
Reproductive Biology of Olive Trees (Arbequina cultivar) at the Northern Limit of Their Distribution Areas

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
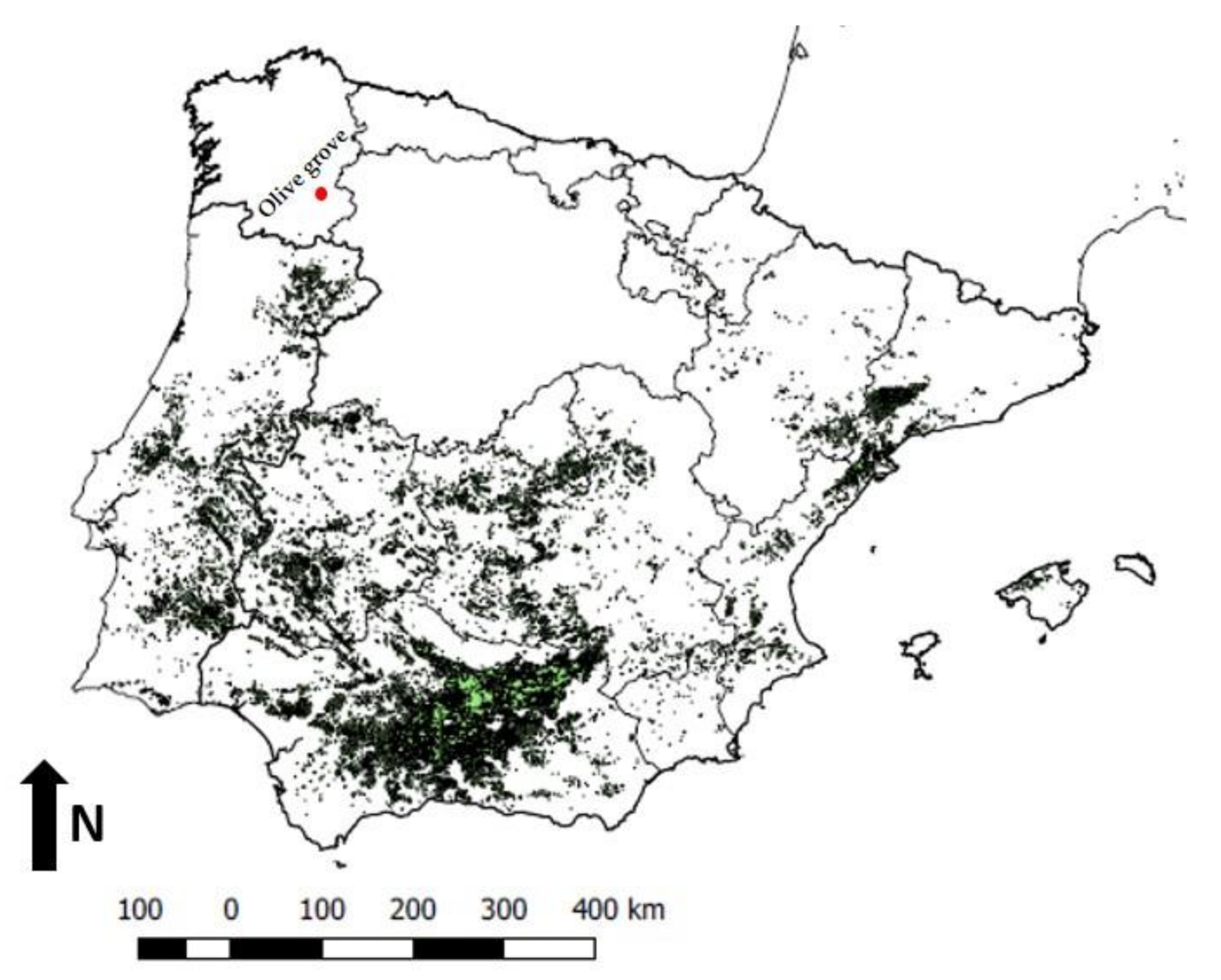
2. Materials and Methods
- N: number of chilling hours;
- B: 24D;
- D: length of the period in days;
- maxT: maximum temperature of the period;
- minT: minimum temperature of the period.
- GDD °C: °C accumulation;
- maxT: maximum diary temperature;
- Th: threshold temperature.
- avgT: daily mean temperature;
- Th: threshold temperature.
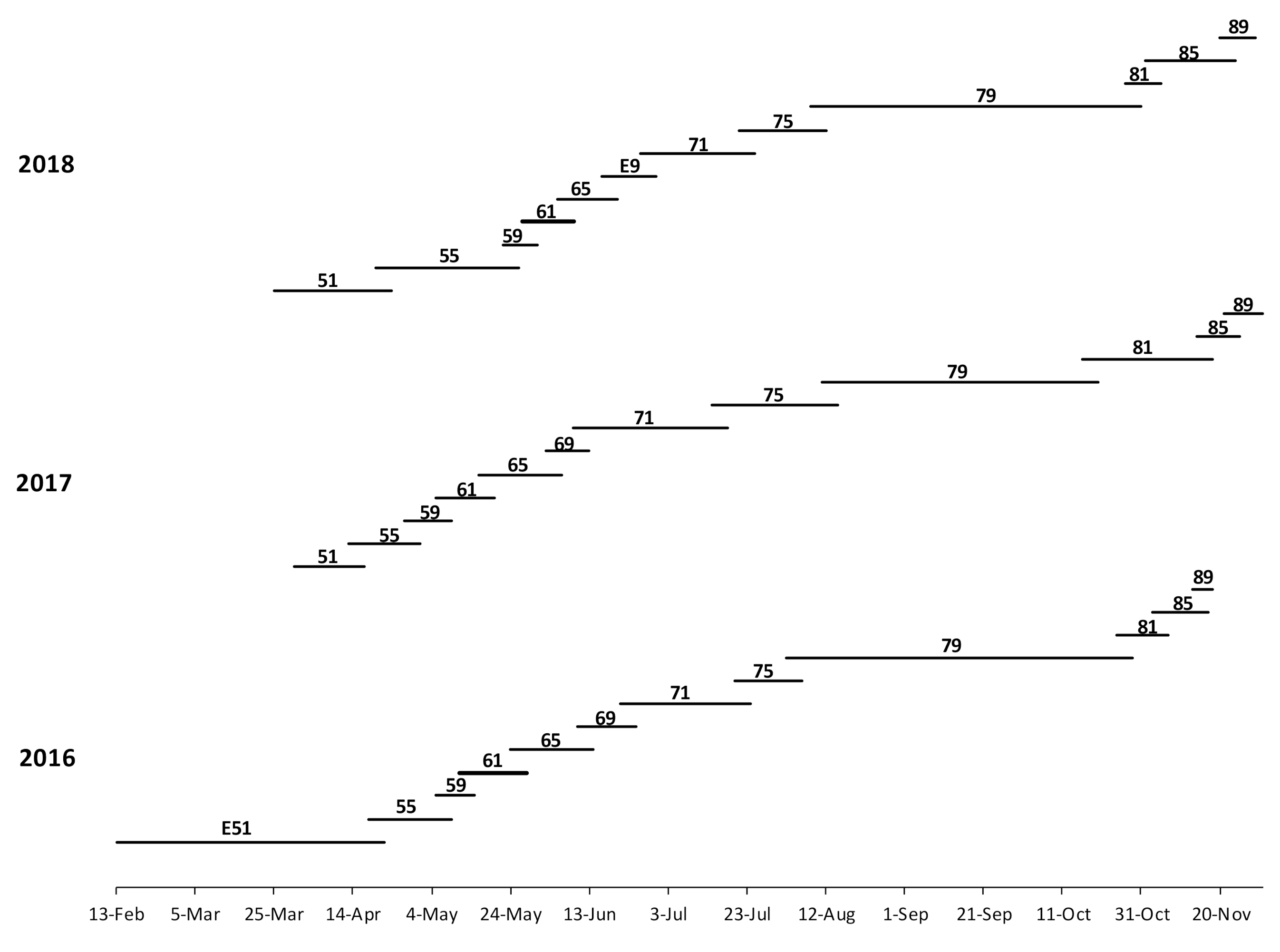
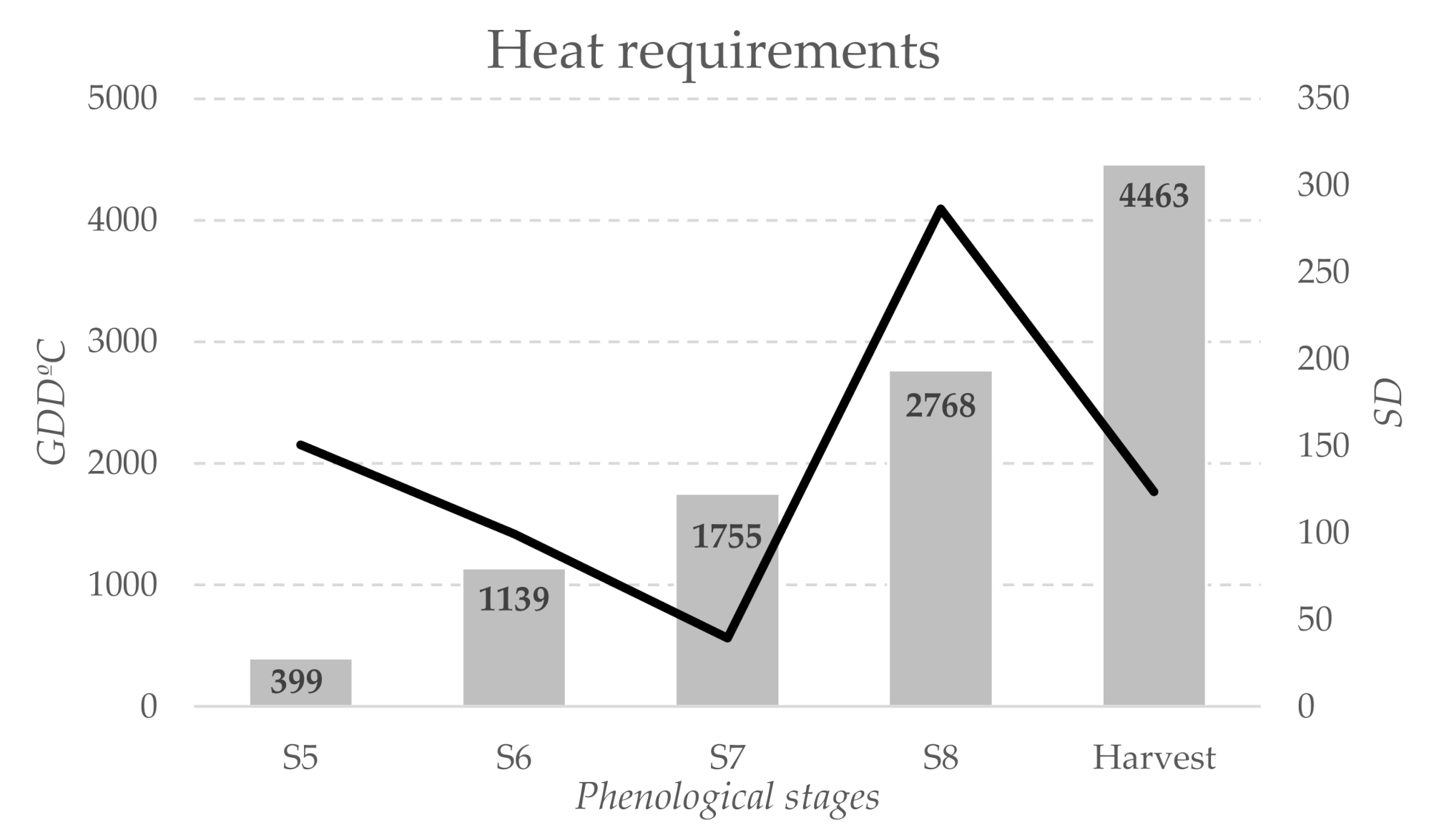
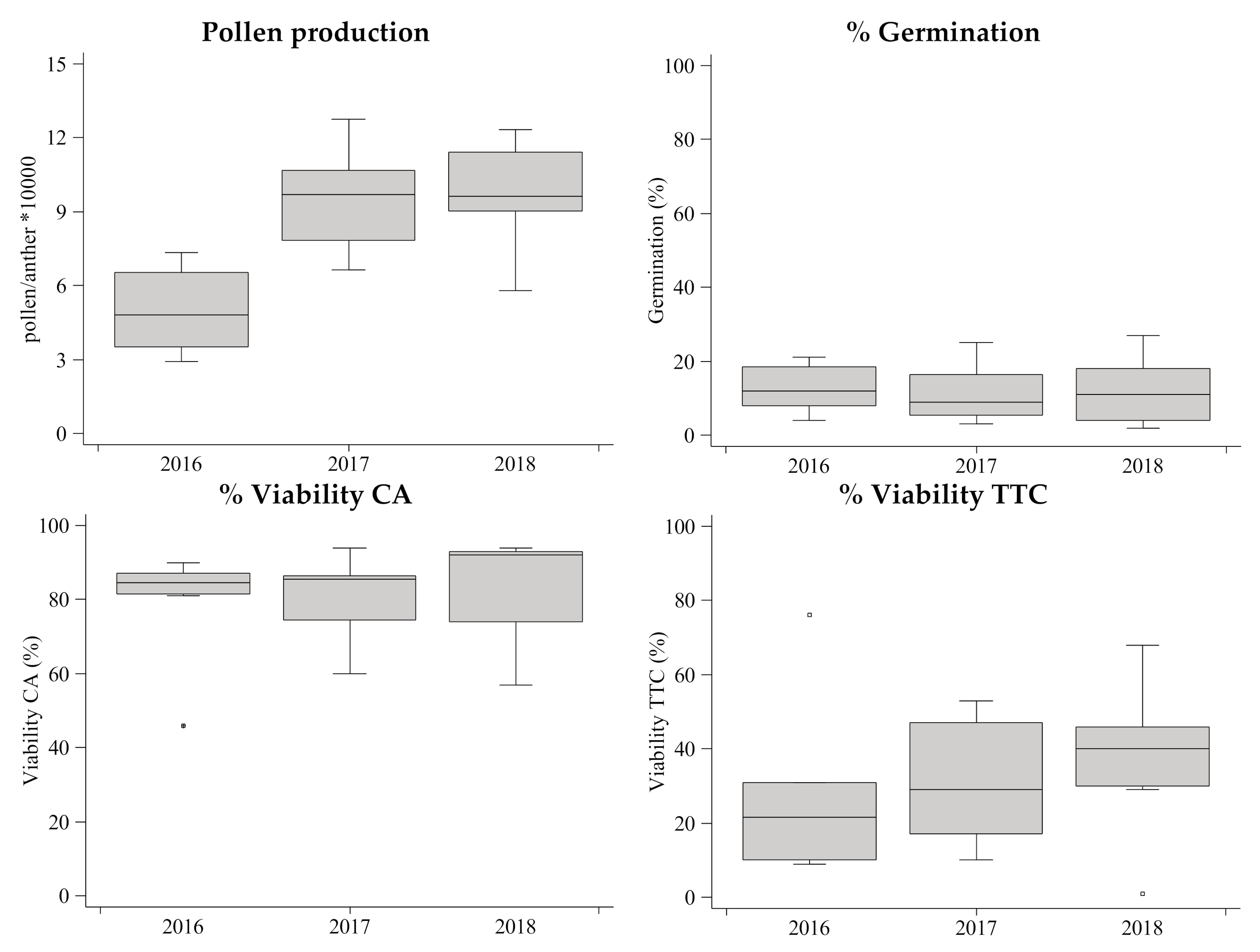
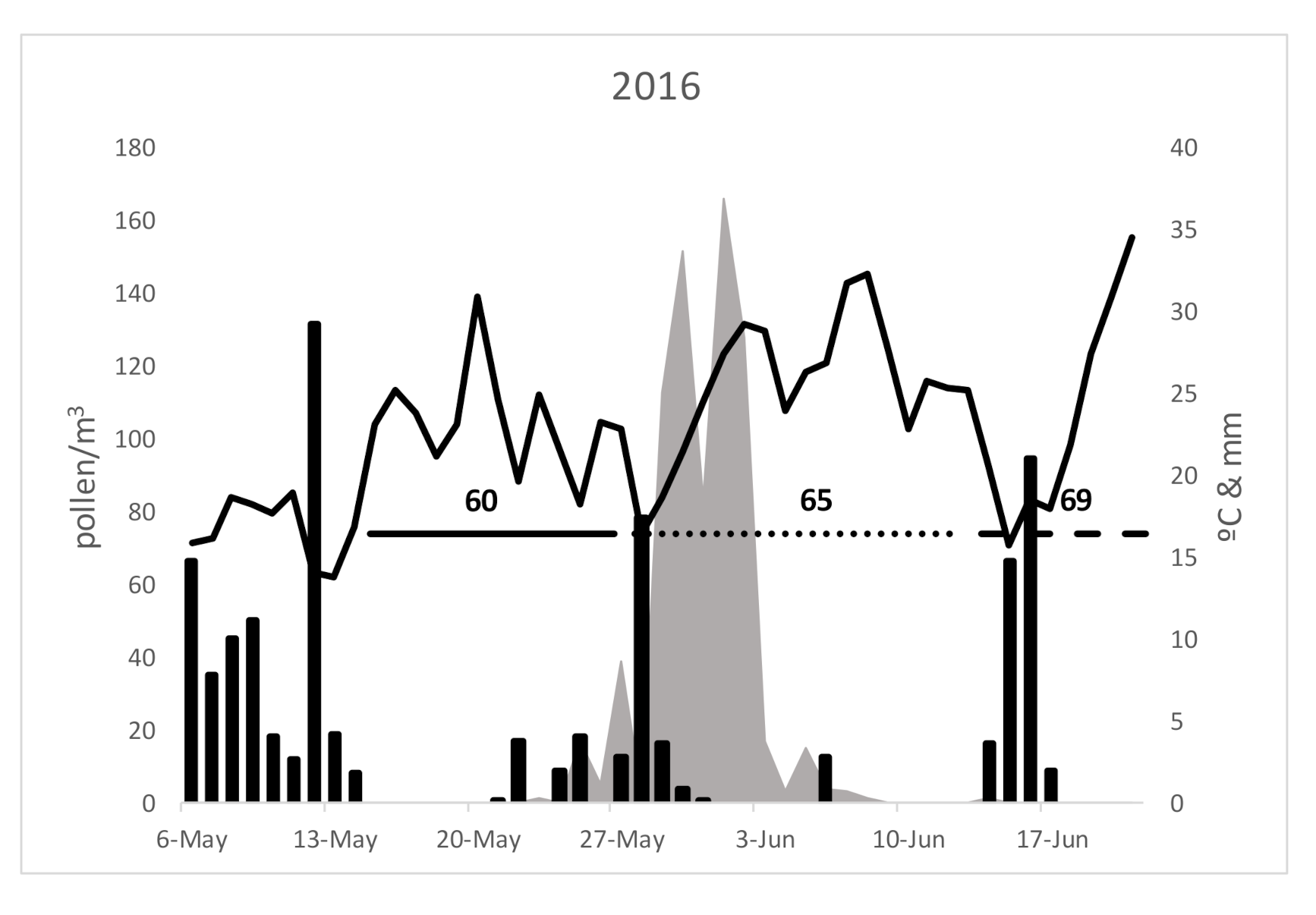
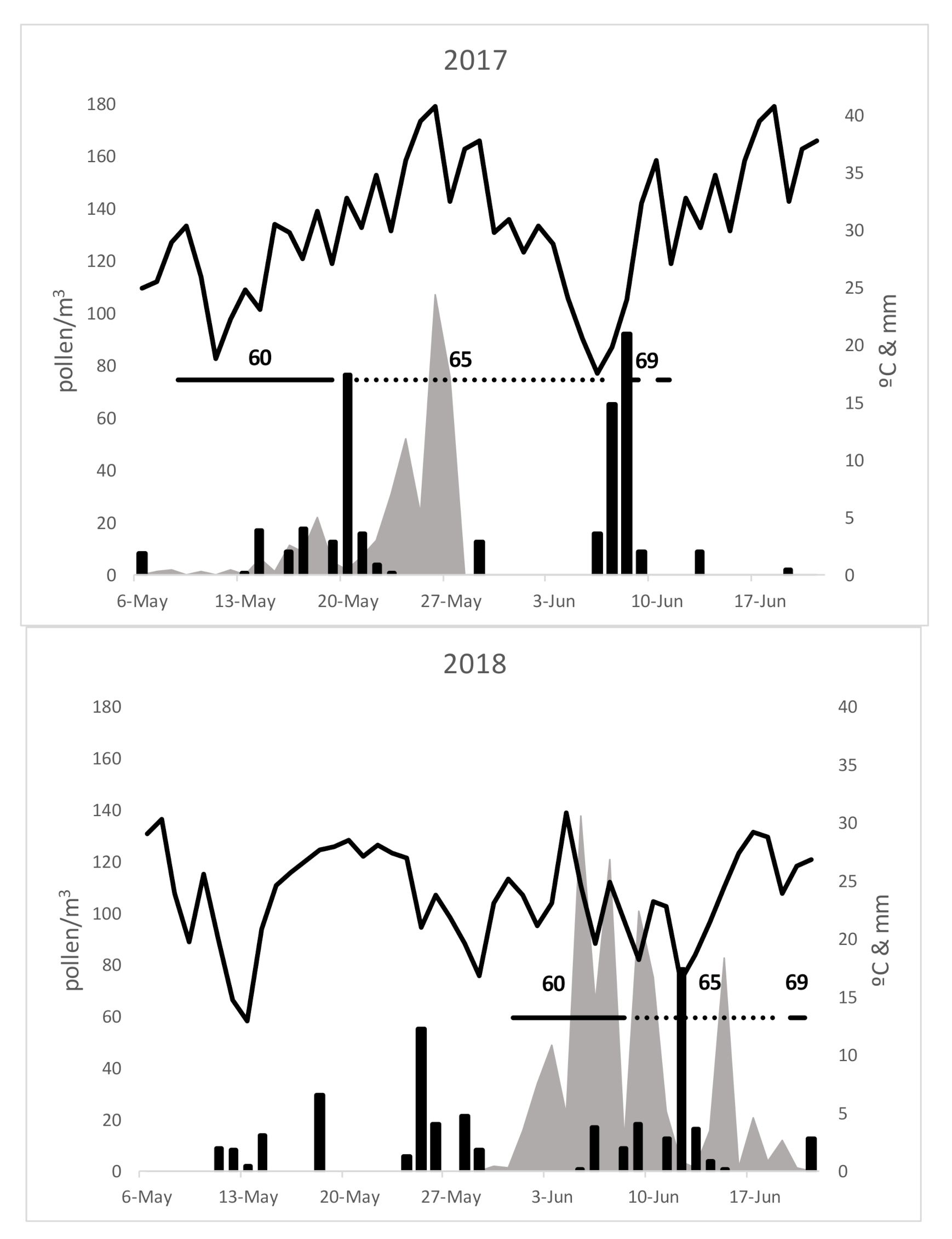
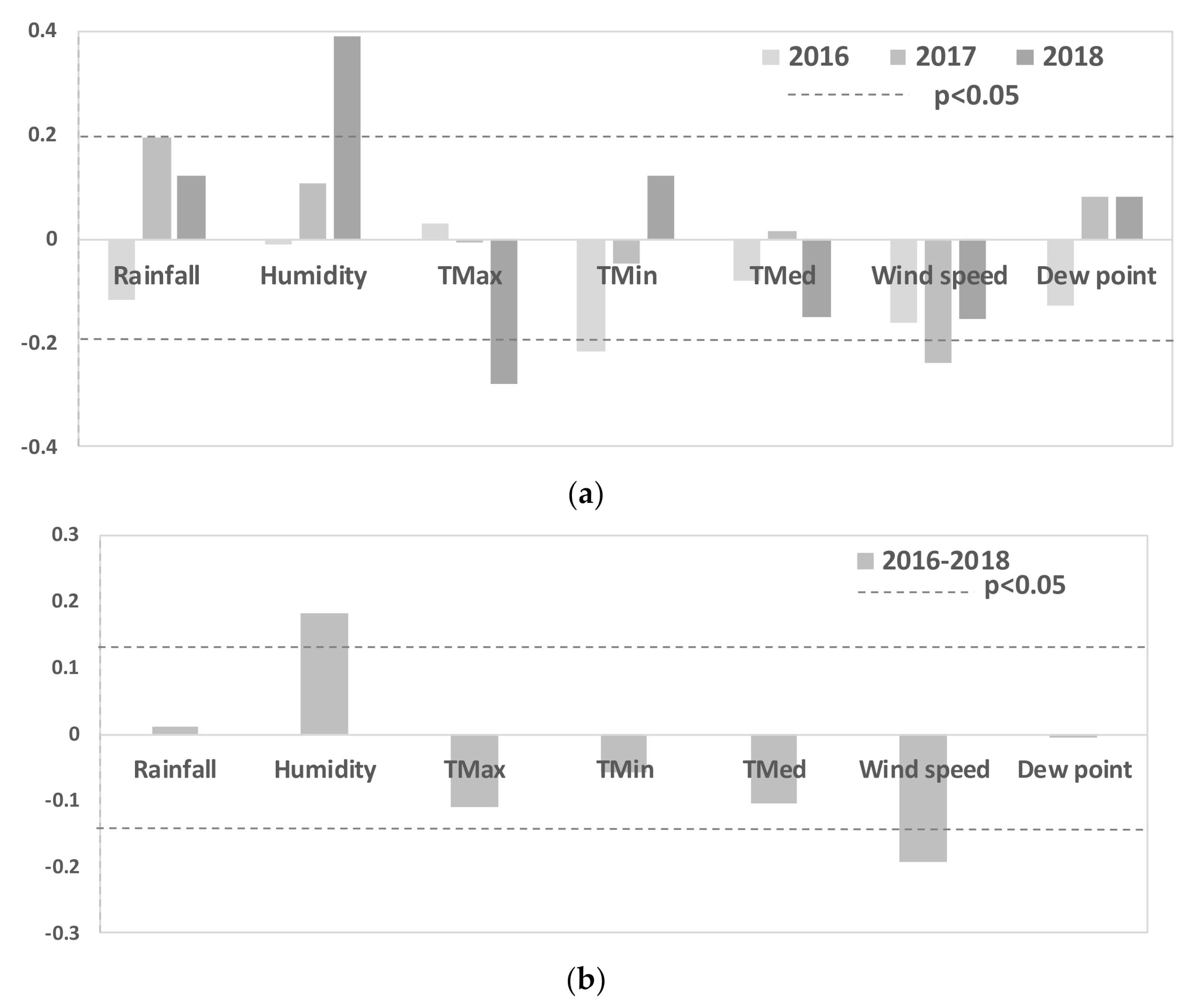
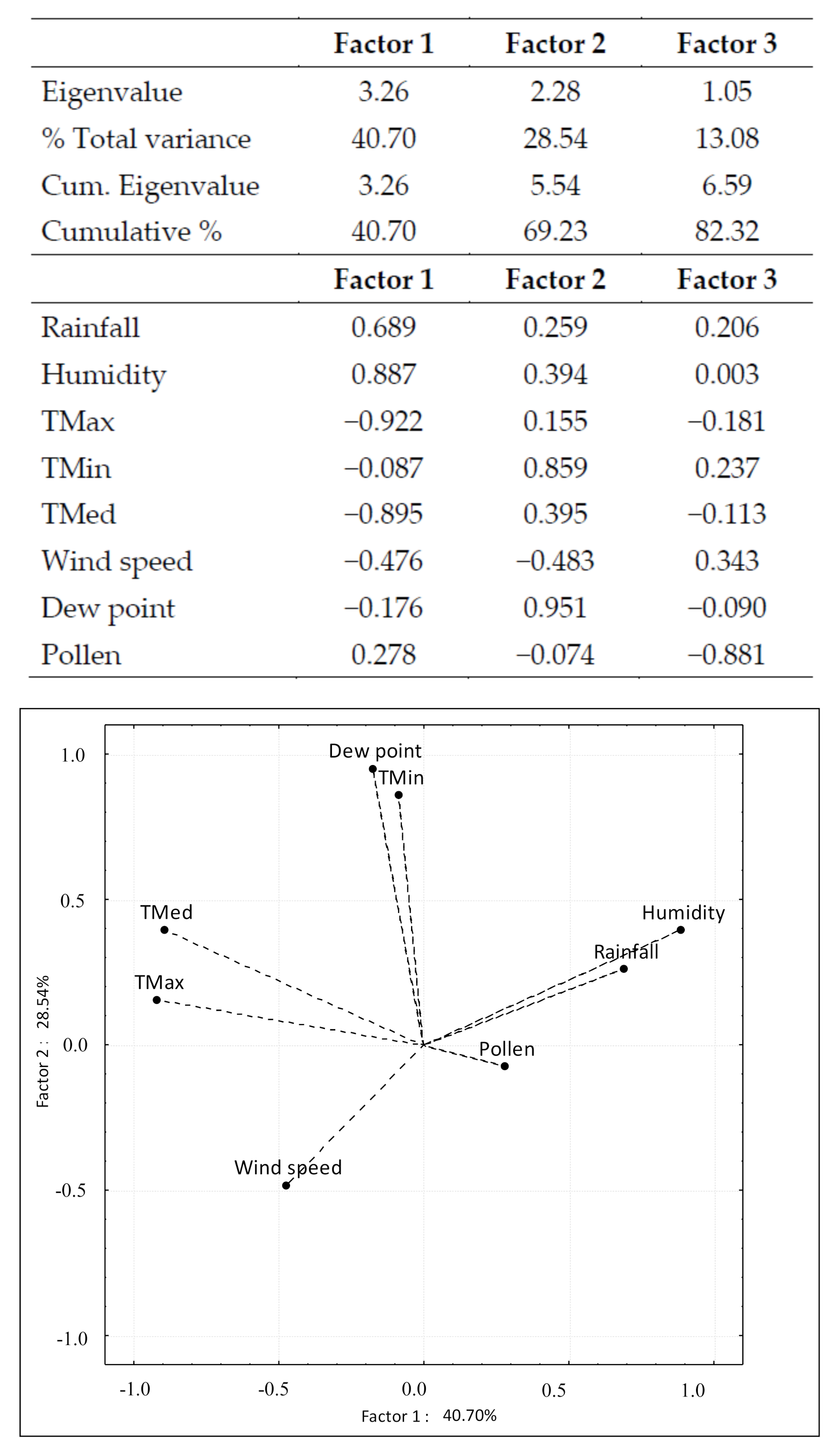
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galán, C.; García-Mozo, H.; Vázquez, L.; Ruiz-Valenzuela, L.; Díaz de la Guardia, C.; Trigo, M.M. Heat requirement for the onset of the Olea europaea L. pollen season in several sites of Andalusia and the effect of the expected future climate change. Int. J. Biometeorol. 2005, 49, 184–188. [Google Scholar] [CrossRef]
- Aguilera, F.; Ruiz-Valenzuela, L.; Fornaciari, M.; Romano, B.; Galán, C.; Oteros, J.; Ben Dihab, A.; Msallem, M.; Orlandi, F. Heat accumulation period in the Mediterranean region: Phenological response of the olive in different climate areas (Spain, Italy and Tunisia). Int. J. Biometeorol. 2014, 58, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Orlandi, F.; Aguilera, F.; Galán, C.; Msallem, M.; Fornaciari, M. Olive yields forecasts and oil price trends in mediterranean areas: A comprehensive analysis of the last two decades. Expl. Agric. 2017, 53, 71–83. [Google Scholar] [CrossRef]
- Loumou, A.; Giourga, C. Olive groves: The life and identity of the Mediterranean. Agric. Hum. Values 2003, 20, 87–95. [Google Scholar] [CrossRef]
- Rojo, J.; Pérez-Badia, R. Effects of topography and crown-exposure on olive tree phenology. Trees 2014, 28, 449–459. [Google Scholar] [CrossRef]
- García-Mozo, H.; Mestre, A.; Galán, C. Phenological trends in southern Spain: A response to climate change. Agric. For. Meteorol. 2010, 150, 575–580. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J.; Lobo, J.M. Determining the environmental factors underlying the spatial variability of insect appearance phenology for the honey bee, Apis mellifera, and the small white, Pieris rapae. J. Insect Sci. 2010, 10, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Dimou, M. Comparison of phenological, aerobiological and melissopalynological patterns of Olea europaea. Apidologie 2012, 43, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Frenguelli, G.; Bricchi, E. The use of the pheno-climatic model for forecasting the pollination of some arboreal taxa. Aerobiologia 1998, 14, 39–44. [Google Scholar] [CrossRef]
- Rodríguez-Rajo, F.J.; Frenguelli, G.; Jato, M.V. Effect of air temperature on forecasting the start of the Betula pollen season at two contrasting sites in the south of Europe (1995–2001). Int. J. Biometeorol. 2003, 47, 117–125. [Google Scholar] [CrossRef]
- Fernández-Escobar, R.; Benlloch, M.; Navarro, C.; Martín, G.C. The time of floral induction in the olive. J. Am. Soc. Hortic. Sci. 1992, 110, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Wareing, P.F.; Phillips, I.D.J. The Control of Growth and Differentiation in Plants, 2nd ed.; Pergamon Oxford: Oxford, UK, 1987. [Google Scholar]
- Chuine, I.; Cour, P.; Rousseau, D.D. Fitting models predicting dates of flowering of temperate-zone trees using simulated annealing. Plant Cell Environ. 1998, 21, 455–466. [Google Scholar] [CrossRef]
- Myking, T.; Heide, M. Dormancy release and chilling requirements of buds of latitudinal ecotypes of Betula pendula and B. pubescens. Tree Physiol. 1995, 15, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Chuine, I.; Cour, P. Climatic determinants of budburst seasonality in four temperate-zone tree species. New Phytol. 1999, 143, 339–349. [Google Scholar] [CrossRef]
- Cortiñas, J.A.; Fernández-González, M.; González-Fernández, E.; Vázquez-Ruiz, R.A.; Rodríguez-Rajo, F.J.; Aira, M.J. Potential Fertilization Capacity of Two Grapevine Varieties: Effects on Agricultural Production in Designation of Origin Areas in the Northwestern Iberian Peninsula. Agronomy 2020, 10, 961. [Google Scholar] [CrossRef]
- Spinardi, A.; Bassi, D. Olive Fertility as Affected by Cross-Pollination and Boron. Sci. World J. 2012, 2, 375631. [Google Scholar] [CrossRef] [Green Version]
- Tello, J.; Montemayor, M.I.; Forneck, A.; Ibáñez, J. A new image-based tool for the high throughput phenotyping of pollen viability: Evaluation of inter- and intra-cultivar diversity in grapevine. Plant Methods 2018, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Geelen, D. The impact of environmental stress on male reproductive development in plants: Biological processes and molecular mechanisms. Plant Cell Environ. 2014, 37, 1–18. [Google Scholar] [CrossRef]
- Castiñeiras, P.; Vázquez-Ruiz, R.A.; Fernández-González, M.; Rodríguez-Rajo, F.J.; Aira, M.J. Production and viability of Fraxinus pollen and its relationship with aerobiological data in the northwestern Iberian Peninsula. Aerobiologia 2019, 35, 227–241. [Google Scholar] [CrossRef]
- Ferrara, G.; Camposeo, S.; Palasciano, M.; Godini, A. Production of total and stainable pollen grains in Olea europaea L. Grana 2007, 46, 85–90. [Google Scholar] [CrossRef]
- Oteros, J.; Orlandi, F.; García-Mozo, H.; Aguilera, F.; Dihab, B.; Bonofiglio, T.; Abichou, M.; Ruiz-Valenzuela, L.; Trigo, M.; Díaz De la Guardia, M.M.; et al. Better prediction of Mediterranean olive production using pollen-based models. Agron. Sustain. Dev. 2014, 34, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Hiura, T. Flowering phenology and pollen fertility in an androdioecious tree, Fraxinus lanuginosa Koidz. (Oleaceae), in Hokkaido. Japan Int. J. Pl. Sci. 1998, 159, 941–947. [Google Scholar] [CrossRef]
- Koubouris, G.C.; Metzidakis, I.T.; Vasilakakis, M.D. Intraspecific variation in pollen viability, germination and ultrastructure of Olea europaea L. Afr. J. Biotechnol. 2012, 11, 13442–13446. [Google Scholar]
- Rojo, J.; Salido, P.; Pérez-Badia, R. Flower and pollen production in the ‘Cornicabra’ olive (Olea europaea L.) cultivar and the influence of environmental factors. Trees 2015, 29, 1235–1245. [Google Scholar] [CrossRef]
- Bonofiglio, T.; Orlandi, F.; Ruga, L.; Romano, B.; Fornaciari, M. Climate change impact on the olive pollen season in Mediterranean areas of Italy: Air quality in late spring from an allergenic point of view. Environ. Monit. Assess. 2013, 185, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Galán, C.; García-Mozo, H.; Vázquez, L.; Ruiz-Valenzuela, L.; Díaz de la Guardia, C.; Domínguez-Vilches, E. Modelling olive crop yield in Andalucía, Spain. Agron. J. 2008, 100, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Orlandi, F.; Sgromo, C.; Bonofiglio, T.; Ruga, L.; Romano, B.; Fornaciari, M. Yield modelling in a Mediterranean species utilizing cause-effect relationships between temperature forcing and biological processes. Sci. Hortic. 2010, 123, 412–417. [Google Scholar]
- Damialis, A.; Fotiou, C.; Halley, J.M.; Vokou, D. Effects of environmental factors on pollen production in anemophilous woody species. Trees 2011, 25, 253–264. [Google Scholar] [CrossRef]
- Rojo, J.; Pérez-Badía, R. Models for forecasting the flowering of Cornicabra olive groves. Int. J. Biometeorol. 2014, 59(11), 1547–1556. [Google Scholar] [CrossRef]
- Aguilera, F.; Ruiz-Valenzuela, L. Altitudinal fluctuations in the olive pollen emission: An approximation from olive groves of the south-east Iberian Peninsula. Aerobiologia 2012, 28, 403–411. [Google Scholar] [CrossRef]
- Garrido, A.; Fernández-González, M.; Álvarez-López, S.; González-Fernández, E.; Rodríguez-Rajo, F.J. First phenological and aerobiological assessment of olive orchards at the Northern limit of the Mediterranean bioclimatic area. Aerobiologia 2020, 36, 641–656. [Google Scholar] [CrossRef]
- Köppen, W. Versuch einer klasification der klimate, vorzugsweise nach iren Bezienhungen zur pflazenwelt. Geogr. Zeitschrift 1990, 6, 593. [Google Scholar]
- Geiger, R. Landolt-Börstein-Zahlenwerte und Funktionen aus Physik, Chemie, Astronomie, Geophysik und Technik; Alte serie Ch. Classification der climate nach W. Köppen; Springer: Berlin/Heidelberg, Germany, 1954; Volume 3, pp. 603–607. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; Mcmahon, T.A. Updated world map of the Köppen-Geiger climate classification-. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Cortizas, A.; Pérez Alberti, A. Atlas Climático de Galicia; Xunta de Galicia: Santiago de Compostela, Spain, 1999; pp. 1–207. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants, BBCH Monograph, 2nd ed; Federal Biological Research Centre for Agriculture and Forestry: Bonn, Germany, 2001. [Google Scholar]
- Sanz-Cortés, F.; Martínez-Calvo, J.; Badenes, M.L.; Bleiholder, H.; Hack, H.; Yacer, G.; Meier, V. Phenological growth stages of olive trees (Olea europaea). Ann. Appl. Biol. 2002, 140, 151–157. [Google Scholar] [CrossRef]
- Aron, R. Availability of chilling temperatures in California. Agric Meteorol 1983, 28, 351–363. [Google Scholar] [CrossRef]
- Jato, V.; Rodríguez-Rajo, F.J.; Méndez, J.; Aira, M.J. Phenological behaviour of Quercus in Ourense (NW Spain) and its relationship with the atmospheric pollen season. Int. J. Biometeorol. 2002, 46, 176–184. [Google Scholar] [PubMed]
- Fernández-González, M.; Rodríguez-Rajo, F.J.; Escuredo, O.; Aira, M.J. Influence of thermal requirement in the aerobiological and phenological behavior of two grapevine varieties. Aerobiologia 2013, 29, 523–535. [Google Scholar] [CrossRef]
- Galán, C.; García-Mozo, H.; Cariñanos, P.; Alcázar, P.; Domínguez, E. The role of temperature in the onset of the Olea europaea L. pollen season in south-western Spain. Int. J. Biometeorol. 2001, 45, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Ring, D.R.; Harris, M.K.; Jackman, J.A.; Henson, J.L. A Fortran Computer Program for Determining Start Date and Base Temperature for Degree-Day Models. The Texas; Agricultural Experiment Station Bull MP-1537, College Station, Texas; The Texas University System: Austin, TX, USA, 1983. [Google Scholar]
- de Oliveira da Silva, L.F.; Ruiz-Zambon, C.; de Oliveira, A.F.; Pio, R.; Dias-Gonçalves, E. Establishment of growth medium and quantification of pollen grains of olive cultivars in Brazil’s subtropical areas. Bragantia 2016, 75, 26–32. [Google Scholar] [CrossRef]
- Dalkiliç, Z.; Mestav, H.O. In vitro pollen quantity, viability and germination tests in quince. Afr. J. Biotechnol. 2011, 10, 16516–16520. [Google Scholar]
- Gaaliche, B.; Majdoub, A.; Trad, M.; Mars, M. Assessment of pollen viability, germination, and tube growth in eight Tunisian caprifigs (Ficus carica L.) cultivars. ISRN Agronomy 2013, 5. [Google Scholar]
- Brewbaker, J.L.; Kwack, B.H. The calcium ion and substances influencing pollen growth. In Pollen Physiology and Fertilization; Linskens, H.F., Ed.; Elsevier: Amsterdam, The Netherlands, 1964. [Google Scholar]
- Rodriguez-Riano, T.; Dafni, A. A new procedure to asses pollen viability. Sex Plant Reprod. 2000, 12, 241–244. [Google Scholar] [CrossRef]
- Rejón, J.D.; Suárez, C.G.; Alché, J.D.; Castro, A.J.; Rodríguez-García, M.I. Evaluación de diferentes métodos para estimar la calidad del polen en distintos cultivares de olivo (Olea europaea L.). Polen 2010, 20, 61–72. [Google Scholar]
- Shivanna, K.R.; Rangaswamy, N.S. Pollen biology: A Laboratory Manual; Library of Congress Cataloging-in-Publication: Berlin, Germany, 1992; p. 119. [Google Scholar]
- Hirst, J.M. An automatic volumetric spore-trap. Ann. Appl. Biol. 1952, 36, 257–265. [Google Scholar] [CrossRef]
- Galán, C.; Cariñanos, P.; Alcázar Teno, P.; Domínguez Vilches, E. Spanish Aerobiology Network (REA): Management and Quality Manual; University of Córdoba Publication Service: Cordoba, Spain, 2007. [Google Scholar]
- Andersen, T.B. A model to predict the beginning of the pollen season. Grana 1991, 30, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Tipping, M.E.; Bishop, C.M. Probabilistic principal component analysis. J. R. Stat. Soc. B. 1999, 61, 611–622. [Google Scholar] [CrossRef]
- Sabatier, R.; Lebreton, J.D.; Chessel, D. Principal component analysis with instrumental variables as a tool for modelling composition data. In Multiway Data Analysis; Coppi, R., Bolasco, S., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 341–352. [Google Scholar]
- Rojo, J.; Orlandini, F.; Ben Dhiab, A.; Lara, B.; Picornell, A.; Oteros, J.; Msallem, M.; Fornaciari, M.; Pérez-Badía, R. Estimation of Chilling and Heat Accumulation Periods Based on the Timing of Olive Pollination. Forests 2020, 11, 835. [Google Scholar] [CrossRef]
- Osborne, C.P.; Chuine, I.; Viner, D.; Woodward, F.I. Olive phenology as a sensitive indicator of future climatic warming in the Mediterranean. Plant Cell Environ. 2000, 23, 1–710. [Google Scholar] [CrossRef] [Green Version]
- Achmakh, L.; Bouziane, H.; Aboulaich, N. Airborne pollen of Olea europaea L. in Tetouan (NW Morocco): Heat requirements and forecasts. Aerobiologia 2015, 31, 191–199. [Google Scholar] [CrossRef]
- de la Rosa, R.; Rallo, L.; Rapoport, H.F. Olive Floral Bud Growth and Starch Content During Winter Rest and Spring Budbreak. HortScience 2000, 35, 1223–1227. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, A.; Alerci, L. Reproductive and vegetative bud differentiation in Olea europaea L. J. Hortic. Sci. Biotechnol. 1999, 74, 522–527. [Google Scholar] [CrossRef]
- Pinney, K.; Polito, V.S. Flower initiation in Manzanilla Olive. Acta Hortic. 1990, 203–206. [Google Scholar] [CrossRef]
- Orlandi, F.; Fornaciari, M.; Romano, B. The use of phenological data to calculate chilling units in Olea europaea L. in relation to the onset of reproduction. Int. J. Biometeorol. 2002, 46, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Orlandi, F.; Lanari, D.; Romano, B.; Fornaciari, M. New model to predict the timing of olive (Olea europaea) flowering: A case study in central Italy. NZ J. Crop. Hortic. Sci. 2006, 34, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Wielgolaski, F.E. Starting dates and basic temperatures in phenological observations of plants. Int. J. Biometeorol. 1999, 42, 158–168. [Google Scholar] [CrossRef]
- Bonofiglio, T.; Orlandi, F.; Sgromo, C.; Romano, B.; Fornaciari, M. Influence of temperature and rainfall on timing of olive (Olea europaea) flowering in southern Italy. NZ J. Crop Hortic. Sci. 2008, 36, 59–69. [Google Scholar] [CrossRef]
- Bonofiglio, T.; Orlandi, F.; Sgromo, C.; Romano, B.; Fornçaciari, M. Evidences of olive pollination date variations in relation to spring temperature trends. Aerobiologia 2009, 25, 227–237. [Google Scholar] [CrossRef]
- Lang, G.A. Dormancy: A new universal terminology. Hortscience 1987, 22, 817–820. [Google Scholar]
- Hackett, P.W.; Hartmann, H.T. The influence of temperature on floral initiation in the olive. Physiology Plantarum 1967, 20, 430–436. [Google Scholar] [CrossRef]
- Ribeiro, H.; Cunha, M.; Abreu, I. Comparison of classical models for evaluating the heat requirements for olive (Olea europaea L.) in Portugal. J. Integr. Plant Biol. 2006, 48, 664–671. [Google Scholar] [CrossRef]
- Crepinsek, Z.; Kajfez-Bogataj, L.; Bergant, K. Modelling of weather variability effect on fitophenology. Ecol. Model. 2006, 194, 256–265. [Google Scholar] [CrossRef]
- De Melo-Abreu, J.P.; Barranco, D.; Cordeiro, A.M.; Tous, J.; Rogado, B.M.; Villalobos, F.J. Modelling olive flowering date using chilling for dormancy release and thermal time. Agric. For. Meteorol. 2004, 125, 117–127. [Google Scholar] [CrossRef]
- Martins, F.B.; Reis, D.F.; Pinheiro, M.V.M. Base temperature and phyllochron in two olive cultivars. Cientia Rural 2012, 42, 1975–1981. [Google Scholar] [CrossRef] [Green Version]
- Ruml, M.; Vukovic, A.; Milatovic, D. Evaluation of different methods for determining growing degree-day thresholds in apricot cultivars. Int. J. Biometeorol. 2010, 54, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Vuletin, G.; Perica, S.; Goreta, S.; Bucan, L. Flower sterility and the germination ability of pollen as genetic traits of seven olive (Olea europaea L.) cultivars grown in Croatia. J. Horticult. Sci. Biotechnol. 2012, 87, 237–242. [Google Scholar] [CrossRef]
- Tormo, R.; Muñóz, A.; Silva, I.; Gallardo, F. Pollen production in anemophilous trees. Grana 1996, 35, 38–46. [Google Scholar]
- Mazzeo, A.; Palasciano, M.; Gallota, A.; Camposeo, S.; Pacifico, A.; Ferrara, G. Amount and quality of pollen grains in four olive (Olea europaea L.) cultivars as affected by ‘on’ and ‘off’ years. Sci. Hortic. 2014, 170, 89–93. [Google Scholar] [CrossRef]
- Wu, S.B.; Collins, G.; Sedgley, M. Sexual compatibility within and between olive cultivars. J. Hortic. Sci. Biotech. 2002, 77, 665–673. [Google Scholar] [CrossRef]
- Bartolini, S.; Guerriero, R. Self-compatibility in several clones of oil olive cv. leccino. Adv. Hortic. Sci. 1995, 9, 71–74. [Google Scholar]
- Fernández-González, M.; González-Fernández, E.; Fernández-González, D.; Rodríguez-Rajo, F.J. Secondary outcomes of the Ole e 1 proteins involved in pollen tube development: Impact on allergies. Front. Plant Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, F.; Fornaciari, M.; Ruiz-Valenzuela, L.; Galán, C.; Msallem, M.; Ben Dihab, A.; Díaz De la Guardia, C.; Trigo, M.; Bonofiglio, T.; Orlandi, F. Phenological models to predict the main flowering phases of olive (Olea europea L.) along a latitudinal and longitudinal gradient across the Mediterranean region. Int. J. Biometeorol. 2015, 59, 629–641. [Google Scholar] [CrossRef]
- Aguilera, F.; Ruiz Valenzuela, L. of the floral phenology of Olea europaea L. in Jaen province (SE Spain) and its relation with pollen emission. Aerobiologia 2009, 25, 217–225. [Google Scholar] [CrossRef]
- Elvira-Rendueles, B.; Moreno, J.M.; Costa, I.; Bañón, D.; Martínez-García, M.J.; Moreno-Grau, S. Pollen calendars of Cartagena, Lorca, and Murcia (Region of Murcia), southeastern Iberian Peninsula: 2010–2017. Aerobiologia 2019, 35, 477–496. [Google Scholar] [CrossRef]
- Alba, F.; Díaz de la Guardia, C. The effect of air temperature on the starting dates of the Ulmus, Platanus and Olea pollen seasons in the SE Iberian Peninsula. Aerobiologia 1998, 14, 191–194. [Google Scholar] [CrossRef]
- Fernández-Rodríguez, S.; Durán-Barroso, P.; Silva-Palacios, I.; Tormo-Molina, R.; Maya-Manzano, J.M.; Gonzalo-Garijo, Á. Forecast model of allergenic hazard using trends of Poaceae airborne pollen over an urban area in SW Iberian Peninsula (Europe). Nat. Hazards 2016, 84, 121–137. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chill Requirements | ||||||||
|---|---|---|---|---|---|---|---|---|
| Th °C | 2016 | 2017 | 2018 | Mean | Min | Max | SD | C.S.% |
| 0 | 747 | 761 | 754 | 747 | 761 | 9.6 | 1.3 | |
| 0.5 | 185 | 757 | 748 | 564 | 185 | 757 | 327.9 | 58.2 |
| 1 | 423 | 747 | 730 | 634 | 423 | 747 | 182.4 | 28.8 |
| 1.5 | 578 | 729 | 713 | 673 | 578 | 729 | 82.7 | 12.3 |
| 2 | 669 | 710 | 703 | 694 | 669 | 710 | 21.9 | 3.2 |
| 2.5 | 712 | 698 | 703 | 704 | 698 | 712 | 7.0 | 1.0 |
| 3 | 722 | 698 | 716 | 712 | 698 | 722 | 12.3 | 1.7 |
| 3.5 | 712 | 713 | 743 | 723 | 712 | 743 | 17.7 | 2.5 |
| 4 | 693 | 745 | 784 | 741 | 693 | 784 | 45.9 | 6.2 |
| 4.5 | 674 | 793 | 837 | 768 | 674 | 837 | 84.6 | 11.0 |
| 5 | 662 | 855 | 899 | 805 | 662 | 899 | 126.1 | 15.7 |
| 5.5 | 662 | 925 | 966 | 851 | 662 | 966 | 164.8 | 19.4 |
| 6 | 678 | 998 | 1032 | 903 | 678 | 1032 | 195.1 | 21.6 |
| 6.5 | 711 | 1065 | 1090 | 955 | 711 | 1090 | 211.8 | 22.2 |
| 7 | 760 | 1116 | 1131 | 1002 | 760 | 1131 | 210.1 | 21.0 |
| 7.5 | 822 | 1139 | 1145 | 1035 | 822 | 1145 | 185.1 | 17.9 |
| 8 | 893 | 1121 | 1121 | 1045 | 893 | 1121 | 131.9 | 12.6 |
| 8.5 | 966 | 1045 | 1047 | 1019 | 966 | 1047 | 46.2 | 4.5 |
| 9 | 1033 | 892 | 907 | 944 | 892 | 1033 | 76.9 | 8.1 |
| 9.5 | 1083 | 645 | 688 | 805 | 645 | 1083 | 241.3 | 30.0 |
| 10 | 1104 | 281 | 371 | 585 | 281 | 1104 | 451.6 | 77.2 |
| Pollen Season | 2016 | 2017 | 2018 | Average | S.D. | C.S.% | |
|---|---|---|---|---|---|---|---|
| Duration | Start | 23-May | 7-May | 30-May | 20-May | 11.79 | 0.03 |
| End | 14-June | 16-June | 20-June | 16-June | 3.06 | 0.01 | |
| Total pollen grains | 750 | 947 | 801 | 833 | 102.25 | 12.28 | |
| Peak day | 1-June | 30-May | 5-June | 1-June | 3.06 | 0.01 | |
| Polen peak day (pollen/m3) | 166 | 135 | 138 | 146 | 17.10 | 11.68 | |
| Growing Degreee Days (GDD) Pollen Season | Start | 1192 | 1053 | 1238 | 1161 | 96.24 | 8.29 |
| End | 1558 | 1785 | 1581 | 1641 | 125.02 | 7.62 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, A.; Fernández-González, M.; Vázquez-Ruiz, R.A.; Rodríguez-Rajo, F.J.; Aira, M.J. Reproductive Biology of Olive Trees (Arbequina cultivar) at the Northern Limit of Their Distribution Areas. Forests 2021, 12, 204. https://doi.org/10.3390/f12020204
Garrido A, Fernández-González M, Vázquez-Ruiz RA, Rodríguez-Rajo FJ, Aira MJ. Reproductive Biology of Olive Trees (Arbequina cultivar) at the Northern Limit of Their Distribution Areas. Forests. 2021; 12(2):204. https://doi.org/10.3390/f12020204
Chicago/Turabian StyleGarrido, Alejandro, María Fernández-González, Rosa A. Vázquez-Ruiz, F. Javier Rodríguez-Rajo, and María J. Aira. 2021. "Reproductive Biology of Olive Trees (Arbequina cultivar) at the Northern Limit of Their Distribution Areas" Forests 12, no. 2: 204. https://doi.org/10.3390/f12020204
APA StyleGarrido, A., Fernández-González, M., Vázquez-Ruiz, R. A., Rodríguez-Rajo, F. J., & Aira, M. J. (2021). Reproductive Biology of Olive Trees (Arbequina cultivar) at the Northern Limit of Their Distribution Areas. Forests, 12(2), 204. https://doi.org/10.3390/f12020204