
Induction of Defense Responses in Pinus sylvestris Seedlings by Methyl Jasmonate and Response to Heterobasidion annosum and Lophodermium seditiosum Inoculation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Exogenous Application of MeJA, Inoculation with H. annosum and L. seditiosum, and Sampling
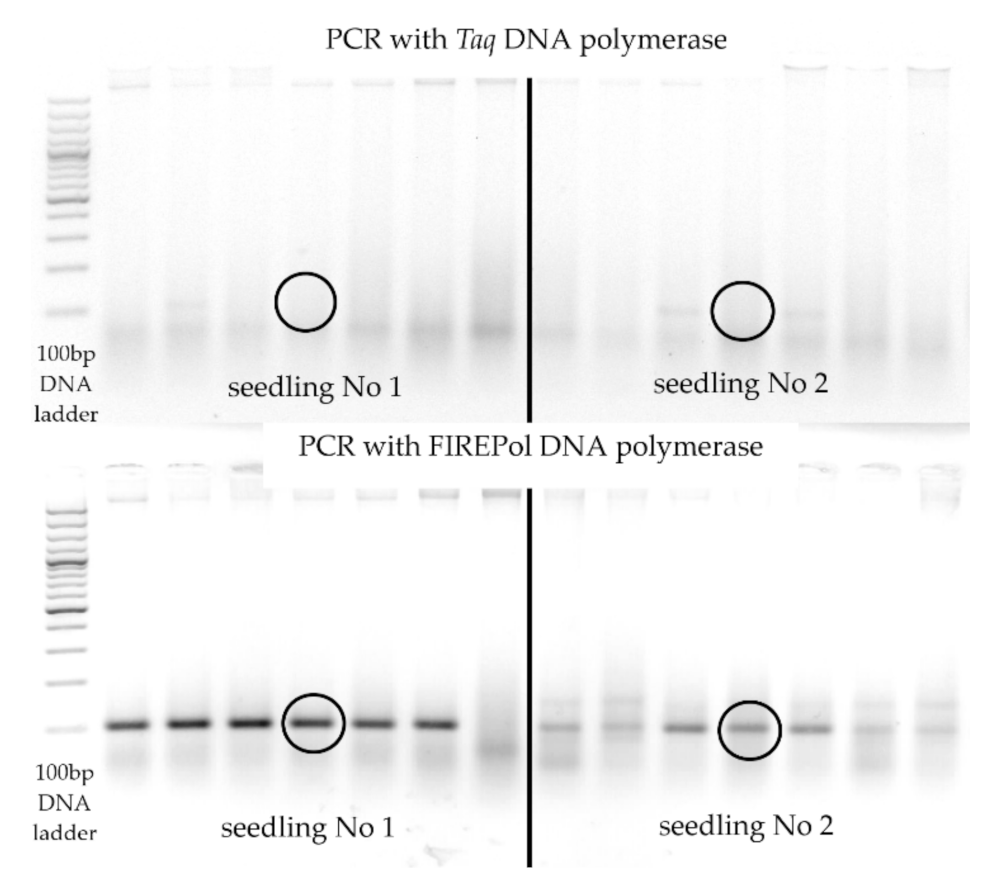
2.2. DNA Extraction, PCR Conditions
2.3. RNA Extraction and Real-Time PCR
3. Results
3.1. Experiment 1: Clonal Ramet Treatment with MeJA, Inoculation with H. annosum, and TLP Gene Expression
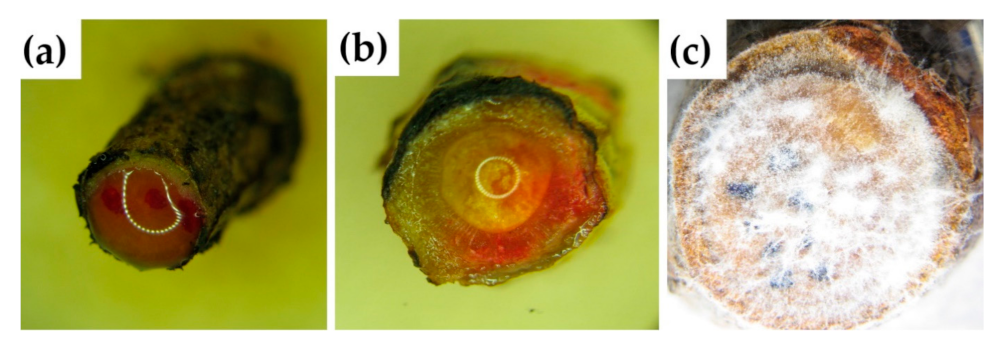
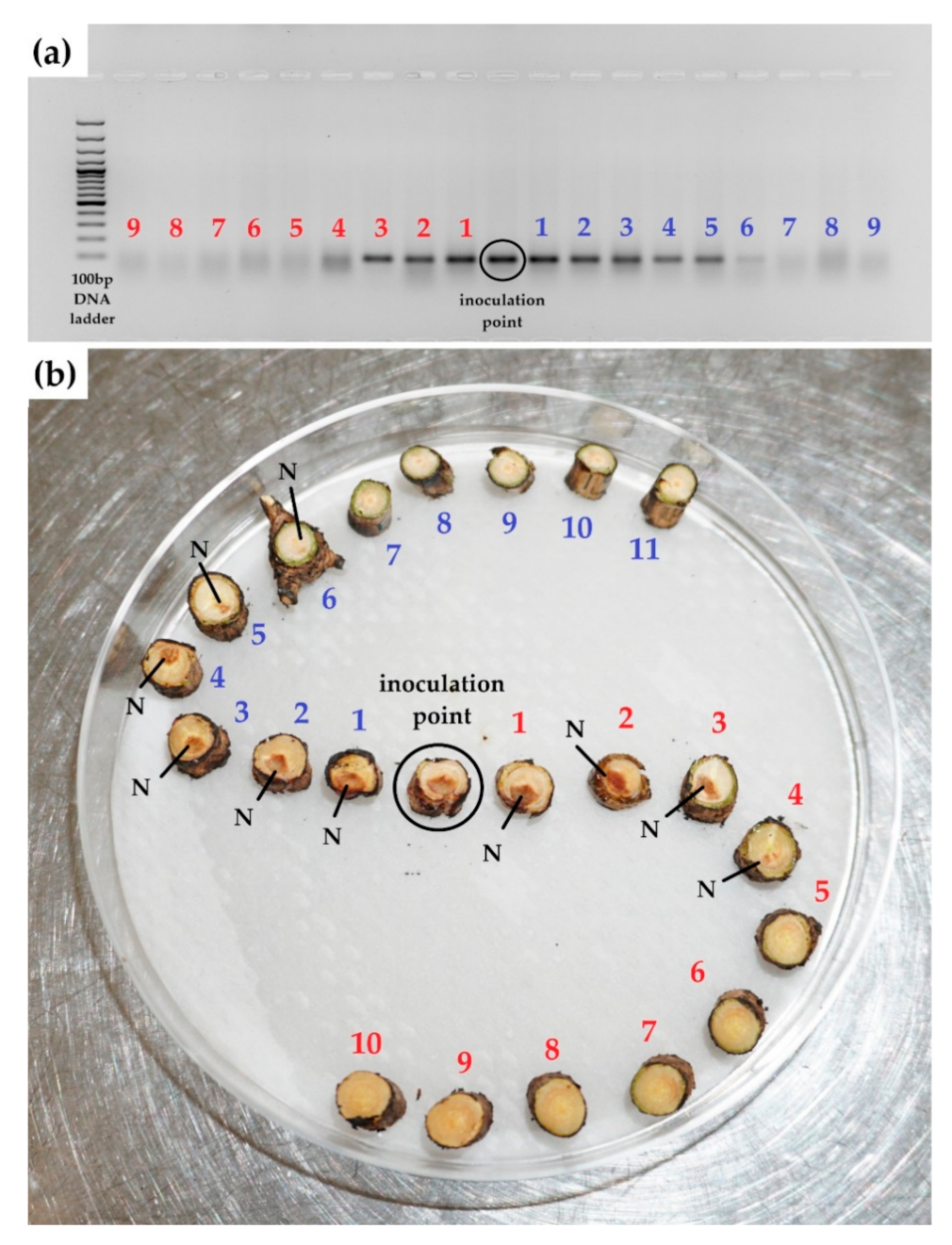
3.2. Experiment 2: P. sylvestris Seedling Treatment with MeJA, PCR Results of Inoculation with H. annosum
3.3. Experiment 3: Seedling Treatment with MeJA and Inoculation with L. seditiosum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heijari, J.; Nerg, A.M.; Kainulainen, P.; Viiri, H.; Vuorinen, M.; Holopainen, J.K. Application of methyl jasmonate reduces growth but increases chemical defence and resistance against Hylobius abietis in Scots pine seedlings. Entomol. Exp. Appl. 2005, 115, 117–124. [Google Scholar] [CrossRef]
- Martin, D.; Tholl, D.; Gershenzon, J.; Bohlmann, J. Methyl Jasmonate Induces Traumatic Resin Ducts, Terpenoid Resin Biosynthesis, and Terpenoid Accumulation in Developing Xylem of Norway Spruce Stems. Plant Physiol. 2002, 129, 1003–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudgins, J.W.; Christiansen, E.; Franceschi, V.R. Induction of anatomically based defense responses in stems of diverse conifers by methyl jasmonate: A phylogenetic perspective. Tree Physiol. 2004, 24, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, X.; Sampedro, L.; Zas, R. Defensive responses of Pinus pinaster seedlings to exogenous application of methyl jasmonate: Concentration effect and systemic response. Environ. Exp. Bot. 2009, 67, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Heijari, J.; Nerg, A.-M.; Kainulainen, P.; Vuorinen, M.; Holopainen, J.K. Long-term effects of exogenous methyl jasmonate application on Scots pine (Pinus sylvestris) needle chemical defence and diprionid sawfly performance. Entomol. Exp. Appl. 2008, 128, 162–171. [Google Scholar] [CrossRef]
- Kozlowski, G.; Buchala, A.; Métraux, J.-P. Methyl jasmonate protects Norway spruce [Picea abies (L.) Karst.] seedlings against Pythium ultimum Trow. Physiol. Mol. Plant Pathol. 1999, 55, 53–58. [Google Scholar] [CrossRef]
- Krokene, P.; Nagy, N.E.; Solheim, H. Methyl jasmonate and oxalic acid treatment of Norway spruce: Anatomically based defense responses and increased resistance against fungal infection. Tree Physiol. 2008, 28, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-J.; Zamani, A.; Ekramoddoullah, A.K.M. Expression profiling of a complex thaumatin-like protein family in western white pine. Planta 2009, 231, 637–651. [Google Scholar] [CrossRef]
- Singh, N.K.; Bracker, C.A.; Hasegawa, P.M.; Handa, A.K.; Buckel, S.; Hermodson, M.A.; Pfankoch, E.; Regnier, F.E.; Bressan, R.A. Characterization of Osmotin. A thaumatin-like protein associated with osmotic adaptation in plant cells. Plant Physiol. 1987, 85, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.K.; Nelson, D.E.; Kuhn, D.; Hasegawa, P.M.; Bressan, R.A. Molecular Cloning of Osmotin and Regulation of Its Expression by ABA and Adaptation to Low Water Potential. Plant Physiol. 1989, 90, 1096–1101. [Google Scholar] [CrossRef] [Green Version]
- Neale, A.D.; Wahleithner, J.A.; Lund, M.; Bonnett, H.T.; Kelly, A.; Meeks-Wagner, D.R.; Peacock, W.J.; Dennis, E.S. Chitinase, beta-1,3-glucanase, osmotin, and extensin are expressed in tobacco explants during flower formation. Plant Cell 1990, 2, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koiwa, H.; Sato, F.; Yamada, Y. Characterization of Accumulation of Tobacco PR-5 Proteins by IEF-Immunoblot Analysis. Plant Cell Physiol. 1994, 35, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Reymond, P.; Farmer, E.E. Jasmonate and salicylate as global signals for defense gene expression. Curr. Opin. Plant Biol. 1998, 1, 404–411. [Google Scholar] [CrossRef]
- Piggott, N.; Ekramoddoullah, A.K.M.; Liu, J.-J.; Yu, X. Gene cloning of a thaumatin-like (PR-5) protein of western white pine (Pinus monticola D. Don) and expression studies of members of the PR-5 group. Physiol. Mol. Plant Pathol. 2004, 64, 1–8. [Google Scholar] [CrossRef]
- Pervieux, I.; Bourassa, M.; Laurans, F.; Hamelin, R.; Séguin, A. A spruce defensin showing strong antifungal activity and increased transcript accumulation after wounding and jasmonate treatments. Physiol. Mol. Plant Pathol. 2004, 64, 331–341. [Google Scholar] [CrossRef]
- Liu, J.-J.; Ekramoddoullah, A.K.M.; Zamani, A. A Class IV Chitinase Is Up-Regulated by Fungal Infection and Abiotic Stresses and Associated with Slow-Canker-Growth Resistance to Cronartium ribicola in Western White Pine (Pinus monticola). Phytopathology 2005, 95, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ng, T.B. Isolation of an antifungal thaumatin-like protein from kiwi fruits. Phytochemistry 2002, 61, 1–6. [Google Scholar] [CrossRef]
- Krebitz, M.; Wagner, B.; Ferreira, F.; Peterbauer, C.; Campillo, N.; Witty, M.; Kolarich, D.; Steinkellner, H.; Scheiner, O.; Breiteneder, H. Plant-based Heterologous Expression of Mal d 2, a Thaumatin-like Protein and Allergen of Apple (Malus domestica), and its Characterization as an Antifungal Protein. J. Mol. Biol. 2003, 329, 721–730. [Google Scholar] [CrossRef]
- Jayaraj, J.; Velazhahan, R.; Fu, D.; Liang, G.H.; Muthukrishnan, S. Bacterially produced rice thaumatin-like protein shows in vitro antifungal activity. J. Plant Dis. Prot. 2004, 111, 334–344. [Google Scholar]
- Vitali, A.; Pacini, L.; Bordi, E.; de Mori, P.; Pucillo, L.; Maras, B.; Botta, B.; Brancaccio, A.; Giardina, B. Purification and characterization of an antifungal thaumatin-like protein from Cassia didymobotrya cell culture. Plant Physiol. Biochem. 2006, 44, 604–610. [Google Scholar] [CrossRef]
- Mohamed, M.S.; El-Soud, W.A.; Mohamed, M.; Fervat, S.M.; Walid, A.E.S.; Magda, F.M. Cloning and expression of the recombinant NP24I protein from tomato fruit and study of its antimicrobial activity. Afr. J. Biotechnol. 2011, 10, 14276–14285. [Google Scholar] [CrossRef]
- Šķipars, V.; Rauda, E.; Snepste, I.; Krivmane, B.; Rungis, D. Assessment of gene copy number variation of Scots pine thaumatin-like protein gene using real-time PCR based methods. Tree Genet. Genomes 2017, 13, 127. [Google Scholar] [CrossRef]
- Šņepste, I.; Šķipars, V.; Krivmane, B.; Bruna, L.; Ruņģis, D. Characterization of a Pinus sylvestris thaumatin-like protein gene and determination of antimicrobial activity of the in vitro expressed protein. Tree Genet. Genomes 2018, 14, 58. [Google Scholar] [CrossRef]
- Kanberga-Siliņa, K.; Rauda, E.; Šķipars, V.; Vivian-Smith, A.; Yakovlev, I.; Krivmane, B.; Šņepste, I.; Ruņģis, D. Transcriptomic response to methyl jasmonate treatment of Scots pine (Pinus sylvestris) seedlings. Environ. Exp. Biol. 2017, 15, 257–274. [Google Scholar] [CrossRef]
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef] [Green Version]
- Thakur, M.; Sohal, B.S. Role of Elicitors in Inducing Resistance in Plants against Pathogen Infection: A Review. ISRN Biochem. 2013, 2013, 762412. [Google Scholar] [CrossRef] [Green Version]
- Nejat, N.; Mantri, N. Plant Immune System: Crosstalk Between Responses to Biotic and Abiotic Stresses the Missing Link in Understanding Plant Defence. Curr. Issues Mol. Biol. 2017, 23, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Arhipova, N.; Gaitnieks, T.; Donis, J.; Stenlid, J.; Vasaitis, R. Butt rot incidence, causal fungi, and related yield loss in Picea abies stands of Latvia. Can. J. For. Res. 2011, 41, 2337–2345. [Google Scholar] [CrossRef]
- Garbelotto, M.; Gonthier, P. Biology, Epidemiology, and Control of Heterobasidion Species Worldwide. Annu. Rev. Phytopathol. 2013, 51, 39–59. [Google Scholar] [CrossRef] [Green Version]
- Swedjmark, G.; Stenlid, J. Between-tree and between-isolate variation for growth of S-group Heterobasidion annosum in sapwood of Picea abies cuttings. Can. J. For. Res. 1997, 27, 711–715. [Google Scholar] [CrossRef]
- Swedjemark, G.; Karlsson, B. Mycelial growth and exclusion of Heterobasidion parviporum inoculated in branches of 15-year-old Picea abies clones. For. Pathol. 2006, 36, 209–214. [Google Scholar] [CrossRef]
- Skrøppa, T.; Solheim, H.; Steffenrem, A. Genetic variation, inheritance patterns and parent–offspring relationships after artificial inoculations with Heterobasidion parviporum and Ceratosystis polonica in Norway spruce seed orchards and progeny tests. Silva Fenn. 2015, 49, 49. [Google Scholar] [CrossRef] [Green Version]
- Marčiulynas, A.; Sirgedaitė-Šėžienė, V.; Žemaitis, P.; Baliuckas, V. The Resistance of Scots Pine (Pinus sylvestris L.) Half-sib Families to Heterobasidion annosum. Forests 2019, 10, 287. [Google Scholar] [CrossRef] [Green Version]
- Zaluma, A.; Gaitnieks, T.; Arhipova, N.; Vasaitis, R. Growth rates of Heterobasidion annosum ss and H. aparviporum in functional sapwood of Pinus sylvestris and Picea abies. For. Pathol. 2015, 45, 437–439. [Google Scholar] [CrossRef]
- Hanso, M.; Drenkhan, R. Lophodermium needle cast, insect defoliation and growth responses of young Scots pines in Estonia. For. Pathol. 2011, 42, 124–135. [Google Scholar] [CrossRef]
- Kowalski, T. Interrelationships among Lophodermium seditiosum, L. pinastri and Cyclaneusma minus in pine plantations (Pinus sylvestris L.) in Poland. In Proceedings of the Conference: Recent Research on Foliage Diseases, Carlisle, PE, USA, 29 May–2 June 1989; pp. 13–18. [Google Scholar]
- Diwani, S.A.; Millar, C.S. Sources of inoculum of Lophodermium seditiosum on Pinus sylvestris. For. Pathol. 1990, 20, 1–7. [Google Scholar] [CrossRef]
- Ganley, R.J.; Newcombe, G. Fungal endophytes in seeds and needles of Pinus monticola. Mycol. Res. 2006, 110, 318–327. [Google Scholar] [CrossRef]
- Bentele, M.; Morgenstern, K.; Krabel, D. Lophodermium seditiosum Minter, Staley & Millar seed-borne on Pinus sylvestris. J. For. Landsc. Res. 2014, 1, 1–8. [Google Scholar] [CrossRef]
- Rajkovic, S.; Markovic, M.; Rakonjac, L. Incubation Methods for Forecasting the Occurrence and Development of Lophodermium seditiosum Minter, Staley & Millar on Pine. In Fungicides—Showcases of Integrated Plant Disease Management from around the World; Mizuho, N., Ed.; INTECH: London, UK, 2013; Chapter 8; pp. 153–183. ISBN 978-953-51-1130-6. [Google Scholar]
- Polmanis, K.; Gaitnieks, T.; Beļeviča, V.; Ruņģis, D.; Baumane, A. Occurrence of Lophodermium spp. in young scots pine stands in Latvia. In Proceedings of the Annual 23rd International Scientific Conference, the Research for Rural Development, Jelgava, Latvia, 17–19 May 2017; Latvia University of Life Sciences and Technologies: Jelgava, Latvia, 2017; pp. 14–20. [Google Scholar]
- Jansons, A.; Neimane, U.; Baumanis, I. Needlecast resistance of Scots pine and possibilities of its improvement. Mežzinātne 2008, 18, 3–18. (In Latvian) [Google Scholar]
- Neimane, U.; Polmanis, K.; Zaluma, A.; Klaviņa, D.; Gaitnieks, T.; Jansons, A. Damage caused by Lophodermium needle cast in open-pollinated and control-crossed progeny trials of Scots pine (Pinus sylvestris L.). For. Chron. 2018, 94, 155–161. [Google Scholar] [CrossRef]
- Vivas, M.; Martin, J.A.; Gil, L.; Solla, A. Evaluating methyl jasmonate for induction of resistance to Fusarium oxysporum, F. circinatum and Ophiostoma novoulmi. For. Syst. 2012, 21, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Šķipars, V.; Krivmane, B.; Runģis, D. Thaumatin-like protein gene copy number variation in Scots pine (Pinus sylvestris). Environ. Exp. Biol. 2011, 9, 75–81. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J. Analysis of phylogenetic relationships by amplification and direct sequencing of ribosomal RNA genes. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; Chapter 38; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Polmanis, K.; Klavina, D.; Gaitnieks, T.; Baumanis, I.; Lazdins, A. Genetic differences in needle cast damage of Scots pine (Pinus sylvestris L.). In Proceedings of the Annual 22nd International Scientific Conference Proceedings, Research for Rural Development, Jelgava, Latvia, 18–20 May 2016; pp. 34–40. [Google Scholar]
- Stenstrom, E.; Ihrmark, K. Identification of Lophodermium seditiosum and L. pinastri in Swedish forest nurseries using species-specific PCR primers from the ribosomal ITS region. For. Pathol. 2005, 35, 163–172. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Hantula, J.; Vainio, E. Specific primers for the differentiation of Heterobasidion annosum (s.str.) and H. parviporum infected stumps in northern Europe. Silva Fenn. 2003, 37, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Zhou, R.; Tang, T.; Shi, S. Preparative biochemistry and biotechnology simple and Efficient Isolation of High-Quality Total RNA from Hibiscus tiliaceus, a Mangrove Associate and Its Relatives. Prep. Biochem. Biotechnol. 2008, 38, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Pérez, O.; Rubio-Piña, J.A. Isolation of total RNA from tissues rich in polyphenols and polysaccharides of mangrove plants. Electron. J. Biotechnol. 2011, 14, 11. [Google Scholar] [CrossRef]
- Panter, S.; Jones, D. Age-related resistance to plant pathogens. Adv. Bot. Res. 2002, 38, 251–280. [Google Scholar] [CrossRef]
- Gould, N.; Reglinski, T.; Spiers, M.; Taylor, J.T. Physiological trade-offs associated with methyl jasmonate—Induced resistance in Pinus radiata. Can. J. For. Res. 2008, 38, 677–684. [Google Scholar] [CrossRef]
- Gould, N.; Reglinski, T.; Northcott, G.L.; Spiers, M.; Taylor, J.T. Physiological and biochemical responses in Pinus radiata seedlings associated with methyl jasmonate-induced resistance to Diplodia pinea. Physiol. Mol. Plant Pathol. 2009, 74, 121–128. [Google Scholar] [CrossRef]
- Sampedro, L.; Moreira, X.; Zas, R. Costs of constitutive and herbivore-induced chemical defences in pine trees emerge only under low nutrient availability. J. Ecol. 2011, 99, 818–827. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Carvalhais, L.C.; Schenk, P.M.; Paul, D.G. Effects of jasmonic acid signalling on the wheat microbiome differ between body sites. Sci. Rep. 2017, 7, 19. [Google Scholar] [CrossRef]
- Makandar, R.; Nalam, V.; Chaturvedi, R.; Jeannotte, R.; Sparks, A.A.; Shah, J. Involvement of Salicylate and Jasmonate Signaling Pathways in Arabidopsis Interaction with Fusarium graminearum. Mol. Plant Microbe Interact. 2010, 23, 861–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, X.; Zas, R.; Sampedro, L. Differential Allocation of Constitutive and Induced Chemical Defenses in Pine Tree Juveniles: A Test of the Optimal Defense Theory. PLoS ONE 2012, 7, e34006. [Google Scholar] [CrossRef] [Green Version]
- Bridgen, M.R.; Hanover, J.W. Indirect selection for pest resistance using terpenoid compounds. In Proceedings of the Third International Workshop on the Genetics of Host-Parasite Interactions in Forestry, Wageningen, The Netherlands, 14–21 September 1980; pp. 161–168. [Google Scholar]
- Stenlid, J.; Swedjemark, G. Differential growth of S- and P-isolates of Heterobasidion annosum in Picea abies and Pinus sylvestris. Trans. Br. Mycol. Soc. 1988, 90, 209–213. [Google Scholar] [CrossRef]
- Piri, T.; Korhonen, K. Infection of advance regeneration of Norway spruce by Heterobasidion parviporum. Can. J. For. Res. 2001, 31, 937–942. [Google Scholar] [CrossRef]
- Wainhouse, D.; Staley, J.T.; Jinks, R.; Morgan, G. Growth and defence in young pine and spruce and the expression of resistance to a stem-feeding weevil. Oecologia 2008, 158, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Pavia, H.; Toth, G.B.; Åberg, P. Optimal Defense Theory: Elasticity Analysis as a Tool to Predict Intraplant Variation in Defenses. Ecology 2002, 83, 891–897. [Google Scholar] [CrossRef]
- Minter, D.W.; Millar, C.S. Ecology and biology of three Lophodermium species on secondary needles of Pinus sylvestris. Eur. J. For. Pathol. 1980, 10, 169–181. [Google Scholar] [CrossRef]
- Diwani, S.A.; Millar, C.S. Infection processes of three Lophodermium species on Pinus sylvestris L. In Proceedings of the Conference on Recent Research on Conifer Needle Disease, Gulfport, MI, USA, 14–18 October 1984; pp. 22–27. [Google Scholar]
- Diwani, S.A.; Millar, C.S. Pathogenicity of three Lophodermium species on Pinus sylvestris L. Eur. J. For. Pathol. 1987, 17, 53–58. [Google Scholar] [CrossRef]
- Snepste, I.; Skipars, V.; Belevica, V.; Krivmane, B.; Voronova, A.; Rauda, E.; Veinberga, I.; Rungis, D. Induced resistance re-sponses in Scots pine (Pinus sylvestris L.). In Proceedings of the International Scientific Conference, Knowledge based Forest Sector, Riga, Latvia, 4–6 November 2015; pp. 130–135. [Google Scholar]
- Minter, D.W.; Millar, C.S. A study of three pine inhabiting Lophodermium species in culture. Nova Hedzvigia 1980, 32, 361–368. [Google Scholar]
- Katagiri, F. A global view of defense gene expression regulation—A highly interconnected signaling network. Curr. Opin. Plant Biol. 2004, 7, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Bowling, S.A.; Clarke, J.D.; Liu, Y.; Klessig, D.F.; Dong, X. The cpr5 Mutant of Arabidopsis Expresses Both NPR1-Dependent and NPR1-Independent Resistance. Plant Cell 1997, 9, 1573–1584. [Google Scholar] [CrossRef] [Green Version]
- Mauch, F.; Mauch-Mani, B.; Gaille, C.; Kull, B.; Haas, D.; Reimmann, C. Manipulation of salicylate content in Arabidopsis thaliana by the expression of an engineered bacterial salicylate synthase. Plant J. 2001, 25, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Heil, M.; Bostock, R.M. Induced Systemic Resistance (ISR) Against Pathogens in the Context of Induced Plant Defences. Ann. Bot. 2002, 89, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Oszako, T.; Voitka, D.; Stocki, M.; Stocka, N.; Nowakowska, J.A.; Linkiewicz, A.; Hsiang, T.; Belbahri, L.; Berezovska, D.; Malewski, T. Trichoderma asperellum efficiently protects Quercus robur leaves against Erysiphe alphitoides. Eur. J. Plant Pathol. 2021, 159, 295–308. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Seedlings | Direct MeJA Application | MeJA Vapor Treatment | |
|---|---|---|---|
| Total no. of seedlings / no. of seedlings utilized in inoculation experiment | 22/22 | 28/19 | 30/30 |
| Mortality of MeJA-treated seedlings (no.) | - | 9 | 0 |
| Infection frequency (%) detected by PCR | 100 | 100 | 63 |
| Mean longitudinal H. annosum growth ± standard error, mm | 58.32 ± 6.50 | 24.58 ± 4.86 | 13.34 ± 1.71 * |
| Minimum and maximum longitudinal H. annosum fungal growth, mm (SD) | 13.00–111.00 (30.49) | 3.50–75.00 (21.17) | 4.00–33.00 (7.45) * |
| Mortality of H. annosum inoculated seedlings, no. (%) | 3 (13) | 5(26) | 0 * |
| Control Seedlings | MeJA-Treated Seedlings | |
|---|---|---|
| Total number of seedlings inoculated with L. seditiosum | 77 | 85 |
| Needle cast damage grade 1 (0–5%) | 41 | 13 |
| 2 (6–35%) | 31 | 3 |
| 3 (36–65%) | - | 2 |
| 4 (66–95%) | - | - |
| 5 (96–100%) | 5 | 5 |
| No yellow or brown spots | - | 62 |
| Number of individuals where L. seditiosum was detected using PCR analyses | 62 | 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šņepste, I.; Krivmane, B.; Šķipars, V.; Zaluma, A.; Ruņģis, D.E. Induction of Defense Responses in Pinus sylvestris Seedlings by Methyl Jasmonate and Response to Heterobasidion annosum and Lophodermium seditiosum Inoculation. Forests 2021, 12, 628. https://doi.org/10.3390/f12050628
Šņepste I, Krivmane B, Šķipars V, Zaluma A, Ruņģis DE. Induction of Defense Responses in Pinus sylvestris Seedlings by Methyl Jasmonate and Response to Heterobasidion annosum and Lophodermium seditiosum Inoculation. Forests. 2021; 12(5):628. https://doi.org/10.3390/f12050628
Chicago/Turabian StyleŠņepste, Ilze, Baiba Krivmane, Vilnis Šķipars, Astra Zaluma, and Dainis E. Ruņģis. 2021. "Induction of Defense Responses in Pinus sylvestris Seedlings by Methyl Jasmonate and Response to Heterobasidion annosum and Lophodermium seditiosum Inoculation" Forests 12, no. 5: 628. https://doi.org/10.3390/f12050628
APA StyleŠņepste, I., Krivmane, B., Šķipars, V., Zaluma, A., & Ruņģis, D. E. (2021). Induction of Defense Responses in Pinus sylvestris Seedlings by Methyl Jasmonate and Response to Heterobasidion annosum and Lophodermium seditiosum Inoculation. Forests, 12(5), 628. https://doi.org/10.3390/f12050628