1. Introduction
Forestry has historically focused on maximizing the wood production of economically desirable tree species. Recently, increasing environmental concern and a growing body of scientific literature has led to a greater awareness of forests as ecological systems with inherent value that needs careful consideration. Forests provide a wide range of important ecosystem services, e.g., climate regulation, carbon sequestration and storage, water regulation, biological control, and nutrient cycling [
1]. Indeed, forests also support biodiversity and provide habitats for wildlife species; the term ‘habitat’ indicates the set of resources and conditions necessary to support an animal population through space and time [
2]. Therefore, forest management can unconsciously or intentionally alter resources for forest-dwelling wildlife species by interfering with successional stages and altering composition of species, vertical stratification of trees, and age structure [
3]. Forestry practices may additionally alter microscale habitat conditions such as temperature, light, moisture, soil, and litter status [
4]. It follows that any forest management practice, including an absence of direct interventions (e.g., in protected areas), has potential short- and long-term effects on the availability, distribution, and quality of habitats resources for forest-dwelling wildlife species [
5]. Because habitat requirements vary deeply among wildlife species, and in accordance with the sequence of seral stages, timber harvest can improve habitat quality for some species while at the same time deteriorating habitat suitability for others. Accordingly, large-scale logging in mature stands draws back the vegetation community from late to early successional stages, benefitting wildlife species more adapted to the early successional stages [
6].
Several studies have investigated the effects of forest management on habitat suitability of forest-dwelling wildlife (and biodiversity in general), including some meta-analyses. According to these investigations, the type and extent of the effects of forestry on wildlife habitat vary deeply depending, above all, on species and taxonomic group [
7,
8,
9], the forest management system [
9,
10], the extent of logging [
11], the time since logging [
5,
12,
13], and the overall landscape matrix [
13,
14], revealing a mix of complementary and compound relationships. Silviculture is, therefore, a powerful tool to achieve wildlife management aims. Creating and manipulating disturbance regimes of forests can enhance the spatial and temporal arrangement of key forest characteristics used or selected by a given wildlife species and can therefore improve quality of the species’ habitat [
15]. This is especially true for the small or threatened wildlife populations in greatest need of active management interventions, and of which the long-term effective conservation rests on structural improvements of their habitat at the landscape scale [
5].
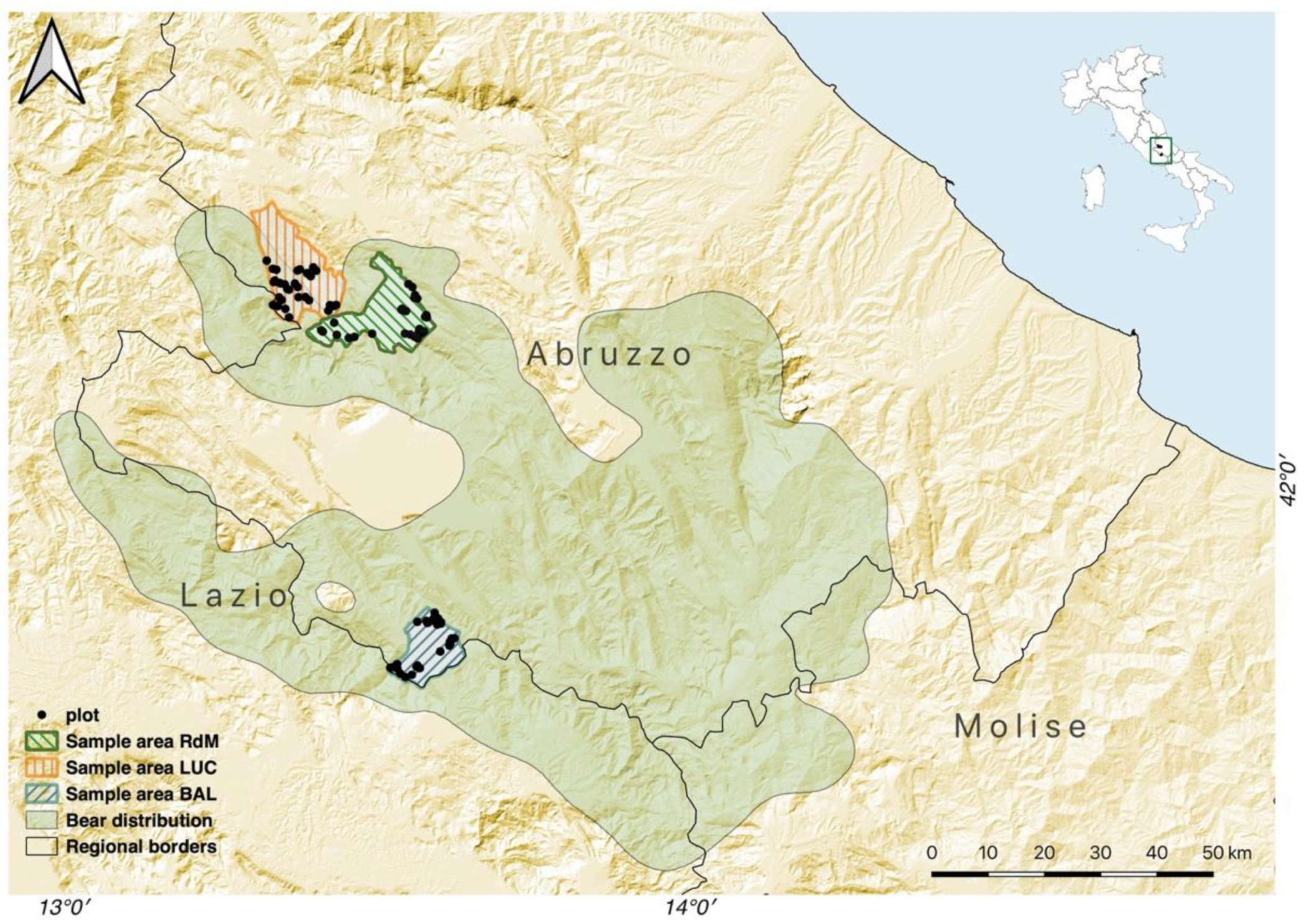
The Apennine brown bear (
Ursus arctos marsicanus) survives in a small and isolated population consisting of about 50 individuals [
16] confined to the Abruzzo, Lazio, and Molise National Park (PNALM) and its surrounding region in central Italy. This bear population faces a severe risk of extinction due to small size and reduced genetic variability [
17,
18]. In addition to mitigation of human-caused mortality and disturbance, habitat management to ensure long-term productivity of the forest ecosystem has been advocated to facilitate expansion of the species’ range [
19,
20,
21]. In fact, no significant expansion of the bear range has been recently observed, despite the availability of suitable habitat at the landscape scale and a number of protected areas therein [
20,
22,
23]. This is probably due to high levels of human-caused mortality [
23], a relatively low reproductive performance [
24], and the likely contribution of stochasticity [
18,
25]. Based on projections of habitat suitability at the landscape scale, the conditions and connectivity of high-quality habitat patches in the central Apennines have been deemed adequate to support a viable population of Apennine bears [
20]. However, available estimates of habitat and landscape suitability did not take into account forest management practices and their effects on forest structure and productivity. Indeed, a more in-depth knowledge of how forest structure and management practices relate to habitat suitability and food productivity for Apennine bears is therefore fundamental to inform forest management [
26].
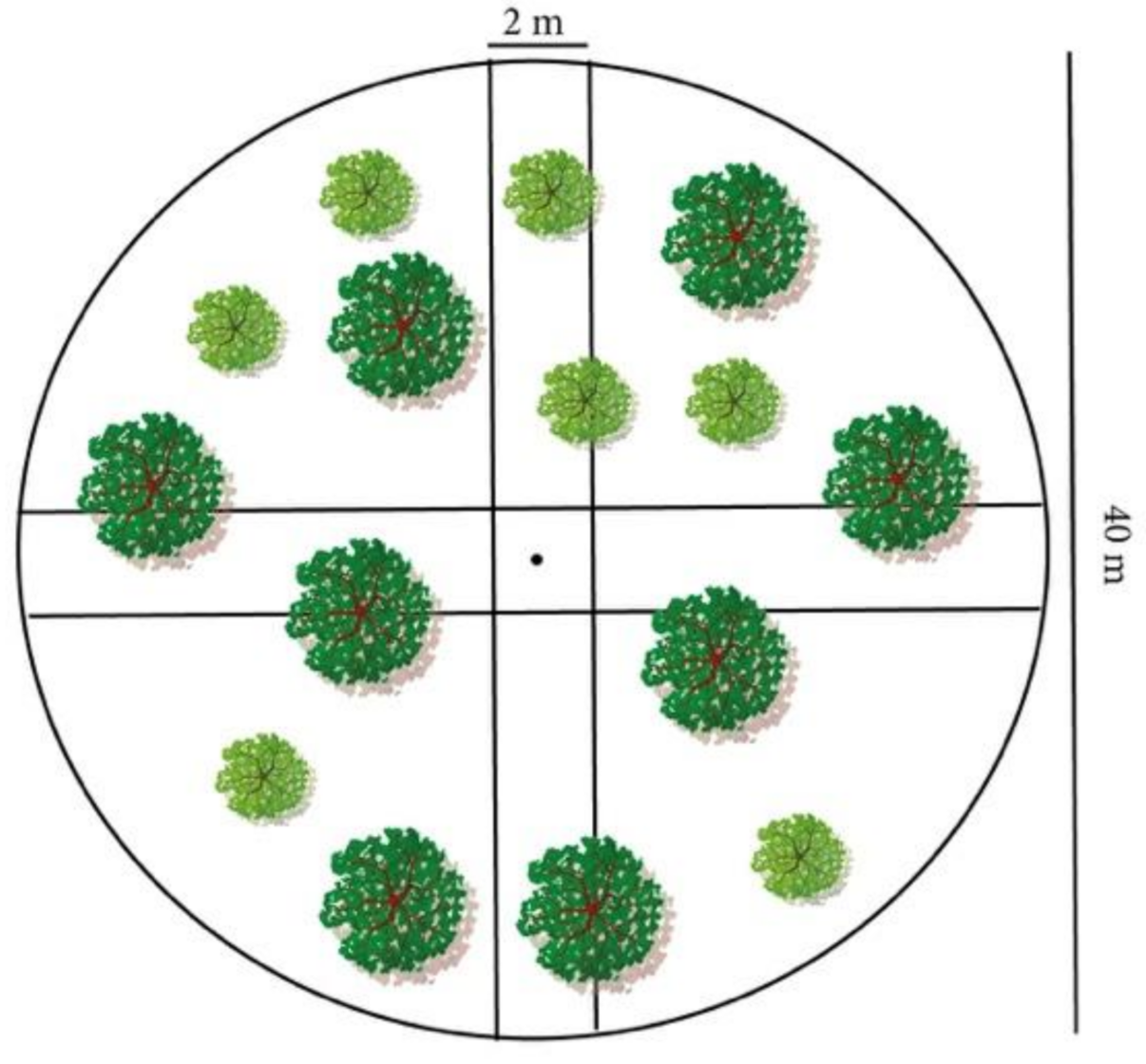
The availability and accessibility of nutritionally adequate food resources are critical drivers of high-quality bear habitat [
27,
28,
29]. Similarly, to other south-European bear populations [
30], Apennine bears have a prevalently vegetarian diet and, even though they consume a wide variety of foods, plant-based items tend to be consumed more frequently throughout the year [
26]. In spring and early summer, herbaceous vegetation represents 19–31.7% of their digestible energy, complemented by wild ungulates and a significant contribution of ants, while berries contribute 56.5% in summer, and in fall, the majority of digestible energy is represented by hard mast (66.9%) followed by fleshy fruits (26.3%) [
26]. The mountain forests of the central Apennines are thought to broadly fulfil the habitat requirements for Apennine brown bears, such as food and thermal and security cover [
22,
23]. However, a lack of detailed data on the relationship between fine-scale forest structure and composition and food productivity prevents the planning of effective forest management to assist the conservation of Apennine bears. Nevertheless, forestry practices are known to play a critical role in affecting the availability of seasonal key foods for bears, especially during hyperphagy [
31]. This is a critical period of the year for bears, as they need to swiftly accumulate fat for the wintering period. In many bear populations, the amount of fat accumulated during fall is correlated with reproductive success [
32,
33,
34,
35], as fat storage in reproducing females is an essential source of energy for cub production and lactation [
36]. In particular, the availability of hard mast (i.e., beechnuts, acorns) during fall, complemented by fleshy fruits, seems to boost reproductive success in Apennine female bears [
37]. Although a substantial body of research has determined how forest management can affect habitat quality for black (
Ursus americanus) (e.g., [
38,
39,
40]) and grizzly (
Ursus arctos) (e.g., [
13,
31,
41,
42,
43,
44,
45]) bears, no such information is currently available for Apennine brown bears [
46]. To fill this knowledge gap, the aim of this study was to investigate the relationship between fine-scale forest structure metrics, reflecting forest structure and management practices, and indirect measures of productivity of key foods for Apennine bears. By collecting forest structural metrics during the fall, we focused on forest-related key foods consumed by bears during hyperphagia (soft and hard mast), even though we also made inferences regarding key foods consumed by bears in other seasons. Since topography is known to affect forest structure and composition [
47,
48] and, in turn, habitat conditions for forest-dwelling species [
49], we also considered topographical features as factors potentially affecting productivity of bear key foods. We hypothesized that food availability for bears varies as a function of the structural and topographical characteristics of forests. To this end, we predicted that forest structural predictors could be identified for each food item, implying that these can be manipulated to improve the habitat productivity of bears.
4. Discussion
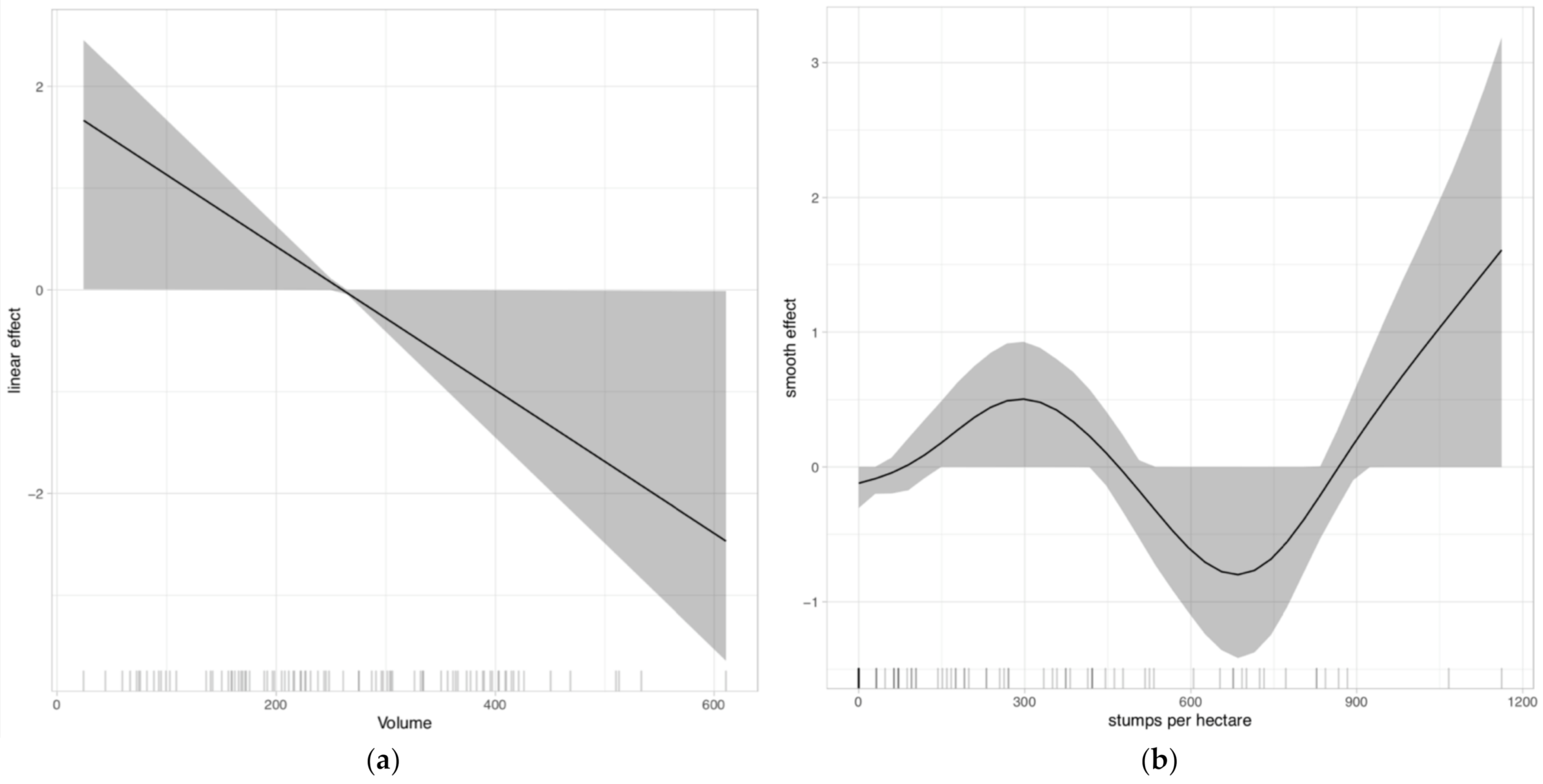
This study was the first to attempt an assessment of the functional relationship between characteristics of Apennine forests and the productivity of key foods for Apennine bears. Since beech and oak forests are predominant in our study area, it is not surprising that forest typology exerted the most relevant effect on hard-mast availability. We found negative relationships between basal area of hard-mast-producing species (our proxy of fruit productivity) and both the number of cut stumps and the deadwood decay stage. This suggests that density may be a detrimental factor for radial growth, development of crown area, and fruit production. Deadwood availability is related to high competition in stands with high tree density [
84] and the main force of accumulation was forestry, since in the recent past, wood and logging slash were regularly removed [
85]. Considering the number of decay classes as an index of forestry operations (i.e., time since the last intervention), hard-mast availability was lower in stands where deadwood was successfully stored (i.e., greater time elapsed since the last forestry operation). Traditionally, in central Italy, deadwood was removed to avoid the risk of forest fires and to limit the potential spread of pathogens. In addition, rural communities used to collect deadwood to be used as fuel. Now that the forests are no longer managed and the mountains are depopulating, dead wood can accumulate, and its degree of decomposition can therefore be an indication of the time of the last intervention. In the studied latitudes and climatic conditions, the decay time is not so fast, especially since we considered in our analysis the largest fraction of dead wood (snags, logs, and stumps). Our results also showed the negative role of overmature coppice in hard-mast productivity; this is probably due to patterns of spatial aggregation of the stems, which are aggregated in stumps. The high density of stems and the vertical layering of these stands produce a higher level of competition for light, [
86] therefore affecting the development of the crown [
87] and influencing fruit production [
88,
89]. This trend seems also to be confirmed by local studies of acorns and beechnuts production that indicated a low hard-mast provision in overmature coppice stands or stands in the early stage of conversion Our results suggest that a lower tree density in overmature coppices and irregular high forests could improve hard-mast availability, as also evidenced in an experimental study of Apennine beech coppices under conversion to high forest [
90].
Another critical factor affecting hard-mast availability was the level of interspersion of producing species at a local (i.e., bear’s home range scale) and landscape scale. Reflecting a common pattern across the Apennines, we found that the European beech grows in pure stands with a low occurrence of other hard-mast producers, and that the contact zones with oaks depends mainly on slope and aspect [
91]. Mixed stands of oak and beech (or distributed in patches) occur on sites where the competitive capacity of beech is reduced and oak can compete successfully. The presence of beech mixtures can negatively affect the oak increment of a basal area [
92] proportionally more in closed-canopy forests, where a lower light transmittance creates an unfavorable habitat for heliophilic oaks. We determined that hard-mast-producing species also occurred sporadically in old hornbeam coppices: beech and Turkey oak at the higher altitudes and the downy oak at the lower altitudes. The secondary succession of these stands is only theoretical [
93] as there are no studies or data about their evolution. However, [
94] showed a decline in light-demanding oaks and an increase in nutrient-demanding shady species after the abandonment of coppicing.
A good interspersion of hard-mast producers is an important indicator of bear habitat quality, as the spatial distribution of food resources strongly affects the size, configuration, and location of bear home ranges at the landscape scale [
95,
96,
97]. Moreover, hard-mast production exhibits large temporal and spatial variation, as this is a pulsed resource that is subject to marked annual fluctuations and occasional mast failures [
98,
99]. Silvicultural practices can enhance the functional diversity of hard-mast species for bears [
100] at a landscape scale, and also ensure supplies of alternate foods in years when the major food source is scarce. To promote oak diffusion, or alternatively to complement beech, hornbeam and mixed old coppice stands (Turkey oak dominated) could be managed by thinning to reduce the tree density removing hornbeam shoots and some old oaks standards around the best oak shoots and secondary species. The removal of some senescent plants, even though this would cause an initial decrease in the production of acorns, could be necessary to initiate new oak stands, contributing to hard-mast production and mitigating future oak losses driven by oak decline and ecological succession [
101].
With respect to soft-mast-producing species, we were able to find a significant relationship with slope. Although no studies on soft-mast-producing trees support our result, it is known that slope is negatively related to percent cover of soft-mast shrub and herbaceous species [
102], understory plant richness [
103], and basal area of trees in plantation sites [
104]. Terrain morphology (slope, altitude, and aspect) influences many ecological aspects of forests [
105], but we believe that slope in our models captured the effects of more than just a topographic factor. All the soft-mast-producing species that we considered in our study are heliophilous, even though some species (e.g., service trees) are shade-tolerant [
106] and able to remain under a dense canopy layer [
107,
108]. Besides ecological traits, the distribution of soft-mast tree species was likely shaped by past forestry practices designed to eliminate discarded species and enhance production of commercially valued species [
109,
110]. The joint effect of ecological requirements and the legacy of past management has resulted in a scattered distribution of these species, rarely as an old standard in coppice stands and most frequently in the subcanopies of oak and beech forests. The presence of these sporadic species therefore could be related to past forestry practices such as coppicing and to more open forest structures with greater light availability [
111]. It is also negatively related to forests along steep slopes, which are traditionally managed through high tree density and a low rate of silvicultural disturbance, essentially to increase stability and to reduce soil erosion [
112]. In such stands, unfavorable conditions exist for soft-mast-producing species, due to the fact that canopy closure reduces light infiltration and increases competitive effects [
113,
114,
115]. As a result, in our study, we observed a very low frequency and often a lack of soft-mast-producing species (
Table 6). Being light-demanding and poorly competitive species, their promotion runs through silvicultural practices aimed at reducing competition and enabling an adequate crown development. For example, group selection [
116] and tree-oriented silviculture [
117] can be enforced even in trees that have long been suppressed [
118].
Regarding herbaceous vegetation, ants, and wild ungulates, the results of our models provided only preliminary clues, since the abundance and occurrence of these food items is predictably largely affected by phenology. Nonetheless, we believe that our findings will be useful to inform further investigations and more adequate seasonal sampling. Aspect and slope strongly affected the distribution of herbaceous vegetation. In particular, south-facing aspects were the richest and north-facing ones showed comparable trends, possibly because environmental heterogeneity creates a mosaic of microenvironments shaping different plant communities. Shifts in microclimate across north- and south-facing plant communities are a well-established pattern [
119,
120,
121]. South-facing slopes tend to receive greater insolation, resulting in drier conditions that support thermophile species (e.g., graminoids). North-facing slopes receive less sunlight, creating cool and moist conditions favorable for moisture-loving hygrophile plants. Our findings also suggest that greater slope steepness can negatively influence herb cover, in agreement with similar studies [
103,
122,
123,
124]. Slope is an environmental factor known to influence soil moisture (the most important driver for herb cover in forests: [
125,
126,
127,
128], air humidity, and soil chemistry [
125]). Moreover, we showed that increasing basal area negatively affects herb cover regulating light transmittance through the canopy [
129,
130,
131]. The negative association we revealed with understory cover indicates that light availability on the ground limits the forest herbaceous layer, similarly to other studies [
103,
120,
132,
133,
134]. We caution that the relationships between basal area and herb cover should be interpreted according to plant functional groups—this caveat being relevant for ‘true forest species’ (with habitat requirements strictly tied to closed canopy forests) for which increased light availability corresponds to a decrease of their incidence [
135,
136]. For this reason, and by virtue of the complex response of the herbaceous vegetation to thinning [
136], it would be necessary to work at the landscape scale and to provide different forest structures with different levels of light requirements.
Concerning the occurrence of animal foods for Apennine bears, we revealed that enhanced occurrence of ants is to be expected in forest stands characterized by low volumes and high tree density, similarly to other Mediterranean mountain environments [
137], which are characteristics of young forests and aged coppice. Ants are thermophilic species and their presence in forests is linked to low forest canopy and warmer microclimates [
138]. Some species of ants fed upon by bears [
26,
139] are indeed associated with deadwood [
74,
76,
137,
140], which ants use as a substrate or to build nests [
141]. In dense forests, high competition levels produce high quantities of deadwood, but in the Apennines, the main driver of deadwood availability is past forest management regimes. As expected, the best predictor of deadwood was the number of decay classes, which is an index of the management forest history of the stand [
67,
68].
With regard to wild ungulates, we failed to find a robust relationship with forest structure. Although our findings indicated an effect of altitude and soil type on wild ungulate occurrence, the sample size was too low to draw any meaningful conclusion.
In conclusion, the availability of natural food resources is essential to guaranteeing the long-term conservation of the Apennine brown bear, and our study highlights that key bear foods are indeed affected by forest structural and topographical characteristics. The productivity of hard mast was positively associated with both forest typology and high forest system and negatively related to both the time from the last forest utilization and the amount of deadwood. Past forest management was also related to availability of soft-mast-producing species, since forests on steep slopes have been historically managed with high tree densities and low silvicultural disturbance. Our findings also indicate that herb cover was negatively affected by terrain steepness and basal area, while herb productivity was positively affected by northern and southern exposure. Finally, we revealed that forests characterized by low volume and high density corresponded to a higher richness of forest ants. Evidence concerning the effect of forest structure in the occurrence of wild ungulates was less conclusive, but may provide useful hints for further studies.
Our study has some relevant caveats. First, due to logistical constraints, we could collect data only in a single bear dietary season and with no annual replicates. Although we chose the most energetically critical season for bears (i.e., hyperphagia), this was hazardous for assessing the productivity of both hard and soft mast and of herbaceous vegetation, because seed production is characterized by annual variability and occurrence of masting. To overcome this problem, we used basal area, since it has less annual and interannual variability. Second, as we used proxies of key food productivity, our findings are not based on direct measurements of the variables of interest. In order to obtain greater robustness, we encourage future studies to obtain direct measures of productivity and to link resource availability with bear vital rates and population growth [
34]. Nevertheless, our study provides general indications on how management of forest structure could enhance productivity of key bear foods. As such, it represents an important first step towards filling the knowledge gap concerning the functional relationship between forest structure and habitat productivity for Apennine bears, and possibly other forest-dwelling species.






{kind=link}
{kind=link}
{kind=link}