
Asynchrony and Time-Lag between Primary and Secondary Growth of Norway Spruce Growing in Different Elevations


Abstract
:1. Introduction
2. Materials and Methods
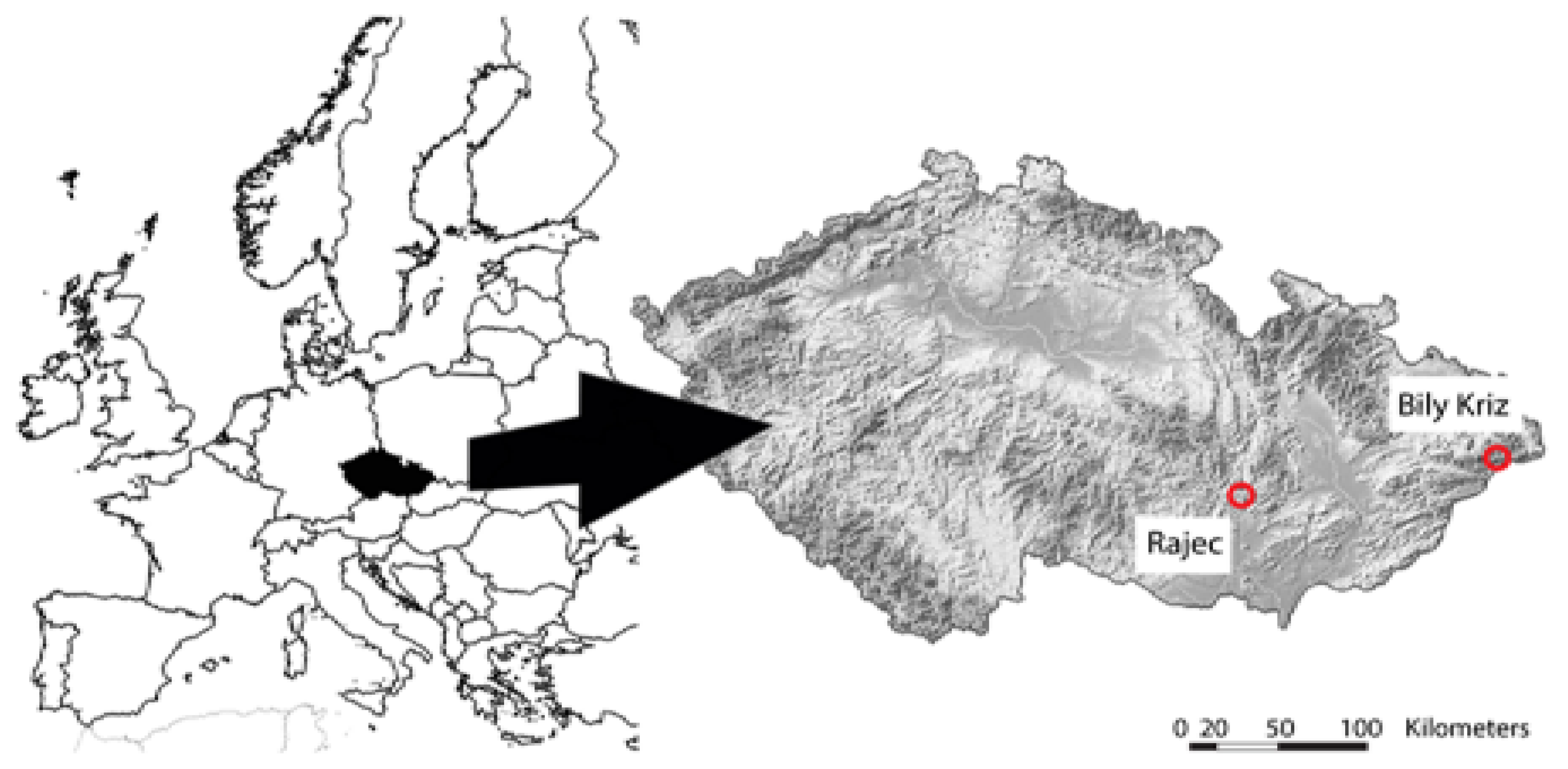
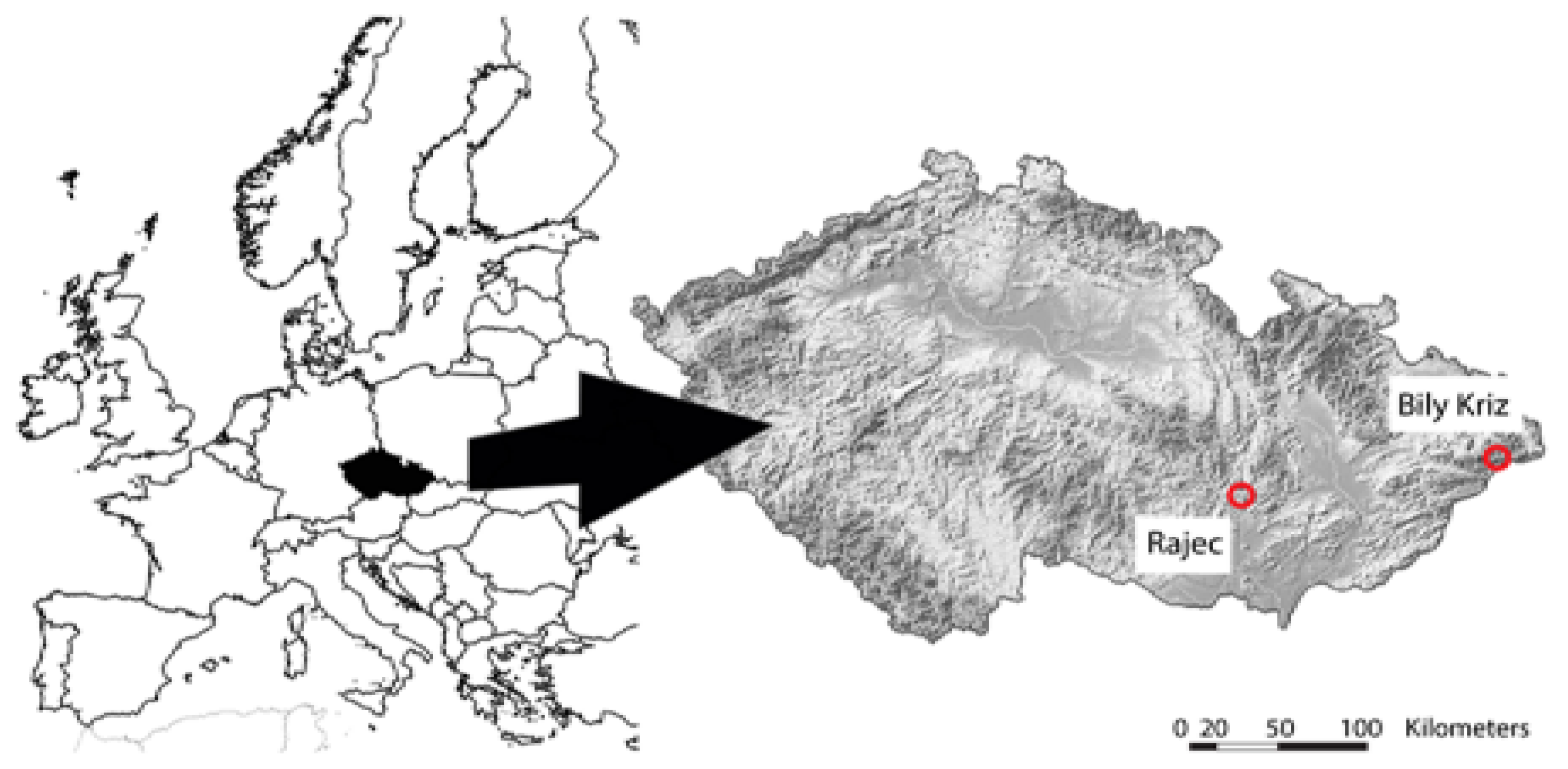
2.1. Experimental Study Plots
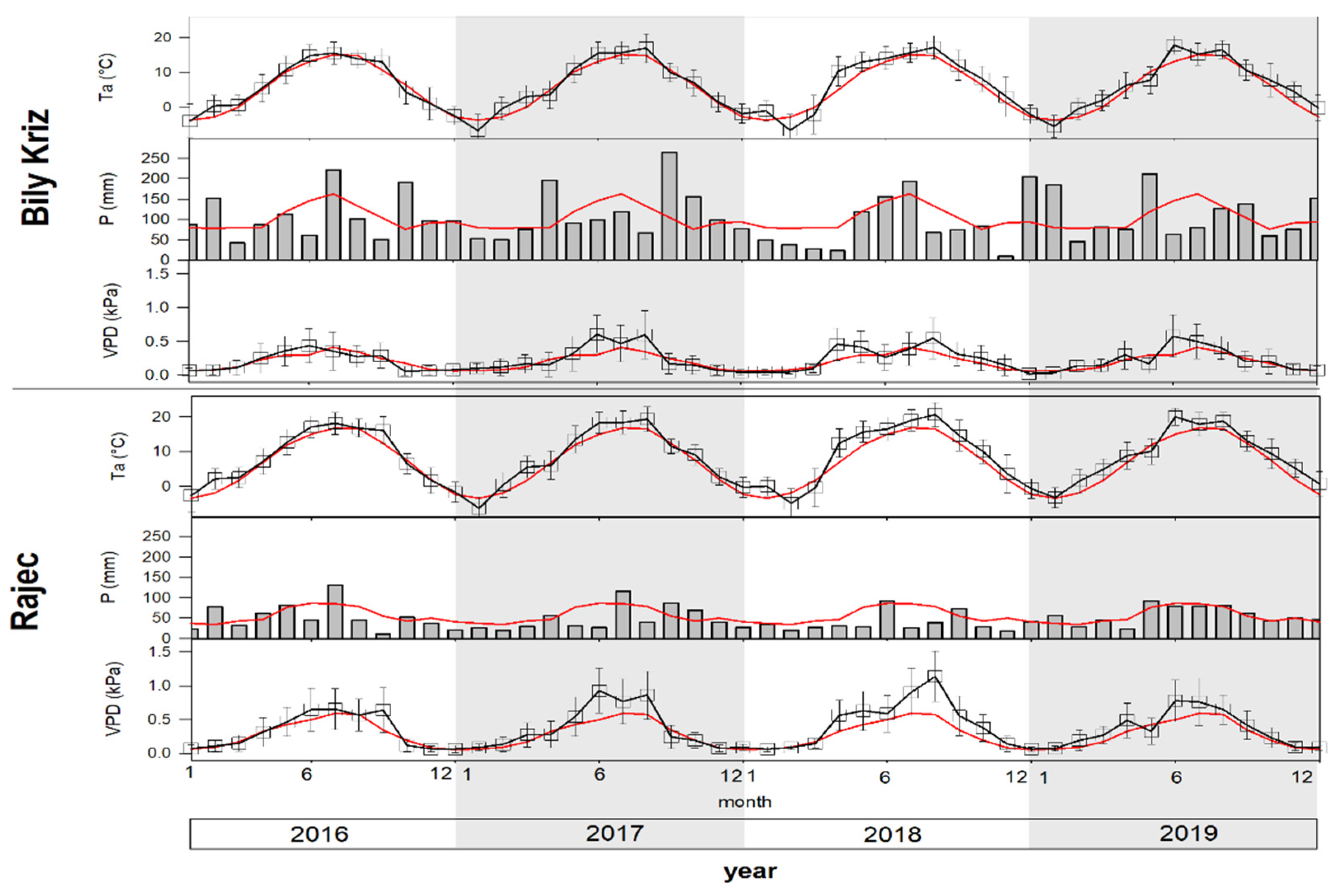
2.2. Micrometeorological Conditions
2.3. Primary Growth
2.4. Secondary Growth
2.5. Statistical Analysis
3. Results
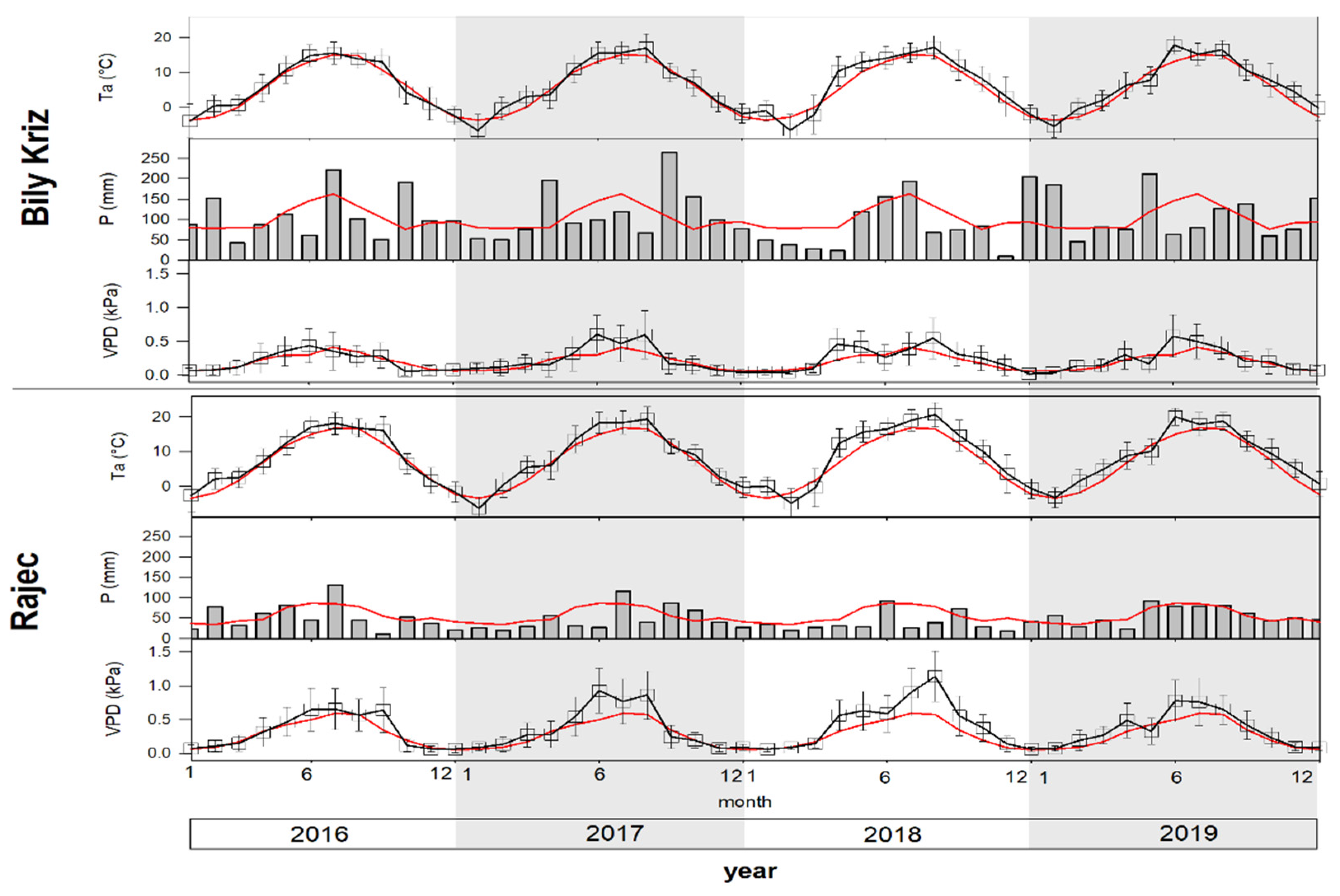
3.1. Variation of Climatic Conditions and GS Prolongation
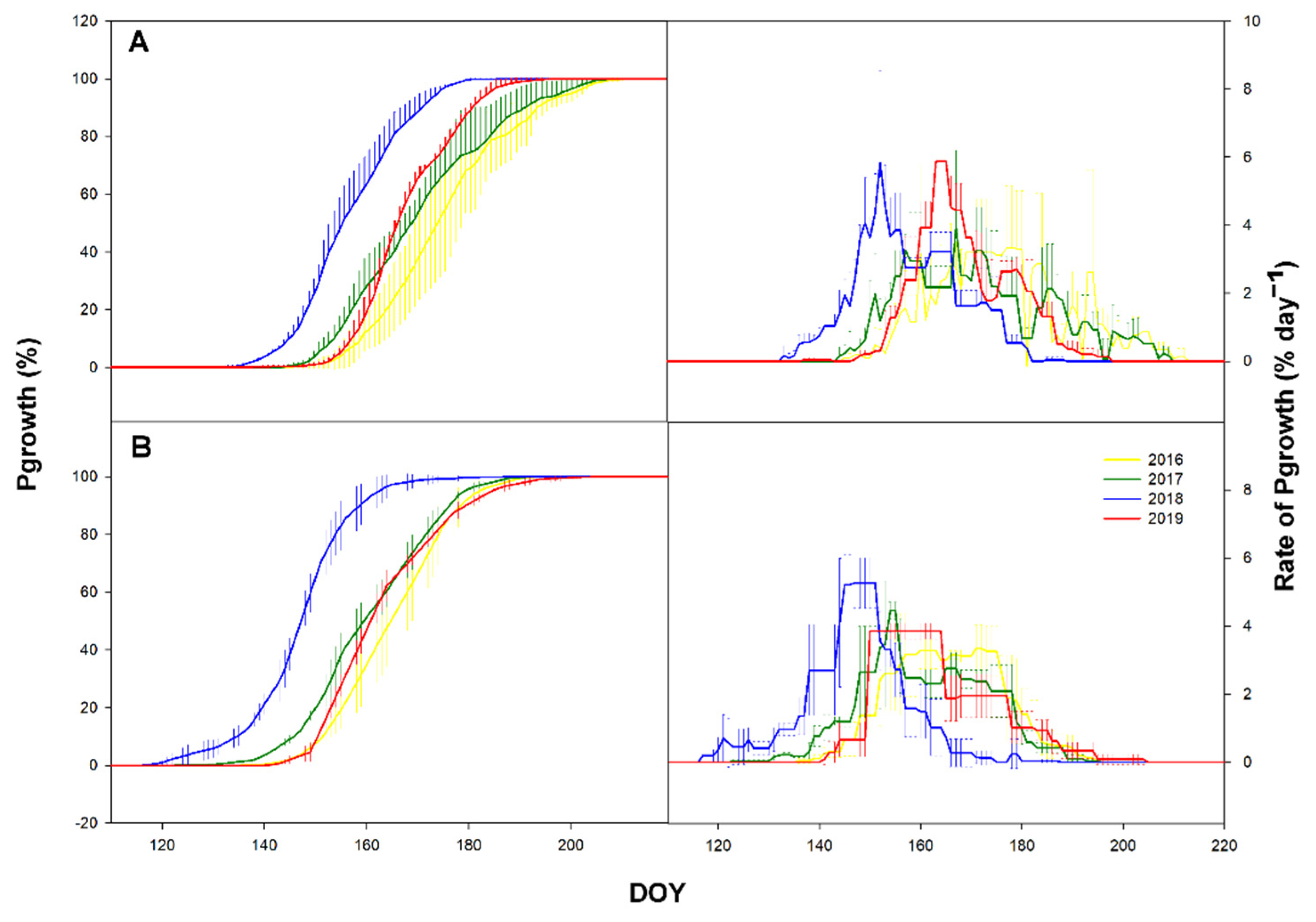
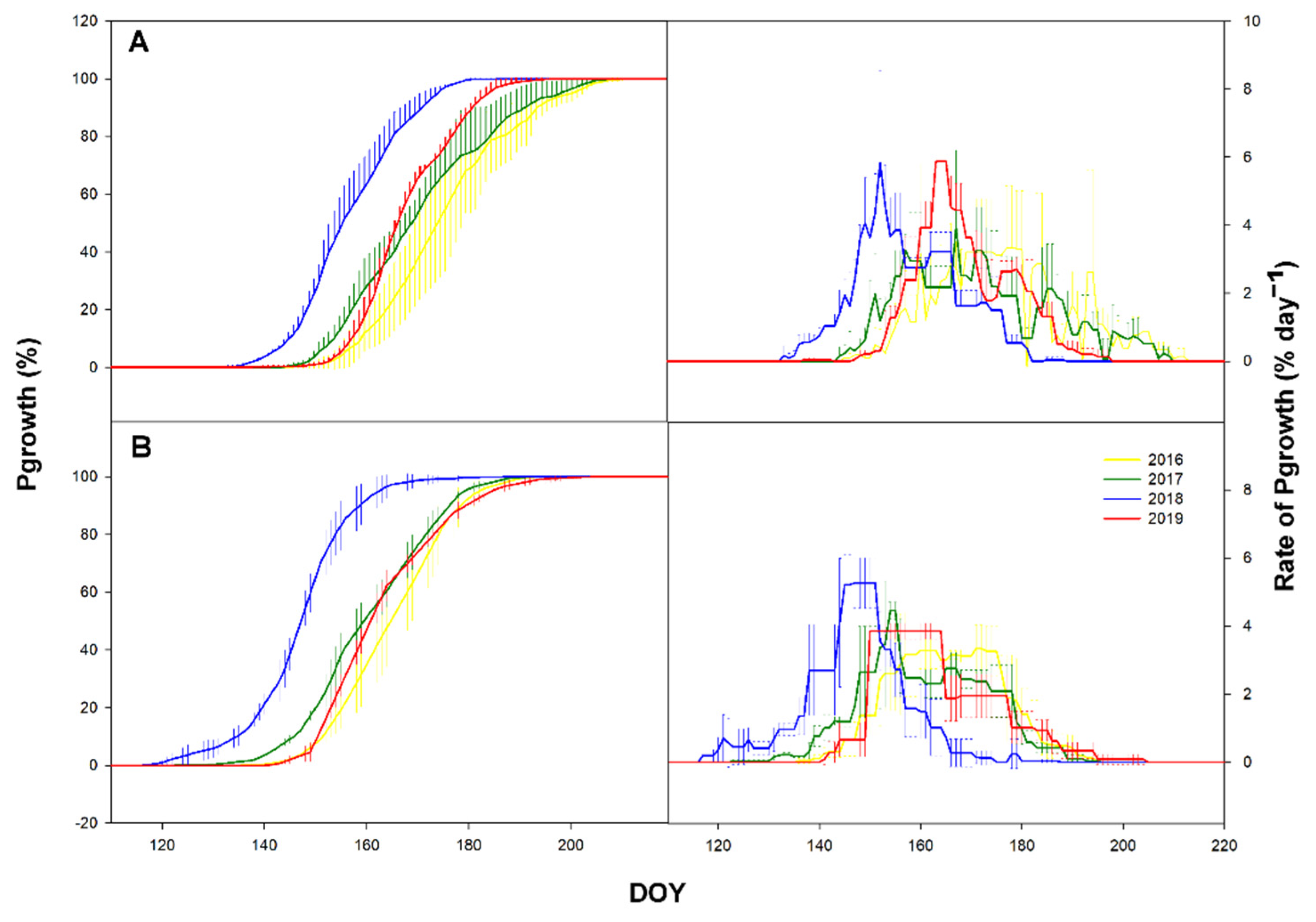
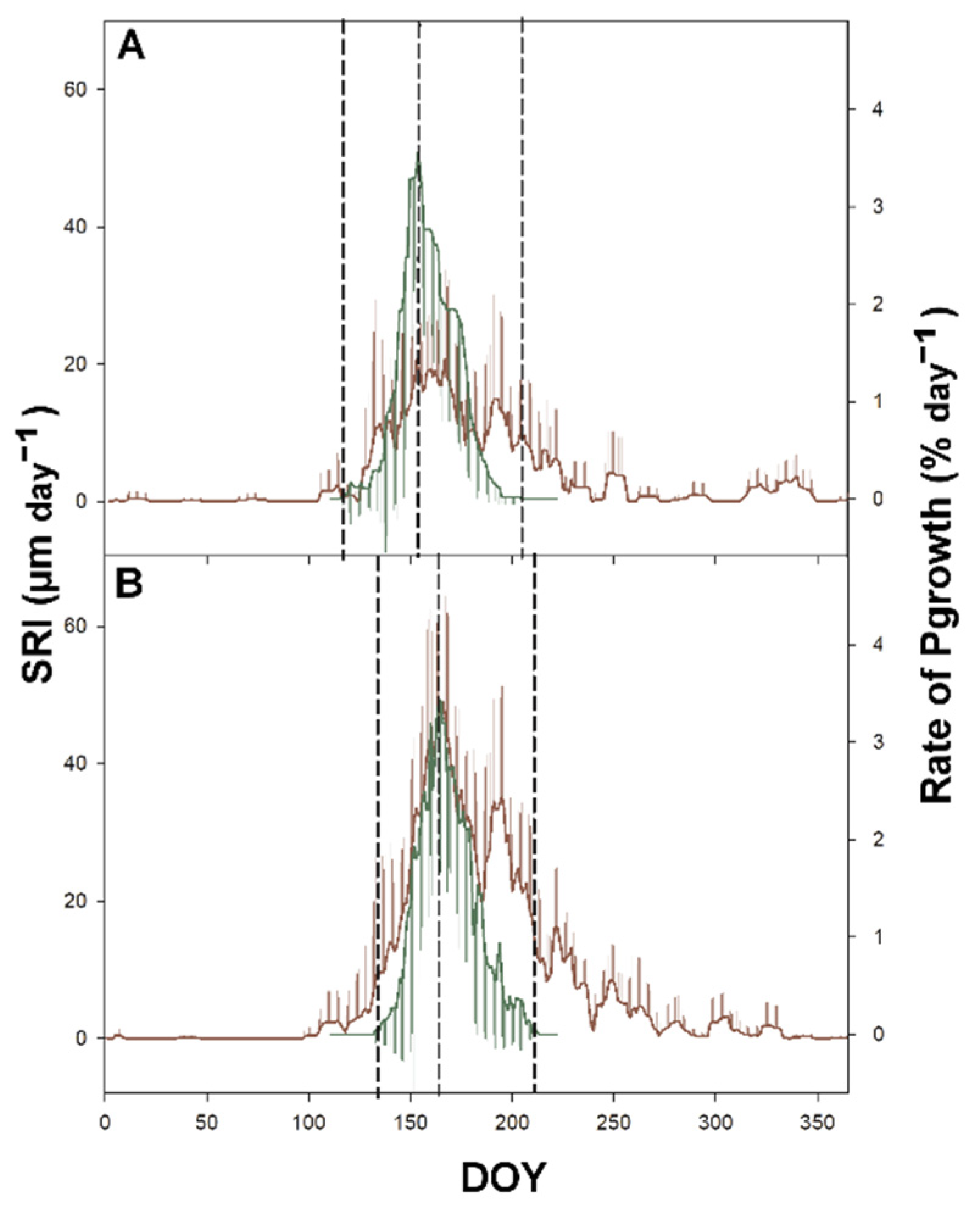
3.2. Primary Growth
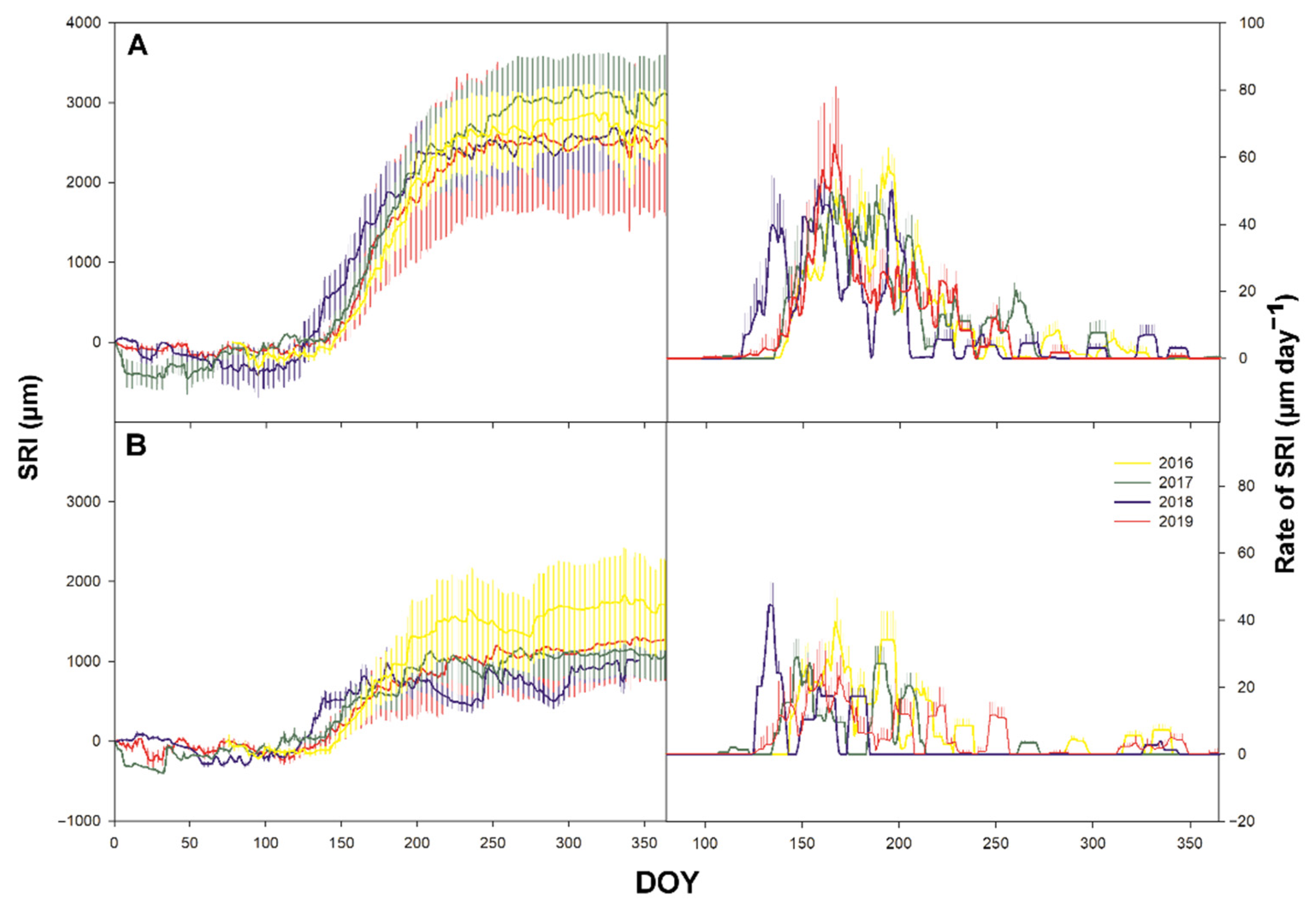
3.3. Secondary Growth
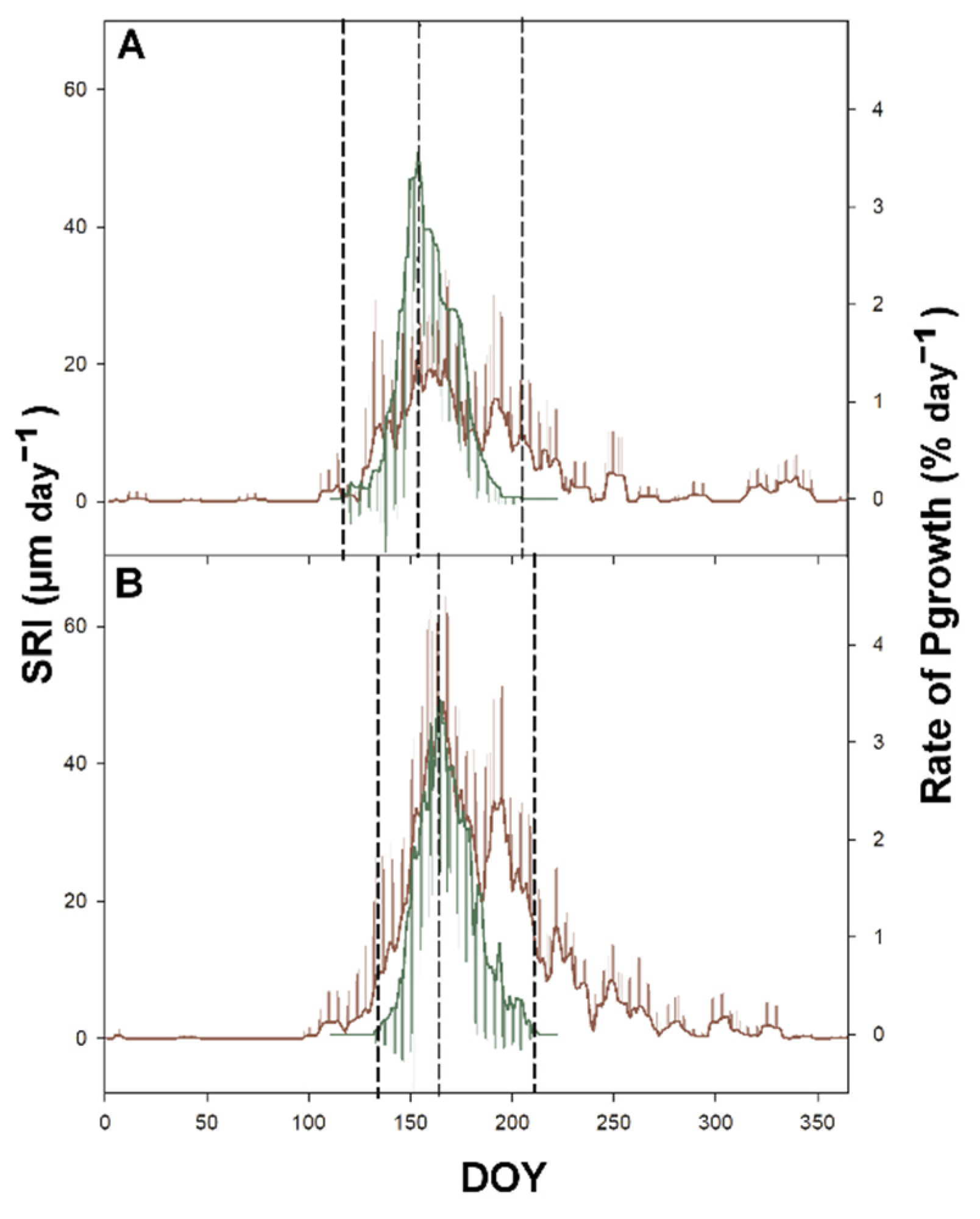
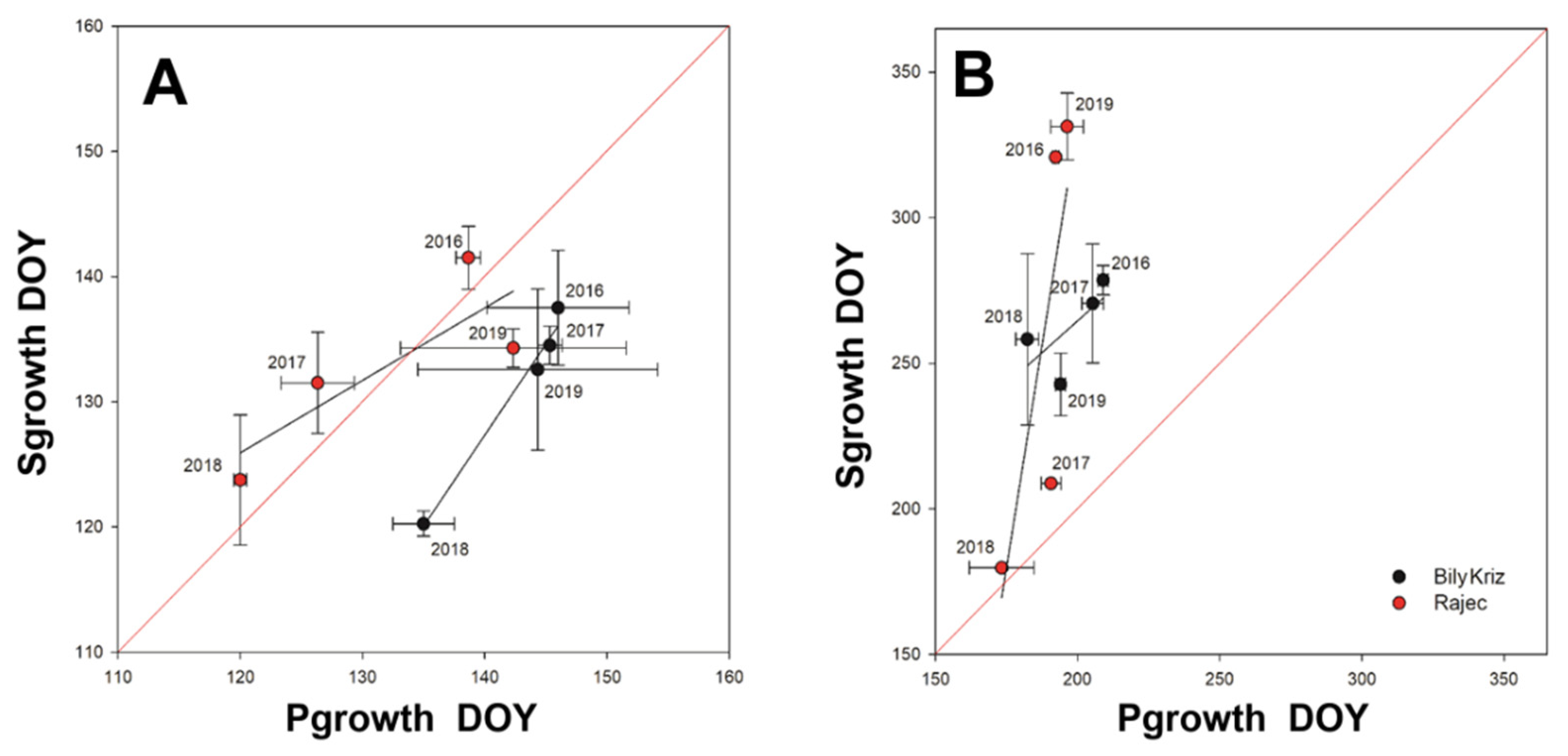
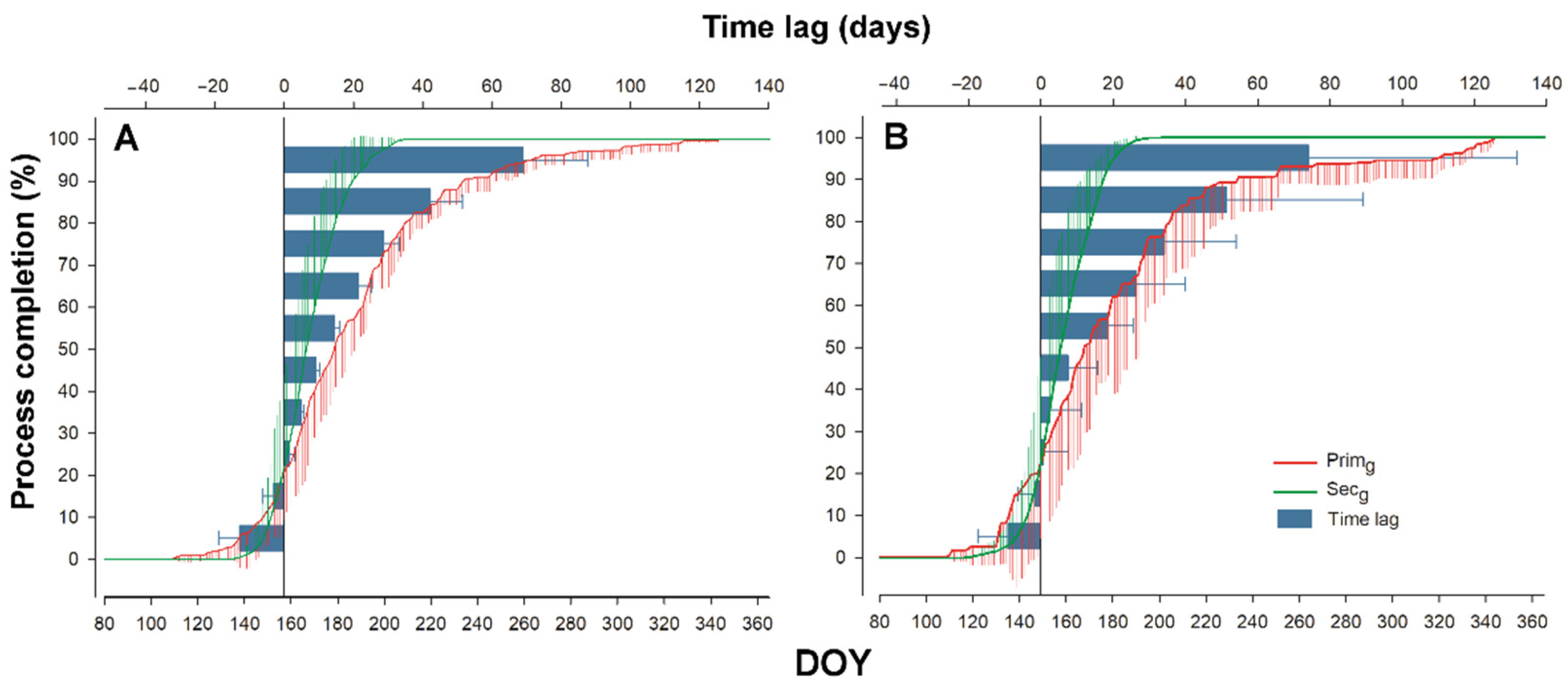
3.4. Time-Lag
4. Discussion
4.1. Variation of Climatic Conditions and GS Prolongation
4.2. Sgrowth Started Earlier Than Pgrowth
4.3. Timing of Pgrowth and Sgrowth Regulated by Different Triggers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bui, M.; Adjiman, C.; Bardow, A.; Boston, A.; Brown, S.; Fennell, P.; Fuss, S. Carbon capture and storage: The way forward. Energy Environ. Sci. 2018, 11, 1062–1176. [Google Scholar] [CrossRef] [Green Version]
- Ciais, P.; Schelhaas, M.; Zaehle, S.; Piao, S.L.; Cescatti, A.; Liski, J.; Luyssaert, S.; Le-Maire, G.; Schulze, E.D.; Bouriaud, O.; et al. Carbon accumulation in European forests. Nat. Geosci. 2008, 1, 425–429. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W.; Rigo, D. Picea Abies in Europe: Distribution, Habitat, Usage and Threats. European Atlas of Forest Tree Species, 1st ed.; Publication Office of the European Union: Luxembourg, 2016; pp. 114–116. [Google Scholar]
- Klimo, E.; Kulhavy, J. Norway spruce monocultures and their transformation to close-to-nature forests from the point of view of soil changes in the Czech Republic. Ekológia 2006, 25, 27–43. [Google Scholar]
- Lindner, M.; Gonzalo, J.G.; Kolström, M.; Green, T. Impacts of Climate Change on European Forests and Options for Adaptation; European Forestry Institute: Boku, Vienna, 2008; pp. 1–173. [Google Scholar]
- Pasqui, M.; Giuseppe, D.E. Climate change, future warming, and adaptation in Europe. Anim. Front. 2019, 9, 6–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidl, R.; Rammer, W.; Jäger, D.; Lexer, M.J. Impact of bark beetle (Ips typographus L.) disturbance on timber production and carbon sequestration in different management strategies under climate change. For. Ecol. Manag. 2008, 256, 209–220. [Google Scholar] [CrossRef]
- Piao, S.; Wang, X.; Ciais, P.; Zhu, B.; Wang, T.; Liu, J. Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Glob. Chang. Biol. 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- Allen, C.; Macalady, A.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.; Hogg, E.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Reyer, C.P.; Leuzinger, S.; Rammig, A.; Wolf, A.; Bartholomeus, R.P.; Bonfante, A.; de Lorenzi, F.; Dury, M.; Gloning, P.; Abou Jaoudé, R.; et al. A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability. Glob. Chang. Biol. 2013, 19, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999, 397, 659. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Rötzer, T. Response of tree phenology to climate change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B.K.; Frank, D.; Fonti, P.; Mäkinen, H.; Prislan, P.; Rossi, S.; del Castillo, E.M.; Campelo, F.; Vavrčík, H. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 2015, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.S.H.; Zhao, H.F.; Piao, S.L.; Peaucelle, M.; Peng, S.S.; Zhou, G.Y.; Ciais, P.; Huang, M.T.; Menzel, A.; Uelas, J.P. Declining global warming effects on the phenology of spring leaf unfolding. Nature 2015, 526, 104–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuine, I.; Beaubien, E.G. Phenology is a major determinant of tree species range. Ecol. Lett. 2001, 4, 500–510. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.G.; Bergeron, Y.; Zhai, L.H.; Denneler, B. Variation in intra-annual radial growth (xylem formation) of Picea mariana (pinaceae) along a latitudinal gradient in western Quebec, Canada. Am. J. Bot. 2011, 98, 792–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Penuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis ofJuniperus przewalskiiin a semi-arid area of the north-eastern Tibetan. Plateau Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef]
- Antonucci, S.; Rossi, S.; Deslauriers, A.; Lombardi, F.; Marchetti, M.; Tognetti, R. Synchronisms and correlations of spring phenology between apical and lateral meristems in two boreal conifers. Trees Struct. Funct. 2015, 35, 1086–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Zhang, Y.; Guo, Y.; Kang, M.; Wang, M.; Wang, B. Intra-annual xylem growth of Larixprincipis-rupprechtii at its upper and lower distribution limits on the Luyashan mountain in north-central China. Forests 2015, 6, 3809–3827. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Hollinger, D.Y. A method to estimate the additional uncertainty in gap-filled NEE resulting from long gaps in the CO2 flux record. Agric. For. Meteorol. 2007, 147, 199–208. [Google Scholar] [CrossRef]
- Soudani, K.G.; Hmimina, N.; Delpierre, J.; Pontailler, M.; Aubinet, D.; Bonal, B.; Caquet, A.; Grandcourt, B.; Burban, C.; Flechard, D.; et al. Ground-based Network of NDVI measurements for tracking temporal dynamics of canopy structure and vegetation phenology in different biomes. Remote Sens. Environ. 2012, 123, 234–245. [Google Scholar] [CrossRef]
- Sonnentag, O.; Hufkens, K.; Teshera-Sterne, C.; Young, A.M.; Friedl, M.; Braswell, B.H.; Milliman, T.; O’Keefe, J.; Richardson, A.D. Digital repeat photography for phenological research in forest ecosystems. Agric. For. Meteorol. 2012, 152, 159–177. [Google Scholar] [CrossRef]
- Brown, T.B.; Hultine, K.R.; Steltzer, H.; Denny, E.G.; Denslow, M.W.; Granados, J.; Henderson, S.; Richardson, A.D. Using phenocams to monitor our changing earth: Toward a global phenocam network. Front. Ecol. Environ. 2016, 14, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Drew, D.M.; Downes, G. The use of precision dendrometers in research on daily stem size and wood property variation: A review. Dendrochronologia 2009, 27, 159–172. [Google Scholar] [CrossRef]
- Zweifel, R.; Haeni, M.; Buchmann, N.; Eugster, W. Are trees able to grow in periods of stem shrinkage? New Phytol. 2016, 211, 839–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocozza, C.; Palombo, C.; Tognetti, R.; Porta, N.; Anichini, M.; Giovannelli, A.; Emiliani, G. Monitoring intra-annual dynamics of wood formation with microcores and dendrometers in Picea abies at two different altitudes. Tree Physiol. 2016, 36, 832–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, L.; Zweifel, R.; Kahmen, A. Daily stem diameter variations can predict the canopy water status of mature temperate trees. Tree Physiol. 2018, 38, 941–952. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, Y.; Wen, Y.; Ding, X.; Wang, B.; Xu, J. Comparing Primary and Secondary Growth of Co-Occurring Deciduous and Evergreen Conifers in an Alpine Habitat. Forests 2019, 10, 574. [Google Scholar] [CrossRef] [Green Version]
- Quitt, E. Climatic Regions of Czechoslovakia, 1st ed.; Studia Geographica: Brno, Czech Republic, 1971; Volume 16, p. 73. [Google Scholar]
- Pokorný, R.; Rajsnerová, P.; Kubásek, J.; Marková, I.; Tomášková, I. Effect of early to late wood proportion on Norway spruce biomass. Acta Univ. Agric. Silvic. Mendel. Brun. 2012, 60, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Nezval, O.; Krejza, J.; Světlík, J.; Šigut, L.; Horáček, P. Comparison of traditional ground-based observations and digital remote sensing of phenological transitions in a floodplain forest. Agric. For. Meteorol. 2020, 291, 108079. [Google Scholar] [CrossRef]
- Abramoff, M.D.; Magelhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Mini32 Software v. 10.2.11, EMS Brno, The Czech Republic. 2021. Available online: http://www.emsbrno.cz/p.axd/en/Software.html (accessed on 9 March 2021).
- SigmaPlot v. 11.0. Systat Software, San Jose, USA. 2021. Available online: http://www.sigmaplot.co.uk/products/sigmaplot/ (accessed on 9 March 2021).
- Körner, C.; Basler, D. Phenology under global warming. Science 2010, 327, 1461–1462. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.G.; Ma, Q.; Rossi, S.; Biondi, F.; Deslauriers, A.; Fonti, P.; Ziaco, E. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers. Proc. Natl. Acad. Sci. USA 2020, 117, 20645–20652. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.; Vince-Prue, D. Photoperiodism in Plants, 2nd ed.; Academic Press: London, UK, 1996; pp. 1–428. [Google Scholar]
- Olsen, J.E. Light and temperature sensing and signaling in induction of bud dormancy in woody plants. Plant Mol. Biol. 2010, 73, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Tanino, K.K.; Kalcsits, L.; Silim, S.; Kendall, E.; Gray, G.R. Temperature-driven plasticity in growth cessation and dormancy development in deciduous woody plants: A working hypothesis suggesting how molecular and cellular function is affected by temperature during dormancy induction. Plant Mol. Biol. 2010, 73, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.E.; Jensen, J.B.; Molmann, J.A.; Ernstsen, A.; Junttila, O. Photoperiodic Regulation of Apical Growth Cessation in Northern Tree Species: The Role of Phytochrome and Gibberellin. J. Crop. Improv. 2004, 10, 77–112. [Google Scholar] [CrossRef]
- Krejza, J.; Cienciala, E.; Světlík, J.; Bellan, M.; Noyer, E.; Horáček, P.; Marek, M.V. Evidence of climate-induced stress of Norway spruce along elevation gradient preceding the current dieback in Central Europe. Trees 2020, 35, 103–119. [Google Scholar] [CrossRef]
- Chenkova, N.; Nikolova, N. Air temperature and precipitation variability in northeastern Bulgaria on the background of climate change. Therm. Sci. 2015, 19, 381–390. [Google Scholar] [CrossRef]
- Mäkinen, H.; Nöjd, P.; Kahle, H.P.; Neumann, U.; Tveite, B.; Mielikäinen, K.; Röhle, H.; Spiecker, H. Radial growth variation of Norway spruce (Picea abies (L.) Karst.) across latitudinal and altitudinal gradients in central and northern Europe. For. Ecol. Manag. 2002, 171, 243–259. [Google Scholar] [CrossRef]
- Neuwirth, B.; Esper, J.; Schweingruber, F.H.; Winiger, M. Site ecological differences to the climatic forcing of spruce pointer years from the Lötschental, Switzerland. Dendrochronologia 2004, 21, 69–78. [Google Scholar] [CrossRef]
- Büntgen, U.; Frank, D.C.; Kaczka, R.J.; Verstege, A.; Zwijacz, K.T.; Esper, J. Growth responses to climate in a multi-species tree-ring network in the Western Carpathian Tatra Mountains, Poland and Slovakia. Tree Physiol. 2007, 27, 689–702. [Google Scholar] [CrossRef] [Green Version]
- Affolter, P.; Büntgen, U.; Esper, J.; Rigling, A.; Weber, P.; Luterbacher, J.; Frank, D. Inner Alpine conifer response to 20th century drought swings. Eur. J. For. Res. 2010, 129, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Tolasz, R.; Míková, T.; Voženílek, V. Climate Atlas of Czechia, 1st ed.; Czech Hydrometeorological Institute: Prague-Olomouc, Czech Republic, 2007; pp. 1–245. [Google Scholar]
- Liu, Q.; Piao, S.; Janssens, I.M.; Fu, Y.; Peng, S.; Lian, X.; Ciais, P.; Myneni, R.B.; Penuelas, J.; Wang, T. Extension of the growing season increases vegetation exposure to frost. Nat. Commun. 2018, 9, 426. [Google Scholar] [CrossRef] [Green Version]
- Lenz, A.; Hoch, G.; Körner, C.; Vitasse, Y. Convergence of leaf-out towards minimum risk of freezing damage in temperate trees. Funct. Ecol. 2016, 30, 1480–1490. [Google Scholar] [CrossRef]
- Klein, T.; Zeppel, M.J.B.; Anderegg, W.R.L.; Bloemen, J.; Kauwe, M.G.; Hudson, P.; Nardini, A. Xylem embolism refilling and resilience against drought-induced mortality in woody plants: Processes and trade-offs. Ecol. Res. 2018, 33, 839–855. [Google Scholar] [CrossRef]
- Mencuccini, M.; Salmon, Y.; Mitchell, P.; Hölttä, T.; Choat, B.; Meir, P.; Pfautsch, S. An empirical method that separates irreversible stem radial growth from bark water content changes in trees: Theory and case studies. Plant Cell Environ. 2017, 40, 290–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zweifel, R.; Zimmermann, L.; Zeugin, F.; Newbery, D.M. Intra-annual radial growth and water relations of trees: Implications towards a growth mechanism. J. Exp. Bot. 2006, 57, 1445–1459. [Google Scholar] [CrossRef] [Green Version]
- Mäkinen, H.; Seo, J.-W.; Nöjd, P.; Schmitt, U.; Jalkanen, R. Seasonal dynamics of wood formation: A comparison between pinning, microcoring and dendrometer measurements. Eur. J. For. Res. 2008, 127, 235–245. [Google Scholar] [CrossRef]
- Cruz-García, R.; Balzano, A.; Čufar, K.; Scharnweber, T.; Smiljanić, M.; Wilmking, M. Combining Dendrometer Series and Xylogenesis Imagery—DevX, a Simple Visualization Tool to Explore Plant Secondary Growth Phenology. Front. For. Glob. Chang. 2019, 2, 60. [Google Scholar] [CrossRef]
- Stangler, D.F.; Kahle, H.-P.; Raden, M.; Larysch, E.; Seifert, T.; Spiecker, H. Effects of Intra-Seasonal Drought on Kinetics of Tracheid Differentiation and Seasonal Growth Dynamics of Norway Spruce along an Elevational Gradient. Forests 2021, 12, 274. [Google Scholar] [CrossRef]
- Zhang, S.; Butto, V.; Khare, S.; Deslauriers, A.; Morin, H.; Huang, J.G. Calibrating PhenoCam Data with Phenological Observations of a Black Spruce Stand. Can. J. Remote Sens. 2020, 46, 154–165. [Google Scholar] [CrossRef]
- Blauhut, V.; Stahl, K.; Stagge, J.H.; Tallaksen, L.M.; Stefano, L.; Vogt, J. Estimating drought risk across Europe from reported drought impacts, drought indices, and vulnerability factors. Hydrol. Earth Syst. Sci. 2016, 20, 2779–2800. [Google Scholar] [CrossRef] [Green Version]
- Lisar, Y.S.L.; Agdam, H.B. Drought Stress in Plants: Causes, Consequences, and Tolerance, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–16. [Google Scholar]
- Gricar, J.; Zupancic, M.; Cufar, K.; Oven, P. Regular cambial activity and xylem and phloem formation in locally heated and cooled stem portions of Norway spruce. Wood Sci. Technol. 2007, 41, 463–475. [Google Scholar] [CrossRef]
- Fajstavr, M.; Giagli, K.; Vavrčík, H.; Gryc, V.; Urban, J. The effect of stem girdling on xylem and phloem formation in Scots pine. Silva Fenn. 2017, 51, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Zhang, A. Phosphorous fertilization alleviates drought effects on Alnus cremastogyne by regulating its antioxidant and osmotic potential. Sci. Rep. 2018, 8, 5644. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Eugster, W.; Ammann, C.; Häni, M.; Zielis, S.; Hiller, R.; Buchmann, N. Contrasting response of grassland versus forest carbon and water fluxes to spring drought in Switzerland. Environ. Res. Lett. 2013, 8, 35007. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. Paradigm shift in plant growth control. Curr. Opin. Plant Biol. 2015, 25, 107–114. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Rajec | Bily Kriz | ||||||
|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | 2019 | 2016 | 2017 | 2018 | 2019 | |
| Ta (°C) | 8.1 | 8.2 | 9.0 | 9.0 | 6.1 | 6.3 | 6.9 | 6.9 |
| Tadev. (°C) | 1.1 | 1.2 | 2.0 | 2.0 | 0.5 | 0.7 | 1.3 | 1.3 |
| P (mm) | 618.6 | 568.8 | 456.3 | 685.7 | 1298.6 | 1346.6 | 1045.2 | 1289.8 |
| Pdev. (%) | 93.3 | 85.8 | 68.8 | 103.4 | 106.7 | 110.6 | 85.9 | 105.9 |
| Parameter | Items | df | MS | F | p | |
|---|---|---|---|---|---|---|
| OnsetSGrowth | Locality | 1 | 21.63 | 0.52 | 0.48 | |
| Year | 3 | 428.30 | 10.36 | <0.001 | ||
| Residuals | 30 | 41.36 | ||||
| EndSGrowth | Locality | 1 | 44.64 | 0.25 | 0.62 | |
| Year | 3 | 12805.67 | 71.16 | <0.001 | ||
| Residuals | 30 | 179.95 | ||||
| DurationSGrowth | Locality | 1 | 608.52 | 5.54 | 0.03 | |
| Year | 3 | 8690.59 | 79.15 | <0.001 | ||
| Residuals | 30 | 109.81 | ||||
| OnsetPGrowth | Locality | 1 | 704.17 | 47.88 | <0.001 | |
| Year | 3 | 319.83 | 21.75 | <0.001 | ||
| Residuals | 16 | 14.71 | ||||
| EndPGrowth | Locality | 1 | 541.50 | 20.34 | <0.001 | |
| Year | 3 | 636.94 | 23.92 | <0.001 | ||
| Residuals | 16 | 26.63 | ||||
| DurationPGrowth | Locality | 1 | 10.67 | 0.22 | 0.64 | |
| Year | 3 | 185.00 | 3.84 | 0.03 | ||
| Residuals | 16 | 48.21 | ||||
| Onset | End | Duration | |||||
|---|---|---|---|---|---|---|---|
| Mean (DOY) | SD | Mean (DOY) | SD | Mean (DOY) | SD | ||
| Sgrowth | Bily Kriz | 131.21 | 8.80 | 259.37 | 21.75 | 115.11 | 21.59 |
| Rajec | 132.76 | 8.14 | 271.47 | 68.66 | 138.42 | 63.75 | |
| Pgrowth | Bily Kriz | 142.67 | 5.80 | 197.67 | 11.20 | 55.00 | 8.40 |
| Rajec | 131.83 | 9.93 | 188.17 | 10.80 | 56.33 | 8.42 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nezval, O.; Krejza, J.; Bellan, M.; Světlík, J. Asynchrony and Time-Lag between Primary and Secondary Growth of Norway Spruce Growing in Different Elevations. Forests 2021, 12, 627. https://doi.org/10.3390/f12050627
Nezval O, Krejza J, Bellan M, Světlík J. Asynchrony and Time-Lag between Primary and Secondary Growth of Norway Spruce Growing in Different Elevations. Forests. 2021; 12(5):627. https://doi.org/10.3390/f12050627
Chicago/Turabian StyleNezval, Ondřej, Jan Krejza, Michal Bellan, and Jan Světlík. 2021. "Asynchrony and Time-Lag between Primary and Secondary Growth of Norway Spruce Growing in Different Elevations" Forests 12, no. 5: 627. https://doi.org/10.3390/f12050627
APA StyleNezval, O., Krejza, J., Bellan, M., & Světlík, J. (2021). Asynchrony and Time-Lag between Primary and Secondary Growth of Norway Spruce Growing in Different Elevations. Forests, 12(5), 627. https://doi.org/10.3390/f12050627