Author Contributions
Conceptualization, C.S.-P., N.G. and C.B.; methodology, C.S.-P. and N.G.; validation, C.S.-P., N.G. and C.B.; formal analysis, C.S.-P. and N.G.; investigation, N.G. and C.S.-P.; resources, N.G.; writing—original draft preparation, C.S.-P.; writing—review and editing, C.S.-P. and N.G.; funding acquisition, N.G. and C.B. All authors have read and agreed to the published version of the manuscript.
Appendix A
Needle whorls (WHL)—The number of needle ages retained on the branchlet.
Branchlet diameter (BRDIA2)—Diameter measured in mm (±0.01 mm) at the base of the previous year branchlet.
Branchlet elongation (BRLN1)—Measured length of the current year branchlet (±2.0 mm)
Relative needle length (%MxNL1)—Current-year needle length expressed as a percent of the average needle length of the whorl with the longest needles (±5.0 mm).
Chlorosis in 4-year needles (CHL4)—Chlorosis level of 4-year-old needles expressed as a percent of healthy, green needles in the same age class by ocular estimation.
Insect defoliators (LF BI DF)—The summed frequency of foliar (biotic) insect defoliators (weevils, phloem feeders, and scale).
Abiotic defoliators (LF A DF)—The summed frequency of needle tip dieback, dead needles, and early senescence (related to drought stress).
Tree–tree competition (4CZDPP)—average diameter/distance to the nearest four conspecific neighboring trees.
Dwarf mistletoe rank (DMR)—dwarf mistletoe infection ranking (Hawksworth, 1977).
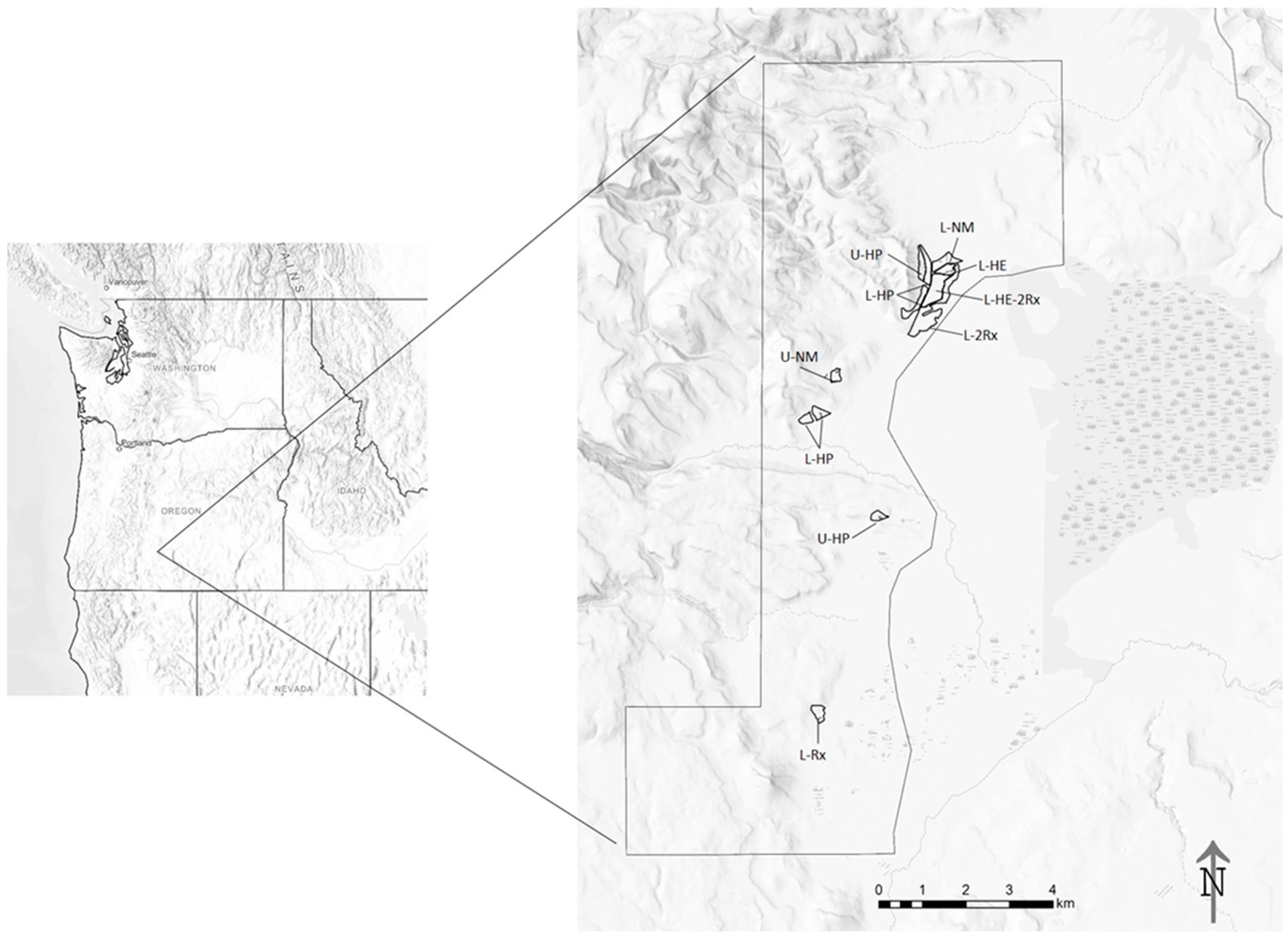
Figure 1.
Map of our study area (8824 ha) in south–central Oregon. Elevations ranged from 1520 m to 1719 m. Locations of stands in the study area where trees were sampled are presented in the large-scale map on the right. Key to stand codes: L = lowland, U = upland, HE = harvest, even spacing, HP = harvest, patchy (clumps and openings), Rx = prescribed fire, NM = no management, no harvest or fire.
Figure 1.
Map of our study area (8824 ha) in south–central Oregon. Elevations ranged from 1520 m to 1719 m. Locations of stands in the study area where trees were sampled are presented in the large-scale map on the right. Key to stand codes: L = lowland, U = upland, HE = harvest, even spacing, HP = harvest, patchy (clumps and openings), Rx = prescribed fire, NM = no management, no harvest or fire.
Figure 2.
Examples of qualitative visual characteristics used to assess trees to vigor classes. (
A) The tree on the right exhibits chlorosis and low needle retention, while the tree on the left has good needle color and mass, (
B) reduced needle elongation and fading, (
C) needle senescence, and (
D) low needle retention (from [
32]).
Figure 2.
Examples of qualitative visual characteristics used to assess trees to vigor classes. (
A) The tree on the right exhibits chlorosis and low needle retention, while the tree on the left has good needle color and mass, (
B) reduced needle elongation and fading, (
C) needle senescence, and (
D) low needle retention (from [
32]).
Figure 3.
Aerial imagery collected in October 2017 (four (R, G, B, IR) bands, 30 cm resolution) with classified individual tree crowns (red = low vigor, blue = not low vigor). This image contains four different site treatments; on the upper right is L-NM, lower right L-HE-2Rx, left of the main road is L-HP, and between the two spur roads on the right is L-HE.
Figure 3.
Aerial imagery collected in October 2017 (four (R, G, B, IR) bands, 30 cm resolution) with classified individual tree crowns (red = low vigor, blue = not low vigor). This image contains four different site treatments; on the upper right is L-NM, lower right L-HE-2Rx, left of the main road is L-HP, and between the two spur roads on the right is L-HE.
Figure 4.
Graph of variable importance produced by the Random Forest model. Vertical axis values are the predictor variables and the horizontal axis shows the decrease in model accuracy if the variable is removed.
Figure 4.
Graph of variable importance produced by the Random Forest model. Vertical axis values are the predictor variables and the horizontal axis shows the decrease in model accuracy if the variable is removed.
Figure 5.
Graph of the proportion of LOW tree crowns (ITCs) in the eight stands with standard error. Stands that are not significantly different (two tailed Z-score tests of a proportion, p > 0.01) have the same italicized letter (a–e). For example, U-NM and L-Rx, are not significantly different; U-HP, L-2Rx and L-HE are not significantly different. Stand treatment descriptions are as follows: U-NM is upland, no management; L-Rx is lowland prescribed fire (2008); L-HP is lowland, harvest in 2016, retention of trees in groups; L-NM is lowland, no management; U-HP is upland, harvest in 2016, retention of trees in groups; L-2Rx is lowland, prescribed fire in 2006 and 2013; L-HE is lowland, harvest even spacing; and L-HE-2Rx is lowland, harvest even spacing, prescribed fire in 2006 and 2013.
Figure 5.
Graph of the proportion of LOW tree crowns (ITCs) in the eight stands with standard error. Stands that are not significantly different (two tailed Z-score tests of a proportion, p > 0.01) have the same italicized letter (a–e). For example, U-NM and L-Rx, are not significantly different; U-HP, L-2Rx and L-HE are not significantly different. Stand treatment descriptions are as follows: U-NM is upland, no management; L-Rx is lowland prescribed fire (2008); L-HP is lowland, harvest in 2016, retention of trees in groups; L-NM is lowland, no management; U-HP is upland, harvest in 2016, retention of trees in groups; L-2Rx is lowland, prescribed fire in 2006 and 2013; L-HE is lowland, harvest even spacing; and L-HE-2Rx is lowland, harvest even spacing, prescribed fire in 2006 and 2013.
Table 1.
Description of stands in the study area where trees were sampled. Key to stand codes: L = lowland, U = upland, HE = harvest, even spacing, HP = harvest, patchy (clumps and openings), Rx = prescribed fire, NM = no management, no harvest or fire. Diameter at breast height (1.37 m above ground, DBH), BA (basal area), and trees per hectare (TPH) data from [
32]. These data are based on the DBH and species of each tree in three 30 m radius plots in each site. Values are the mean ± SD. PP = ponderosa pine. N/A indicates that no belt transects were conducted in these stands.
Table 1.
Description of stands in the study area where trees were sampled. Key to stand codes: L = lowland, U = upland, HE = harvest, even spacing, HP = harvest, patchy (clumps and openings), Rx = prescribed fire, NM = no management, no harvest or fire. Diameter at breast height (1.37 m above ground, DBH), BA (basal area), and trees per hectare (TPH) data from [
32]. These data are based on the DBH and species of each tree in three 30 m radius plots in each site. Values are the mean ± SD. PP = ponderosa pine. N/A indicates that no belt transects were conducted in these stands.
| Stand | Harvest | Rx Burn | Mean DBH (cm) | PP BA (m2/ha) | All Species BA (m2/ha) | PP TPH | All Species TPH |
|---|
| U-NM | N/A | N/A | 35.2 ± 4.0 | 61.4 ± 80.4 | 61.4 ± 80.4 | 559 ± 361 | 559 ± 361 |
| U-HP | 2016 | N/A | 34.6 ± 3.5 | 34.8 ± 15.0 | 34.8 ± 15.0 | 307 ± 50 | 307 ± 50 |
| L-NM | N/A | N/A | 21.2 ± 1.7 | 27.6 ± 12.8 | 30.0 ± 13.8 | 605 ± 237 | 722 ± 289 |
| L-HP | 2016 | N/A | 41.2 ± 2.8 | 18.6 ± 4.9 | 18.6 ± 4.9 | 160 ± 36 | 160 ± 36 |
| L-HE | 2005 | N/A | 37.5 ± 0.8 | 8.8 ± 3.8 | 8.8 ± 3.8 | 137 ± 57 | 137 ± 57 |
| L-HE-2Rx | 2005 | 2006,
2013 | 38.5 ± 1.8 | 17.3 ± 5.7 | 17.3 ± 5.7 | 146 ± 16 | 146 ± 16 |
| L-Rx | N/A | 2008 | 23.9 ± 1.6 | 5.51 ± 1.0 | 16.0 ± 1.2 | 434 ± 22 | 566 ± 14 |
| L-2Rx | N/A | 2006, 2013 | 41.4 ± 1.9 | 28 ± 8.0 | 28 ± 8.0 | 160 ± 36 | 160 ± 36 |
Table 2.
Whole-tree and branch characteristics used in assigning qualitative-vigor classes to ponderosa pine individual tree crowns (ITCs). DMR is the dwarf mistletoe rating.
Table 2.
Whole-tree and branch characteristics used in assigning qualitative-vigor classes to ponderosa pine individual tree crowns (ITCs). DMR is the dwarf mistletoe rating.
| LOW | NOT LOW |
|---|
| Low number of whorls—low needle retention. | Highest number of whorls—high needle retention. |
| High level of needle chlorosis, fading, yellowing. | Low or no visible needle chlorosis, no visible fading. |
| Thinner branchlets. | Thicker branchlets. |
| High frequency of early needle senescence (browning). | Low frequency of early needle senescence (browning). |
| Low needle mass. | High needle mass. |
| High DMR rank. | Low DMR rank. |
Table 3.
Spectral indices calculated from the four-band imagery and used in the vigor model and classification of Ponderosa pine individual tree crowns (ITCs). NIR = near-infrared band, R = red band, B = blue band, G = green band.
Table 3.
Spectral indices calculated from the four-band imagery and used in the vigor model and classification of Ponderosa pine individual tree crowns (ITCs). NIR = near-infrared band, R = red band, B = blue band, G = green band.
| Index | Formula | Citation |
|---|
| Normalized Differential Vegetation Index (NDVI) | (NIR-R)/(NIR ± R) | Rouse et al., 1974; Tucker 1979 |
| Enhanced Vegetation Index (EVI) | 2.5(NIR-R)/(NIR ±(2.4R) ±1) | Jiang et al., 2008. |
| Difference Vegetation Index (DVI) | NIR-R | Richardson and Wiegand 1977. |
| Blue Chromatic Coordinate (BCC) | B/(R ± G ± B) | Gillespie et al., 1987; Stafford 2015. |
| Red Chromatic Coordinate (RCC) | R/(R ± G ± B) | Gillespie et al. 1987; Stafford 2015. |
| Green Chromatic Coordinate (GCC) | G/(R ± G ± B) | Gillespie et al., 1987; Stafford 2015; Reid et al., 2016. |
Table 4.
Individual tree crown (ITC) spectra/index attribute values (MEAN and SE) for trees in the two vigor classes. Marked (*) attributes are significantly different (p < 0.05) between the LOW and NOT LOW vigor classes using the Mann–Whitney test for differences in means. Indices defined: DVI = difference vegetation index; NDVI = normalized differential vegetation index; EVI=enhanced vegetation index; RCC = red chromatic coordinate; BCC = blue chromatic coordinate; GCC = green chromatic coordinate. SD indicates the standard deviation of the pixels within an ITC. For example, GREEN SD is the standard deviation of the green value of all the pixels within an ITC.
Table 4.
Individual tree crown (ITC) spectra/index attribute values (MEAN and SE) for trees in the two vigor classes. Marked (*) attributes are significantly different (p < 0.05) between the LOW and NOT LOW vigor classes using the Mann–Whitney test for differences in means. Indices defined: DVI = difference vegetation index; NDVI = normalized differential vegetation index; EVI=enhanced vegetation index; RCC = red chromatic coordinate; BCC = blue chromatic coordinate; GCC = green chromatic coordinate. SD indicates the standard deviation of the pixels within an ITC. For example, GREEN SD is the standard deviation of the green value of all the pixels within an ITC.
| SPECTRA/INDEX | LOW | NOT LOW | p-Value |
|---|
| BLUE_MEAN | 124 (0.27) | 124 (0.24) | 0.85 |
| RED_MEAN | 143 (0.45) | 143 (0.35) | 0.68 |
| GREEN_MEAN | 145 (0.37) | 146 (0.29) | 0.42 |
| NIR MEAN * | 221 (0.31) | 222 (0.24) | 0.0057 |
| DVI MEAN * | 77 (0.40) | 78 (0.33) | 0.0089 |
| NDVI MEAN * | 0.21 (0.0013) | 0.21 (0.0010) | 0.039 |
| EVI MEAN * | 0.34 (0.0023) | 0.34 (0.0019) | 0.036 |
| RCC MEAN * | 0.35 (0.00036) | 0.35 (0.00026) | 0.012 |
| BCC MEAN | 0.30 (0.00033) | 0.30 (0.00022) | 0.997 |
| GCC MEAN | 0.35 (0.00024) | 0.35 (0.00017) | 0.0011 |
| BLUE SD * | 2.7 (0.12) | 3.4 (0.076) | 0.0000088 |
| RED SD * | 4.2 (0.16) | 5.1 (0.11) | 0.000024 |
| GREEN SD * | 3.9 (0.15) | 4.5 (0.092) | 0.0097 |
| NIR SD | 5.0 (0.23) | 5.4 (0.15) | 0.14 |
| NIRR SD * | 4.1 (0.23) | 5.0 (0.14) | 0.00036 |
| NDVI SD * | 0.012 (0.00060) | 0.015 (0.00038) | 0.000018 |
| EVI SD * | 0.021 (0.0010) | 0.026 (0.00066) | 0.000014 |
| RCC SD * | 0.0026 (0.00010) | 0.0032 (7.8 × 10−5) | 0.0000076 |
| BCC SD * | 0.0027 (9.9 × 10−5) | 0.0029 (6.1 × 10−5) | 0.026 |
| GCC SD * | 0.0020 (6.8 × 10−5) | 0.0023 (4.9 × 10−5) | 0.0053 |
Table 5.
Tree-vigor attribute values (MEANS and SE) for trees in the two vigor classes. Marked (*) attributes significantly differed (
p < 0.05) using the Wilcoxon rank sum test. Tree-vigor attribute attributes were defined as follows: WHL is the number of years needles retained; BRDIA2 is the previous year branchlet diameter (±/− 0.01mm); BRLN1 is the current year branchlet length (±/− 2.0 mm); %MxNL1 is the current-year needle length relative to the longest needles produced on the branchlet; CHL4 is the level of chlorosis in 4-year-old needles; LF BI DF is a synthetic index of foliar (biotic) insect defoliators (including summed frequency of weevil, unknown phloem feeders, and scale); LF A DF is the summed frequency of needle tip dieback, dead needles, and early senescence; DMR is the dwarf mistletoe rating [
32].
Table 5.
Tree-vigor attribute values (MEANS and SE) for trees in the two vigor classes. Marked (*) attributes significantly differed (
p < 0.05) using the Wilcoxon rank sum test. Tree-vigor attribute attributes were defined as follows: WHL is the number of years needles retained; BRDIA2 is the previous year branchlet diameter (±/− 0.01mm); BRLN1 is the current year branchlet length (±/− 2.0 mm); %MxNL1 is the current-year needle length relative to the longest needles produced on the branchlet; CHL4 is the level of chlorosis in 4-year-old needles; LF BI DF is a synthetic index of foliar (biotic) insect defoliators (including summed frequency of weevil, unknown phloem feeders, and scale); LF A DF is the summed frequency of needle tip dieback, dead needles, and early senescence; DMR is the dwarf mistletoe rating [
32].
| RANK | WHL * | BRDIA2 | BRLN1 | %MxNL1 | CHL4 * | LF BI DF | LF A DF | DMR |
|---|
| NOT LOW | 6.3 (0.09) | 6.1 (0.10) | 14 (0.54) | 89.9 (0.89) | 8 (1.0) | 2.7 (0.05) | 0.67 (0.05) | 0.0 (0.10) |
| LOW | 6.0 (0.2) | 5.7 (0.21) | 13 (1.1) | 89 (1.7) | 13 (3.8) | 2.3 (0.14) | 1 (0.12) | 0.0 (0.27) |
Table 6.
OOB (out of bag) error matrix for the Random Forest model vigor predictions for all ITCs (individual tree crowns, outliers removed). The columns are the crowns as predicted by the model and the rows are the vigor assessments made in the field (reference data). The overall accuracy of 0.78 is calculated as the number of correctly classified crowns divided by the total number of crowns. The KAPPA statistic is the overall accuracy adjusted for random correct classification.
Table 6.
OOB (out of bag) error matrix for the Random Forest model vigor predictions for all ITCs (individual tree crowns, outliers removed). The columns are the crowns as predicted by the model and the rows are the vigor assessments made in the field (reference data). The overall accuracy of 0.78 is calculated as the number of correctly classified crowns divided by the total number of crowns. The KAPPA statistic is the overall accuracy adjusted for random correct classification.
| | LOW | NOT LOW | ROW TOTAL | USERS ACCURACY | Error of Commission (Type II) |
|---|
| LOW | 33 | 69 | 102 | 0.32 | 0.68 |
| NOT LOW | 18 | 251 | 269 | 0.93 | 0.07 |
| COLUMN TOTAL | 51 | 320 | 371 | | |
| PRODUCERS ACCURACY | 0.65 | 0.78 | | | |
| Error of Omission (Type I) | 0.35 | 0.22 | | | |
| | | ACCURACY: | 0.77 | | |
| | | KAPPA STATISTIC: | 0.30 | | |
Table 7.
Proportion of LOW RF-modeled individual tree crowns (ITCs) in upland and lowland sites. The proportion of LOW crowns was significantly different (two tailed Z-score tests of a proportion, p = 0.023) for the two site types.
Table 7.
Proportion of LOW RF-modeled individual tree crowns (ITCs) in upland and lowland sites. The proportion of LOW crowns was significantly different (two tailed Z-score tests of a proportion, p = 0.023) for the two site types.
| | LOW | NOT LOW | TOTAL | LOW/All Crowns |
|---|
| Lowland | 5096 | 19855 | 24951 | 0.20 |
| Upland | 1768 | 6419 | 8187 | 0.22 |
Table 8.
Proportion of LOW trees in each stand type as determined by the Random Forest model using spectral data (RF model), field qualitative assessment of the belt transect trees (n = 108) and 298 additional trees (All trees), and field qualitative assessment of 30 trees in a belt transect (Belt transect trees). Stand codes are L = lowland; U = upland; HP = harvested, patchy; HE = harvested, even spacing; NM = no management, Rx = prescribed burn. N/A indicates that no belt transects were conducted in this stand.
Table 8.
Proportion of LOW trees in each stand type as determined by the Random Forest model using spectral data (RF model), field qualitative assessment of the belt transect trees (n = 108) and 298 additional trees (All trees), and field qualitative assessment of 30 trees in a belt transect (Belt transect trees). Stand codes are L = lowland; U = upland; HP = harvested, patchy; HE = harvested, even spacing; NM = no management, Rx = prescribed burn. N/A indicates that no belt transects were conducted in this stand.
| STAND | RF Model | All Trees | Belt Transect Trees |
|---|
| U-NM | 0.32 | 0.20 | 0.18 |
| U-HP | 0.18 | 0.24 | 0.18 |
| L-NM | 0.26 | 0.35 | 0.40 |
| L-HP | 0.22 | 0.61 | 0.05 |
| L-HE | 0.16 | 0.28 | 0.13 |
| L-HE-2Rx | 0.14 | 0.04 | 0.07 |
| L-Rx | 0.34 | 0.35 | N/A |
| L-2Rx | 0.2 | 0.14 | 0.07 |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}