Antibacterial effect of ginger (Zingiber officinale) roscoe and bioactive chemical analysis using Gas chromatography mass spectrum. Orient. J. Chem. 2016, 32, 817–837. [Google Scholar] [CrossRef]
- Kim, J.-E.; Seo, J.-H.; Bae, M.-S.; Bae, C.-S.; Yoo, J.-C.; Bang, M.-A.; Cho, S.-S.; Park, D.-H. Antimicrobial constituents from Allium hookeri Root. Nat. Prod. Commun. 2016, 11, 237–238. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pine Bark Water Extracts | Water pH7/80 °C | Water pH9/80 °C | Water pH7/120 °C | Water pH9/120 °C | |
|---|---|---|---|---|---|
| B. dothidea CMW 39314 | 20% | 39 bA | 36 abA | 35 aA | 34 aA |
| 30% | 44 aB | 43 aB | 39 aB | 40 aB | |
| D. sarmentorum CMW 39365 | 20% | 56 aA | 56 aA | 48 aA | 60 aA |
| 30% | 60 abA | 62 abB | 53 aA | 66 bA | |
| N. parvum BOT 275 | 20% | NI | NI | NI | NI |
| 30% | NI | NI | NI | NI | |
| Pathogen | Trichoderma sp. DEMf TR 1 | T. citrinoviride DEMf TR3 | T. citrinoviride DEMf TR4 |
|---|---|---|---|
| B. dothidea CMW 39314 | 76 ab | 67 a | 85 b |
| D. sarmentorum CMW 39365 | 75 b | 63 a | 75 b |
| N. parvum BOT 275 | 59 b | 55 a | 62 c |
| Strains | Cell-Wall Degrading Enzymes | Sid | ||||
|---|---|---|---|---|---|---|
| Lipase | Est-Lip | Na-β | β-Glu | Cell | ||
| Trichoderma sp. DEMf TR 1 | 2 | 2 | 3 | 2 | + | + |
| T. citrinoviride DEMf TR3 | 2 | 2 | 3 | 3 | + | + |
| T. citrinoviride DEMf TR4 | 2 | 2 | 3 | 1 | + | + |
| Isolate | Retention Time(min) | Peak | Volatile Compound | Relative Abundance |
|---|---|---|---|---|
| Trichoderma sp. DEMf TR 1 | 13.8756 | A | Nonanoic acid | 4.84 |
| 17.8761 | B | Trichoacorenol | 17.68 | |
| T. citrinoviride DEMf TR3 | 2.7580 | A | Acetic acid | 1.19 |
| 15.6375 | B | Cubenene | 3.75 | |
| 15.7034 | C | Cubenene | 4.54 | |
| 16.2008 | D | cis-α-bergamotene | 0.93 | |
| 17.8873 | E | Trichoacorenol | 18.88 | |
| 17.9652 | F | Trichoacorenol | 1.54 | |
| 21.9053 | G | Verticillol | 3.43 | |
| 22.4483 | H | Hexanedioic acid | 1.64 | |
| T. citrinoviride DEMf TR4 | 13.9 | A | Nonanoic acid | 5.38 |
| 17.8773 | B | Trichoacorenol | 3.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karličić, V.; Zlatković, M.; Jovičić-Petrović, J.; Nikolić, M.P.; Orlović, S.; Raičević, V. Trichoderma spp. from Pine Bark and Pine Bark Extracts: Potent Biocontrol Agents against Botryosphaeriaceae. Forests 2021, 12, 1731. https://doi.org/10.3390/f12121731
Karličić V, Zlatković M, Jovičić-Petrović J, Nikolić MP, Orlović S, Raičević V. Trichoderma spp. from Pine Bark and Pine Bark Extracts: Potent Biocontrol Agents against Botryosphaeriaceae. Forests. 2021; 12(12):1731. https://doi.org/10.3390/f12121731
Chicago/Turabian StyleKarličić, Vera, Milica Zlatković, Jelena Jovičić-Petrović, Milan P. Nikolić, Saša Orlović, and Vera Raičević. 2021. "Trichoderma spp. from Pine Bark and Pine Bark Extracts: Potent Biocontrol Agents against Botryosphaeriaceae" Forests 12, no. 12: 1731. https://doi.org/10.3390/f12121731
APA StyleKarličić, V., Zlatković, M., Jovičić-Petrović, J., Nikolić, M. P., Orlović, S., & Raičević, V. (2021). Trichoderma spp. from Pine Bark and Pine Bark Extracts: Potent Biocontrol Agents against Botryosphaeriaceae. Forests, 12(12), 1731. https://doi.org/10.3390/f12121731