
Productivity of Eucalyptus pellita in Sumatra: Acacia mangium Legacy, Response to Phosphorus, and Site Variables for Guiding Management


Abstract
:1. Introduction
- Examine the long term trend in productivity over 30 years, and across four successive rotations (three rotations of A. mangium, followed by E. pellita) at the same site.
- Evaluate the impacts of successive applications of management practices representing depletion or retention of slash and litter over the three previous rotations of A. mangium, (legacy-effects) on productivity of E. pellita.
- Examine the question, what is the nature of growth responses to P (rates of application and the extent across sites) to E. pellita planted on ex. Acacia sites, given the history of retention of slash and litter, and the application of P to previous A. mangium rotations?
- Using the data from the network of experiments across a range of sites and contrasting growth rates, examine how the site and stand influence the variation in site productivity.
2. Materials and Methods
2.1. Productivity over Successive Rotations
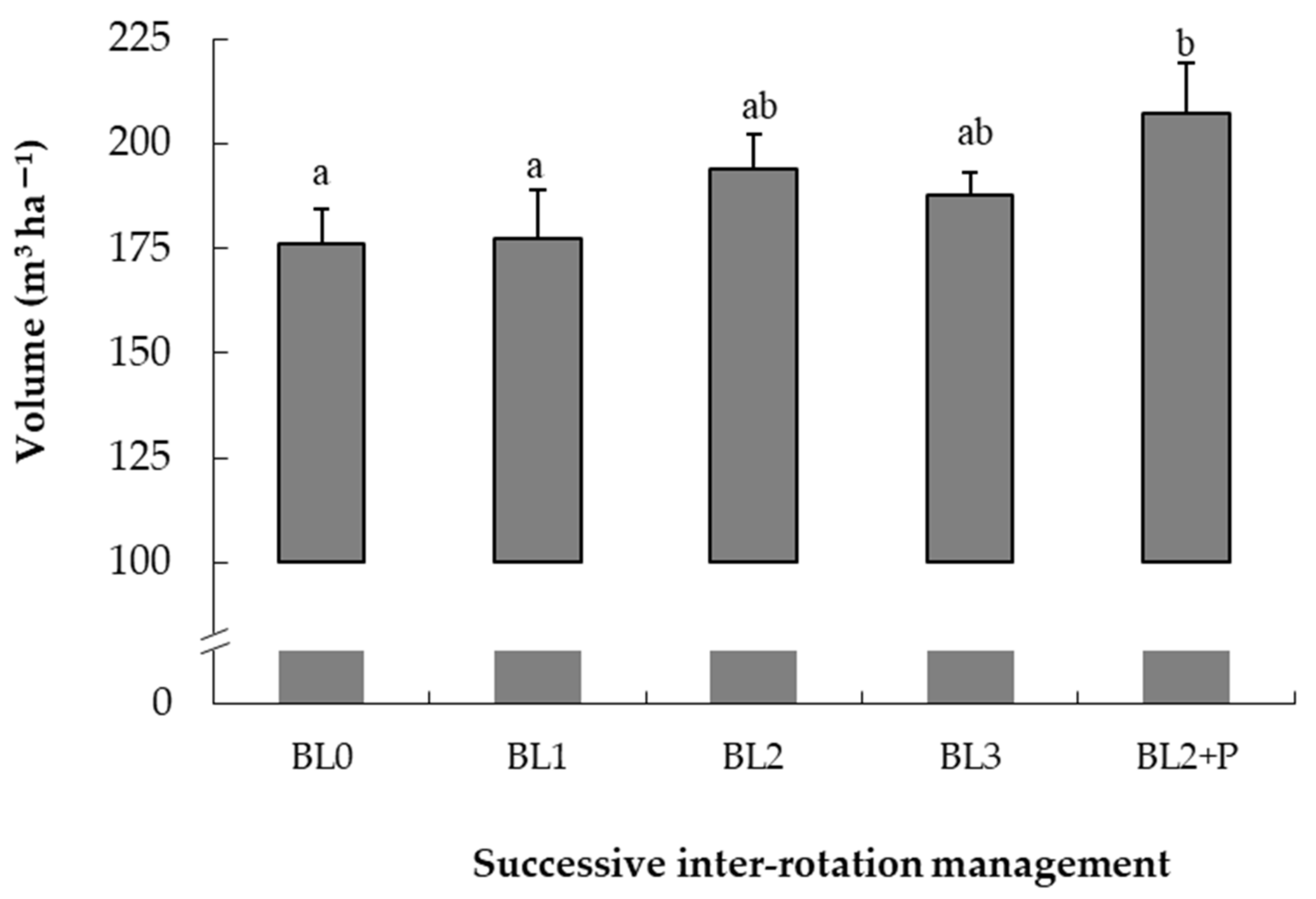
2.1.1. Treatments
- BL0
- All above ground biomass removed: All merchantable stems (more than 7 cm top-end diameter, over bark) slash, litter and understorey were removed.
- BL1
- Whole tree harvested: All trees with merchantable wood including crown were removed from the plots. Litter and understorey were retained.
- BL2
- Merchantable stems harvested: Remaining stem top-ends, branches, leaves, litter and understorey were retained and re-distributed evenly in the same plot.
- BL3
- High organic matter: Same as BL2 but slash from the BL1 plots was brought in and distributed evenly over the slash already present in BL2 plots. This created a high organic matter treatment for comparison. This practice would not occur in operation, but a harvested site would have many small piles of slash giving rise to patches of high organic matter.
2.1.2. Management
2.2. Responses to Phosphorus and Site Variation in E. pellita
2.3. Growth Measurements
2.4. Soil Sampling and Analysis
2.5. Statistical Analyses
3. Results
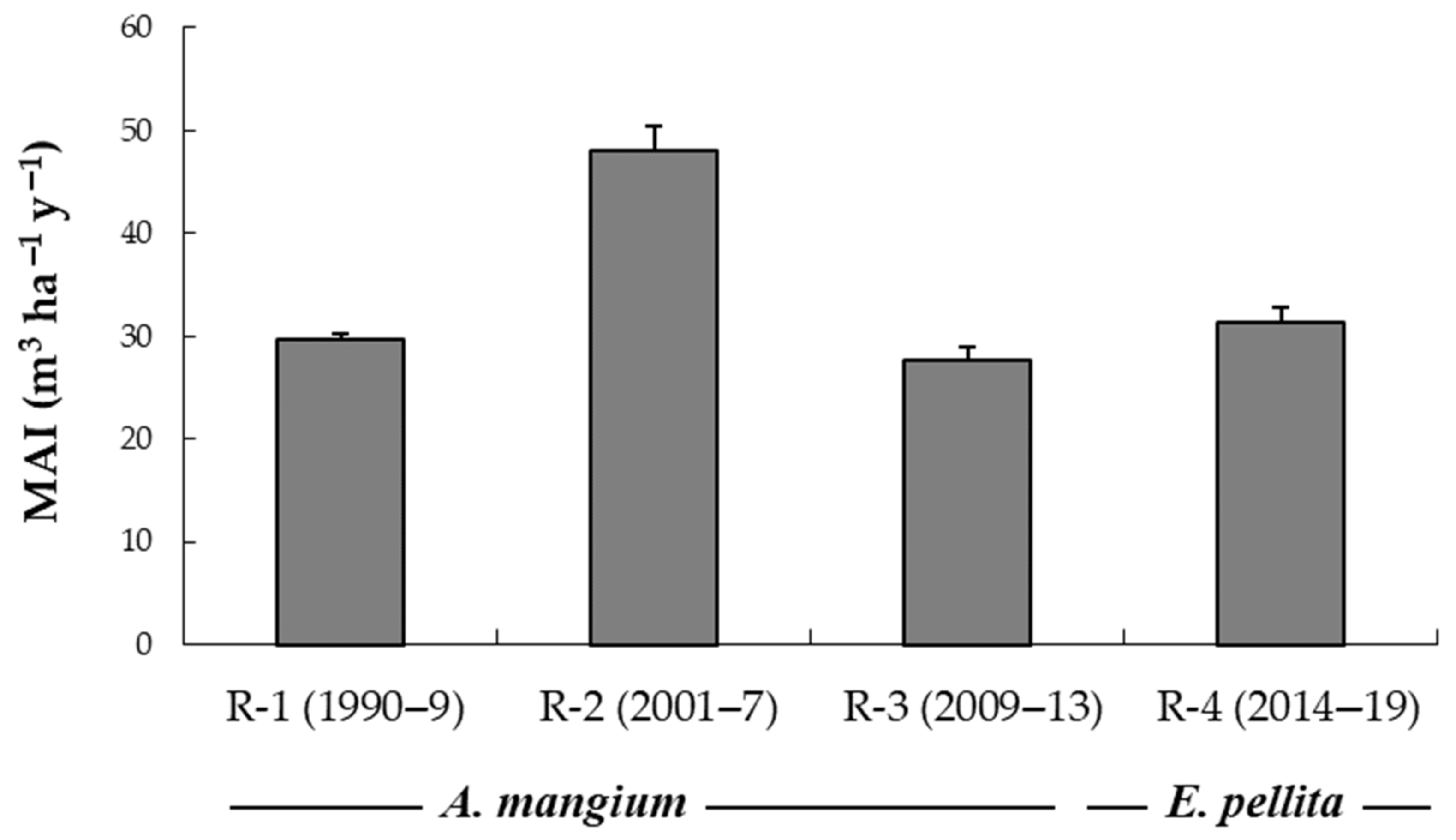
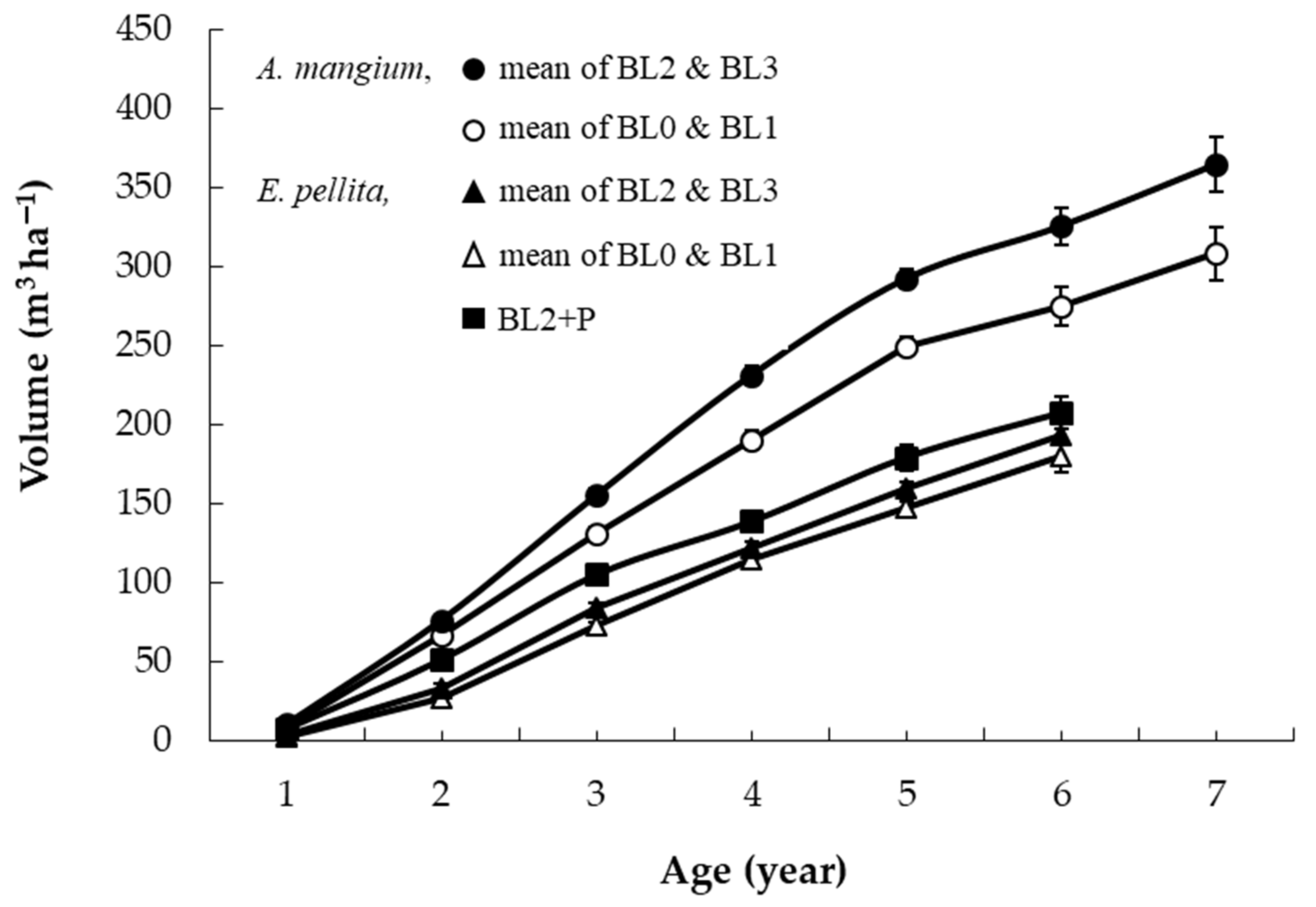
3.1. Productivity over Four Rotations and the Comparative Growth of Species
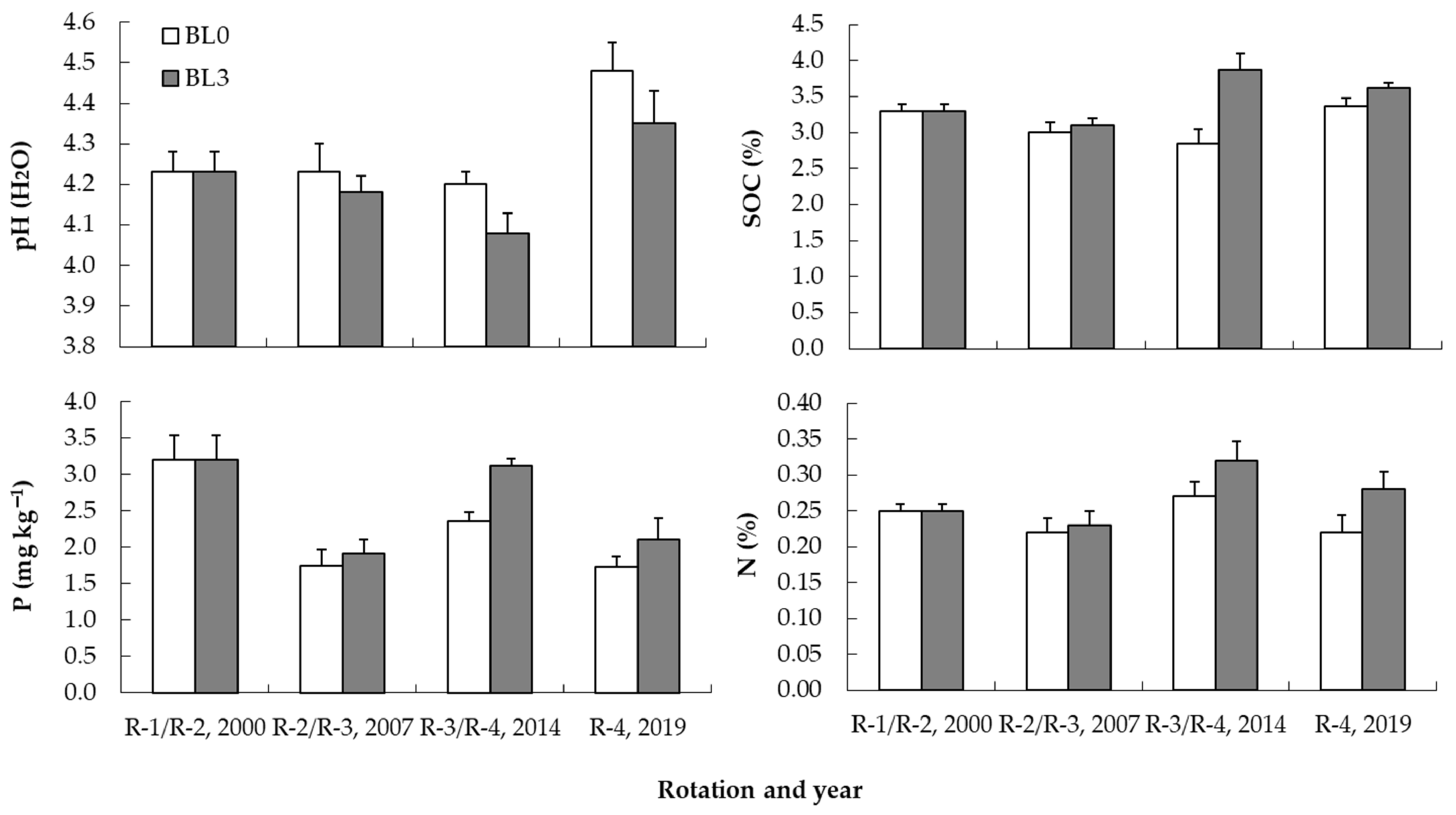
3.2. Changes in Soil Properties across Successive Rotations
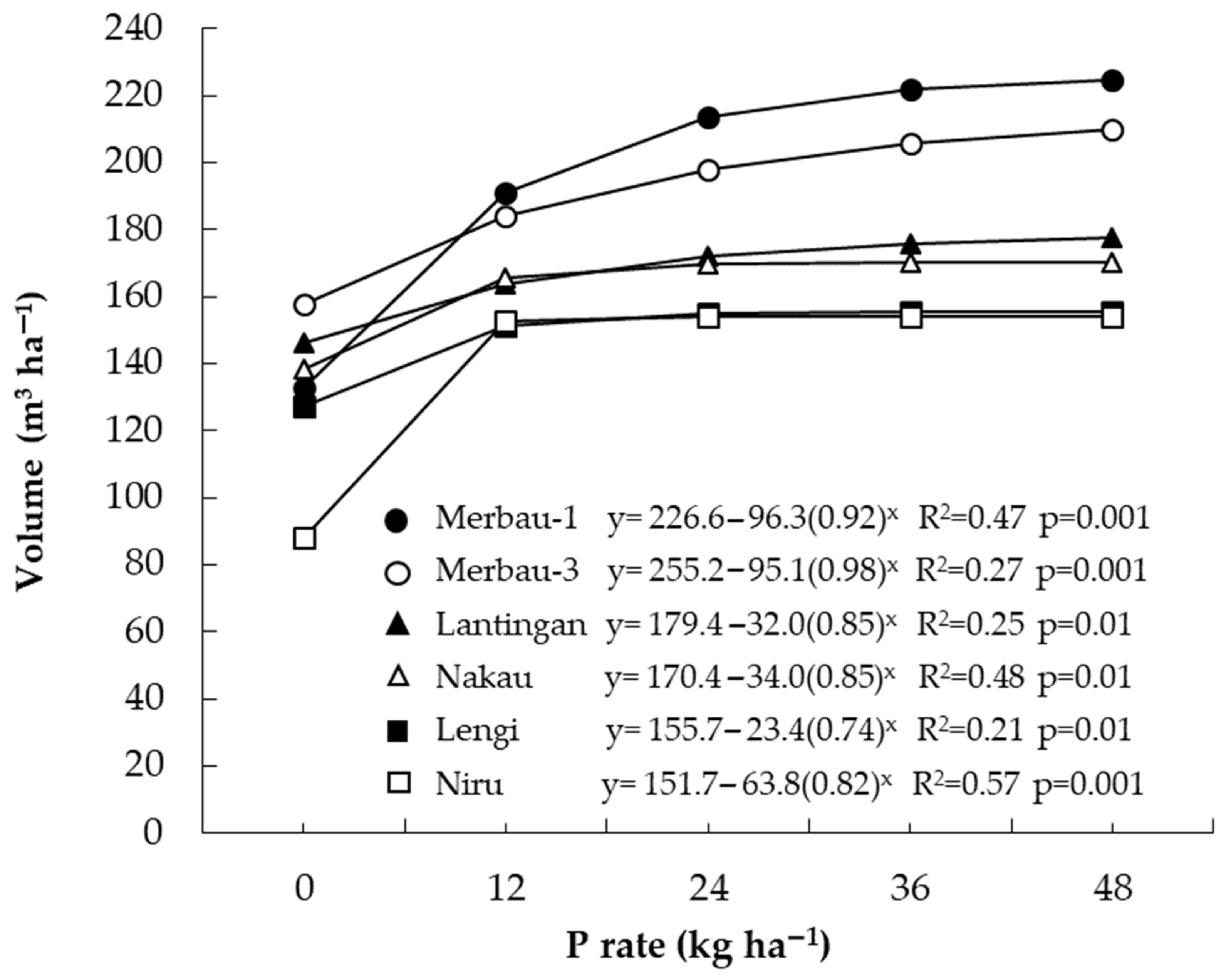
3.3. Responses to Phosphorus by E. pellita
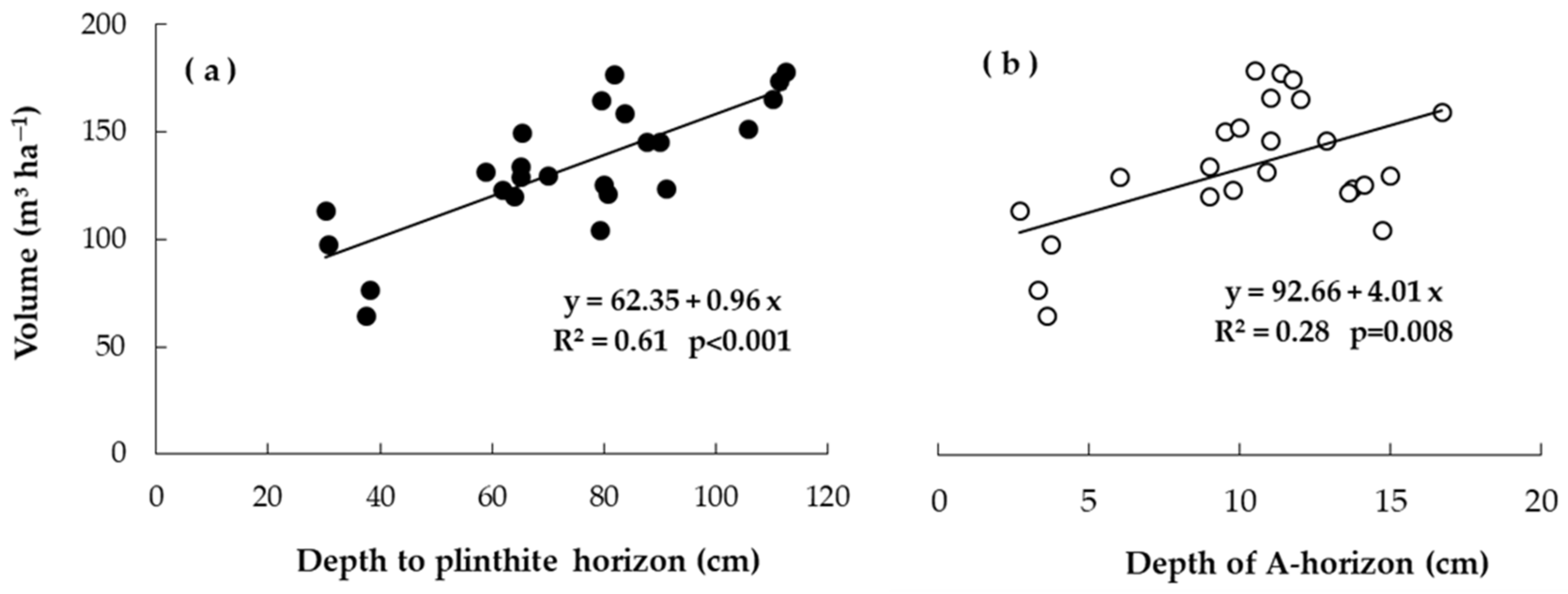
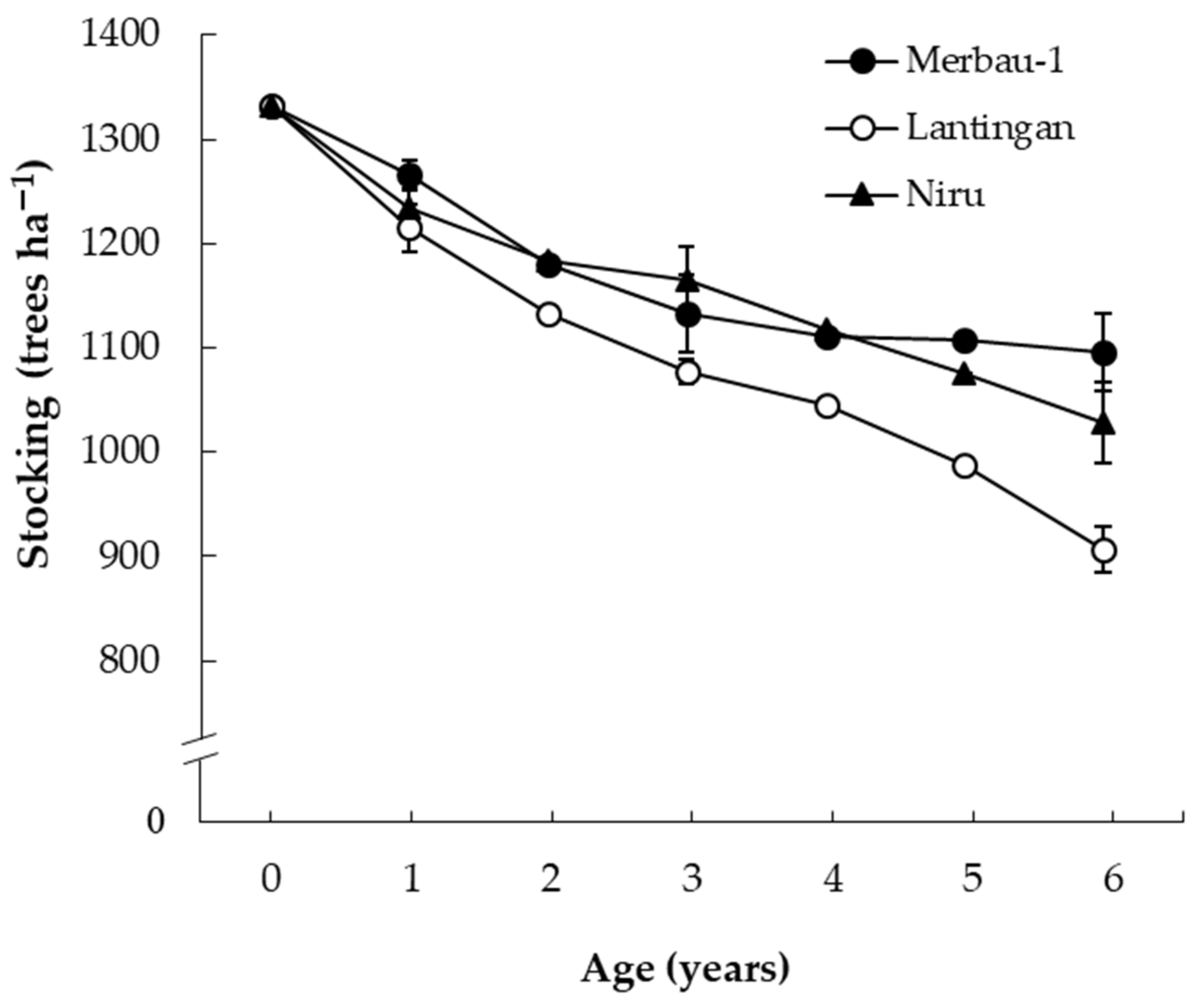
3.4. Site Variations in Productivity
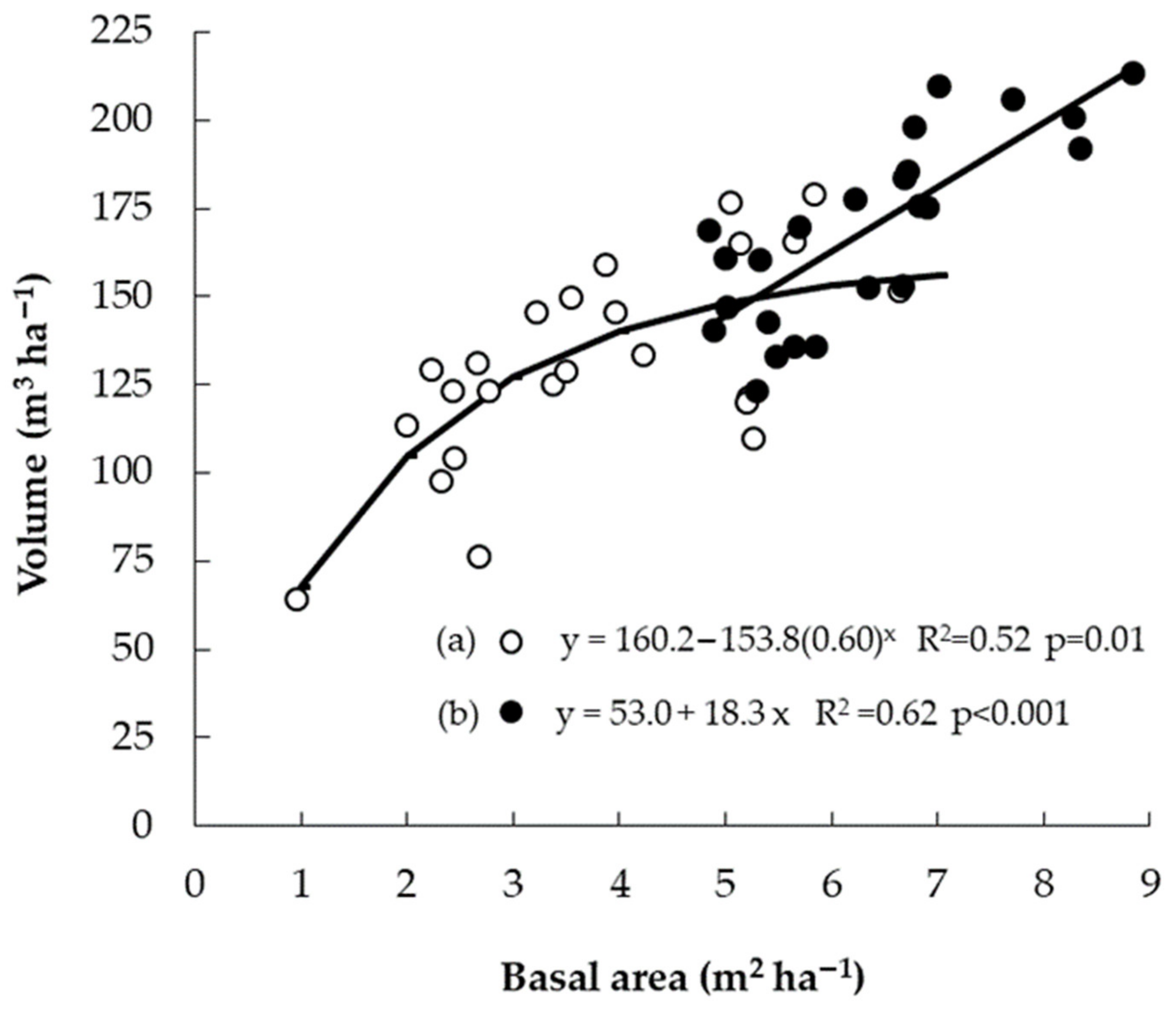
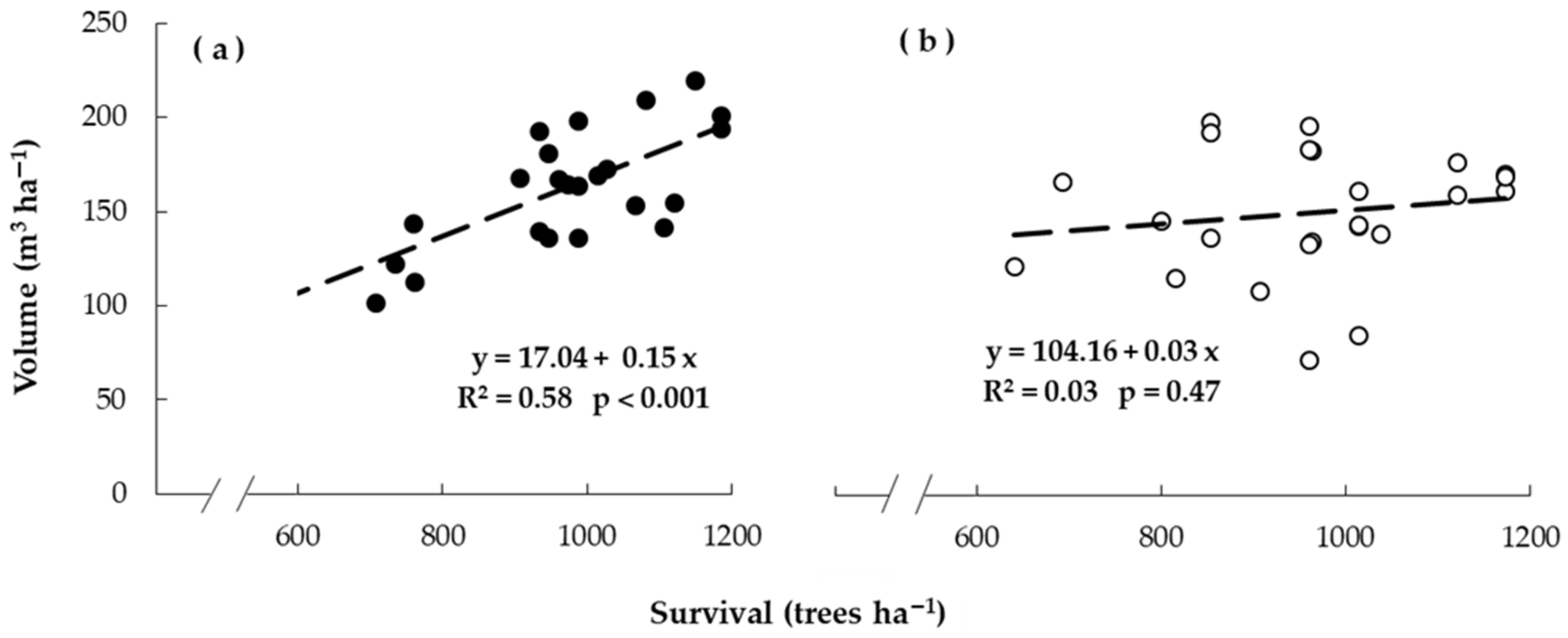
3.5. Relationship between Early Growth and Yield at Harvest
4. Discussion
4.1. Productivity of Successive Rotations
4.2. Managing Productivity across Diverse Sites
4.3. Management of Phosphorus Nutrition
4.4. Site and Wood Production
4.5. Stand Attributes and Production
4.6. Prospects for Increasing Productivity
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harwood, C.E.; Nambiar, E.K.S. Productivity of acacia and eucalypt plantation in South-East Asia. 2. Trends and variations. Int. For. Rev. 2014, 16, 249–260. [Google Scholar]
- Hardiyanto, E.B.; Nambiar, E.K.S. Productivity of successive rotations of Acacia mangium plantations in South Sumatra, Indonesia: Impacts of harvest and site management. New For. 2014, 45, 557–575. [Google Scholar] [CrossRef]
- Eyles, A.; Beadle, C.L.; Barry, K.M.; Francis, A.; Glen, M.; Mohammed, C. Management of fungal root-rot pathogens in tropical Acacia mangium plantations. For. Pathol. 2008, 38, 332–355. [Google Scholar] [CrossRef]
- Glen, M.; Yustikanti, V.; Puspitasari, D.; Francis, A.; Agustini, L.; Rimbawato, A.; Indrayadi, A.; Gafur, A.; Mohammed, C. Identification of basidiomycetes fungi in Indonesian hardwood plantations by DNA barcoding. For. Pathol. 2009, 44, 496–508. [Google Scholar] [CrossRef]
- Francis, A.; Beadle, C.; Puspitasari, D.; Irianto, R.; Agustini, L.; Rimbawanto, A.; Gafur, A.; Hardiyanto, E.; Hidayati, N.; Tjahjono, B.; et al. Disease progression in plantations of Acacia mangium affected by red root rot (Ganodema philippii). For. Pathol. 2014, 44, 447–459. [Google Scholar] [CrossRef]
- Tarigan, M.; Roux, J.; Van Wyk, M.; Tjahjono, B.; Wingfield, M. A new wilt and die-back disease of Acacia mangium associated with Ceratocystis manginecans and C. acaciivora sp. nov. in Indonesia. S. Afr. J. Bot. 2011, 77, 292–304. [Google Scholar] [CrossRef] [Green Version]
- Nambiar, E.K.S.; Harwood, C.E.; Mendham, D.S. Paths to sustainable wood supply to the pulp and paper industry in Indonesia after diseases have forces a change of species from acacia to eucalypts. Aust. For. 2018, 81, 148–161. [Google Scholar] [CrossRef]
- Hardiyanto, E.B. Operational challenges managing productivity in multispecies plantations. In Proceedings of the IUFRO Eucalyptus 2018 Conference, Montpellier, France, 17–21 September 2018. [Google Scholar]
- Data and Information Center (MOI). Energy Requirement in Indonesia’s Pulp and Paper Industry in 2019; Ministry of Industry of Indonesia: Jakarta, Indonesia, 2020; 70p. (In Indonesian)
- BPS. Statistics of Forest Production 2019; BPS Statistics Indonesia: Jakarta, Indonesia, 2019; 80p.
- Nambiar, E.K.S.; Kallio, M.H. Increasing and sustaining productivity in subtropical and tropical plantation forests: Making a difference through research partnership. In Site Management and Productivity in Tropical Plantation Forests, Proceedings of the Seventh Workshop, Piracicaba, Brazil, 22–26 November 2004; Bogor, Indonesia, 6–9 November 2006; Nambiar, E.K.S., Ed.; CIFOR: Bogor, Indonesia, 2008; pp. 205–228. [Google Scholar]
- Nambiar, E.K.S.; Harwood, C.E. Productivity of acacia and eucalyptus plantations in South-east Asia. 1. Bio-physical determinants of production; opportunities and challenges. Int. For. Rev. 2014, 6, 225–248. [Google Scholar]
- Huong, V.D.; Nambiar, E.K.S.; Hai, N.X.; Ha, K.M.; Dang, N.V. Sustainable management of Acacia auriculiformis plantations for wood production over four successive rotations in South Vietnam. Forests 2020, 11, 550. [Google Scholar] [CrossRef]
- Harwood, C.E.; Hardiyanto, E.B.; Yong, W.C. Genetic improvement of tropical acacia: Achievements and challenges. South. For. 2015, 77, 11–18. [Google Scholar] [CrossRef]
- Inail, A.M.; Hardiyanto, E.B.; Mendham, D.S. Growth responses of Eucalyptus pellita F. Muell. Plantation in South Sumatra to macronutrient fertilisers following several rotations of Acacia mangium Willd. Forests 2019, 10, 1054. [Google Scholar] [CrossRef] [Green Version]
- Mendham, D.; Greenhill, M.; Beadle, C.; Hardiyanto, E.; Wibisono, G.; Rimbawanto, A.; Fauzi, A.; Dong, T.L.; Huong, V.D.; Hai, T.A.; et al. Maximising Productivity and Profitability of Eucalypts and Acacias in Indonesia and Vietnam; Final Report of ACIAR Project FST/2014/064; Australian Centre for Agricultural Research: Canberra, Australia, 2020; 68p.
- Hardie, M.; Mendham, D.; Corkrey, R.; Hardiyanto, E.; Maydra, A.; Siregar, S.; Marolop, R.; Wibowo, A. Effects of Eucalypt and Acacia plantations on soil water in Sumatra. New For. 2018, 49, 87–104. [Google Scholar] [CrossRef]
- Hardiyanto, E.B.; Anshori, S.; Sulistyono, D. Early results of site management in Acacia mangium plantations at PT. Musi Hutan Persada, South Sumatra, Indonesia. In Site Management and Productivity in Tropical Plantation Forests, Proceedings of the Workshop, Pointe-Noire, Congo, July 2001; Guangzhou, China, February 2003; Nambiar, E.K.S., Ranger, J., Tiarks, A., Toma, T., Eds.; CIFOR: Bogor, Indonesia, 2004; pp. 93–108. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014; 359p. [Google Scholar]
- Hardiyanto, E.B.; Wicaksono, A. Inter-rotation site management, stand growth and soil properties in Acacia mangium plantations in South Sumatra, Indonesia. In Site Management and Productivity in Tropical Plantation Forests, Proceedings of the Workshops in Piracicaba, Brazil, 22–26 November 2004; Bogor, Indonesia, 6–9 November 2006; Nambiar, E.K.S., Ed.; CIFOR: Bogor, Indonesia, 2008; pp. 107–122. [Google Scholar]
- Huong, V.D.; Nambiar, E.K.S.; Quang, L.T.; Mendham, D.S.; Dung, P.T. Improving productivity and sustainability of successive rotation of Acacia auriculiformis plantations in South Vietnam. South. For. 2015, 77, 51–58. [Google Scholar] [CrossRef]
- Nurudin, M.; Ohta, S.; Hardiyanto, E.B.; Mendham, D.; Wicaksono, A.; Heriyanto, J.; Watanabe, M. Relationship between soil characteristics and productivity of Acacia mangium in South Sumatra. Tropics 2013, 22, 1–12. [Google Scholar] [CrossRef]
- Siregar, S.T.H.; Alpan, U.; Riyantoko, A. Bilangan bentuk batang Acacia mangium umur 5 tahun di Subanjeriji. Tech. Notes R D PT Musi Hutan Persada 1997, 7, 1–5. (In Indonesian) [Google Scholar]
- Supriyadi, B. Form factor for Eucalyptus pellita. Tech. Notes R D PT. Musi Hutan Persada 2011, 21, 1–5. [Google Scholar]
- Mendham, D.S.; Hardiyanto, E.B.; Wicaksono, A.; Nurudin, M. Nutrient management of contrasting Acacia mangium genotypes and weed management strategies in South Sumatra, Indonesia. Aust. For. 2017, 80, 127–134. [Google Scholar] [CrossRef] [Green Version]
- O’Hehir, J.F.; Nambiar, E.K.S. Productivity of successive rotations of P. radiata in South Australia over a century. For. Ecol. Manag. 2010, 209, 1857–1869. [Google Scholar] [CrossRef]
- Rocha, J.H.T.; Menegale, M.C.L.; Rodrigues, M.; Goncalves, J.L.M.; Pavinato, P.S.; Foltran, E.C.; Harrison, R.; James, J.N. 2019b. Impacts of timber harvest intensity and P fertilizer application on soil P fractions. For. Ecol. Manag. 2019, 437, 295–303. [Google Scholar] [CrossRef]
- Siregar, S.T.H.; Nurwahyudi; Mulawarman. Effects of inter-rotation management on site productivity of Acacia mangium in Riau Province, Sumatra, Indonesia. In Site Management and Productivity in Tropical Plantation Forests, Proceedings of the Workshops, Piracicaba, Brazil, 22–26 November 2004; Bogor, Indonesia, 6–9 November 2006; Nambiar, E.K.S., Ed.; CIFOR: Bogor, Indonesia, 2008; pp. 93–106. [Google Scholar]
- Rocha, J.H.T.; Goncalves, J.d.L.M.; Gava, J.L.; Godinho, T.d.O.; Melo, E.A.S.C.; Bazani, J.H.; Hubner, A.; Arthur Junior, J.C.; Wichert, M.P. Forest residue maintenance increased the wood productivity of a Eucalyptus plantation over two short rotations. For. Ecol. Manag. 2016, 379, 1–10. [Google Scholar] [CrossRef]
- Nzila, J.D.; Bouillet, J.-P.; Laclau, J.P.; Ranger, J. The effects of slash management on nutrient cycling and tree growth in Eucalyptus plantation in the Congo. For. Eco. Manag. 2021, 171, 209–221. [Google Scholar]
- Rocha, J.H.T.; Goncalves, J.L.M.; Ferras, A.V.; Poiati, D.A.; Arthur Junior, J.C.; Hubner, A. Growth dynamics and productivity of an Eucalyptus grandis plantation under omission of N, P, K, Ca and Mg over two crop rotation. For. Ecol. Manag. 2019, 447, 158–168. [Google Scholar] [CrossRef]
- Mendham, D.S.; Ogden, G.N.; Short, T.; O’Connell, T.M.; Grove, T.S.; Rance, S.J. Repeated harvest residue removal reduces E. globulus productivity in the 3rd rotation in south-western Australia. For. Ecol. Manag. 2014, 329, 279–286. [Google Scholar] [CrossRef]
- Epron, D.; Mouanda, C.; Mareschal, L.; Koutika, L.S. 2015. Impacts of organic residue management on the soil C dynamics in a tropical eucalypt plantation on a nutrient-poor sandy after three rotations. Soil Biol. Biochem. 2015, 85, 183–189. [Google Scholar] [CrossRef]
- Wibisono, M.G.; Veneklass, E.; Mendham, E.S.; Hardiyanto, E.B. Nitrogen fixation of Acacia mangium Willd. from two seed sources grown at different levels of phosphorus in an Ultisol, South Sumatra. South. For. 2015, 77, 59–64. [Google Scholar] [CrossRef]
- Mendham, D.S.; White, D.A. A review of nutrient, water and organic matter dynamics of tropical acacias on mineral soils for improved management in Southeast Asia. Aust. For. 2019, 82, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, J.L.M.; Stape, J.L.; Laclau, J.-P.; Bouillet, J.-P.; Ranger, J. Assessing the effects of silvicultural management on long-term site productivity of fast-growing eucalyptus plantations: The Brazilian experience. South. For. 2008, 70, 105–118. [Google Scholar] [CrossRef]
- Cook, R.L.; Binkley, D.; Stape, J.L. Eucalyptus plantation effects on soil carbon after 20 years and three rotations in Brazil. For. Ecol. Manag. 2016, 359, 92–98. [Google Scholar] [CrossRef]
- Foltran, E.C.; Rocha, J.H.T.; Bazani, J.H.; Goncalves, J.L.D.; Rodrigues, M.; Pavinato, P.; Valduga, G.R.; Erro, J.; Garcia-Mina, J.M. Phosphorus poll responses under different P inorganic fertilizers for a eucalyptus plantation in a loamy Oxisol. For. Ecol. Manag. 2019, 435, 170–179. [Google Scholar] [CrossRef]
- Nambiar, E.K.S.; Fife, D. Nutrient retranslocation in temperate confers. Tree Physiol. 1991, 9, 185–207. [Google Scholar] [CrossRef]
- Saur, E.; Nambiar, E.K.S.; Fife, D.N. Foliar nutrient retranslocation in Eucalyptus globulus. Tree Physiol. 2000, 20, 1105–1112. [Google Scholar] [CrossRef] [PubMed]
- Laclau, J.P.; Deleporte, P.; Ranger, J.; Bouillet, J.P.; Kazotti, G. Nutrient dynamics throughout the rotation of Eucalyptus clonal stands in Congo. Ann. Bot. 2003, 91, 879–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inail, A.M.; Hardiyanto, E.B. Growth of Eucalyptus pellita under different harvest residue management after three rotations of Acacia mangium at PT. Musi Hutan Persada, South Sumatra. Tech. Notes R D PT. Musi Hutan Persada 2021, 30, 1–9. [Google Scholar]
- Mendham, D.S.; Hardiyanto, E.B. Realising Genetic Gains in Indonesia and Australian Plantations through Water and Nutrient Management; Final Report of ACIAR Project FST/2004/058; Australian Centre for Agricultural Research: Canberra, Australia, 2011; 88p.
- Eze, P.N.; Udeigwe, T.K.; Meandows, M.E. Plinthite and its associated evolutionary forms in soils and landscapes: A review. Pedosphere 2014, 24, 153–166. [Google Scholar] [CrossRef]
- Inail, A.M. Growth of Eucalyptus pellita grown in different stocking densities at two contrasting sites at PT. Musi Hutan Persada, South Sumatra. Tech. Notes R D PT. Musi Hutan Persada 2021, 31, 1–7. [Google Scholar]
- Inail, A.M. Growth of Eucalyptus pellita clones grown at four different sites at PT. Musi Hutan Persada, South Sumatra-A clone-site interaction trial. Tech. Notes R&D PT. Musi Hutan Persada 2021, 31, 1–8. [Google Scholar]
- Mendham, D.; Rimbawanto, A.; Mohammed, C.; Glen, M.; Hardie, M.; Beadle, C.; Greenhill, M. Increasing Productivity of and Profitability of Indonesian Smallholder Plantation; Final Report of ACIAR Project FST/2009/051; Australian Centre for Agricultural Research: Canberra, Australia, 2015; 129p.
- Directorate General of Planning (MOEF). Report on the Performance of the Ministry of Environment and Forestry in 2019; Laporan Kinerja 2019; Ministry of Environment and Forestry of Indonesia: Jakarta, Indonesia, 2020; 167p. (In Indonesian)
- Rahman, S.A.; Sunderland, T.; Roshetko, J.M.; Basuki, I.; Healy, J. Tree culture of smallholder farmers practicing agroforestry in Gunung Salak, West Java, Indonesia. Small Scale For. 2016, 15, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Nadalia, D.; Sutandi, A.; Nugroho, B. Suitability criteria of land characteristics related to Eucalyptus pellita production. In Proceedings of the International e-Conference on Sustainable Agriculture and Farming System, Bogor, Indonesia, 24–25 September 2020; Volume 694, p. 012053. [Google Scholar] [CrossRef]
- Figyantika, A.; Mendham, D.S.; Hardie, M.A.; Hardiyanto, E.B.; Hunt, M.A. Productivity benefits from integrating Acacia auriculiformis and agricultural cropping in Java, Indonesia. Agrofor. Syst. 2020, 94, 2109–2123. [Google Scholar] [CrossRef]
- Flanagan, A.C.; Midgely, S.J.; Stevens, P.R.; McWhirter, L. Small-holder-tree farmers and forest certification in South East Asia: Productivity, risk and policies. Aust. For. 2019, 82, 18–28. [Google Scholar] [CrossRef]
- Nambiar, E.K.S. Strengthening Vietnam’s forestry sectors and rural development: Higher productivity, value, and access to fairer markets are needed to support small forest growers. Trees For. People 2021, 3, 100052. [Google Scholar] [CrossRef]
- Nambiar, E.K.S. Forestry for rural development, poverty reduction, and climate change mitigation: We can help more with wood. Aust. For. 2015, 78, 55–64. [Google Scholar] [CrossRef]
- Greenhill, M.; Walker, I.; Mendham, D.; Permadi, D. West Kalimantan industrial plantation scheme: Twenty years on. For. Trees Livelihoods 2017, 26, 215–228. [Google Scholar] [CrossRef]
- Permadi, D.B.; Burton, M.; Pandit, R.; Race, D.; Ma, C.; Mendham, D.; Hardiyanto, E.B. Socio-economic factors affecting the rate of adoption of acacia plantations by smallholders in Indonesia. Land Use Policy 2018, 76, 215–223. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Site | |||||
|---|---|---|---|---|---|---|
| Merbau-3 | Nakau | Merbau-1 | Lengi | Lantingan | Niru | |
| Slope (%) | 3–8 | 3–6 | 30–40 | 3–5 | 5–8 | 8–10 |
| Bulk density (g cm−3) | 1.07 | 1.02 | 1.16 | 0.90 | 1.40 | 1.17 |
| Depth of A-horizon (cm) | 12 | 15 | 14 | 9 | 10 | 3 |
| Depth to plinthite layer (cm) | 109.9 | 83.7 | 79.9 | 70.4 | 66.9 | 34.2 |
| Sand (%) | 56.7 | 44.6 | 46.7 | 31.9 | 40.8 | 37.4 |
| Silt (%) | 22.1 | 31.3 | 27.5 | 32.1 | 40.6 | 36.3 |
| Clay (%) | 21.2 | 24.1 | 25.8 | 35.9 | 18.6 | 16.3 |
| pH (H2O) | 4.3 | 4.2 | 4.2 | 4.3 | 4.1 | 4.2 |
| Total N (%) | 0.12 | 0.15 | 0.12 | 0.23 | 0.14 | 0.18 |
| Extractable P (mg kg−1) | 2.76 | 2.26 | 3.63 | 2.19 | 3.09 | 2.16 |
| Parameter | Acacia mangium | Eucalyptus pellita |
|---|---|---|
| Stocking: planting/harvest (tree ha−1) | 1111/869 | 1666/1005 |
| Height (m) | 26.4 (0.2) | 18.4 (0.3) |
| Diameter (Dbh, cm) | 20.3 (0.9) | 13.0 (0.1) |
| MAI (m3 ha−1 y−1) | 50.0 (1.5) | 34.9 (0.7) |
| Commercial wood, diameter > 5 cm (m3 ha−1) | 288.3 (8.8) | 178.3 (3.3) |
| Standing biomass (Mg ha−1) | 171.6 (4.9) | 131.9 (3.0 |
| Mean litterfall (Mg ha−1y−1) | 10.0 (0.4) | 8.7 (0.4) |
| Rain during the rotation (mm y−1) | 2570 | 2848 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hardiyanto, E.B.; Inail, M.A.; Nambiar, E.K.S. Productivity of Eucalyptus pellita in Sumatra: Acacia mangium Legacy, Response to Phosphorus, and Site Variables for Guiding Management. Forests 2021, 12, 1186. https://doi.org/10.3390/f12091186
Hardiyanto EB, Inail MA, Nambiar EKS. Productivity of Eucalyptus pellita in Sumatra: Acacia mangium Legacy, Response to Phosphorus, and Site Variables for Guiding Management. Forests. 2021; 12(9):1186. https://doi.org/10.3390/f12091186
Chicago/Turabian StyleHardiyanto, Eko B., Maydra A. Inail, and E. K. Sadanandan Nambiar. 2021. "Productivity of Eucalyptus pellita in Sumatra: Acacia mangium Legacy, Response to Phosphorus, and Site Variables for Guiding Management" Forests 12, no. 9: 1186. https://doi.org/10.3390/f12091186
APA StyleHardiyanto, E. B., Inail, M. A., & Nambiar, E. K. S. (2021). Productivity of Eucalyptus pellita in Sumatra: Acacia mangium Legacy, Response to Phosphorus, and Site Variables for Guiding Management. Forests, 12(9), 1186. https://doi.org/10.3390/f12091186