
The Spatial Pattern of the Upper Limit of Montane Deciduous Broad-Leaved Forests and Its Geographical Interpretation in the East Monsoon Realm of China


Abstract
:1. Introduction
2. Data and Methods
2.1. Data Sets for MDB
2.2. Environmental Data
2.3. Statistical Analyses
3. Results
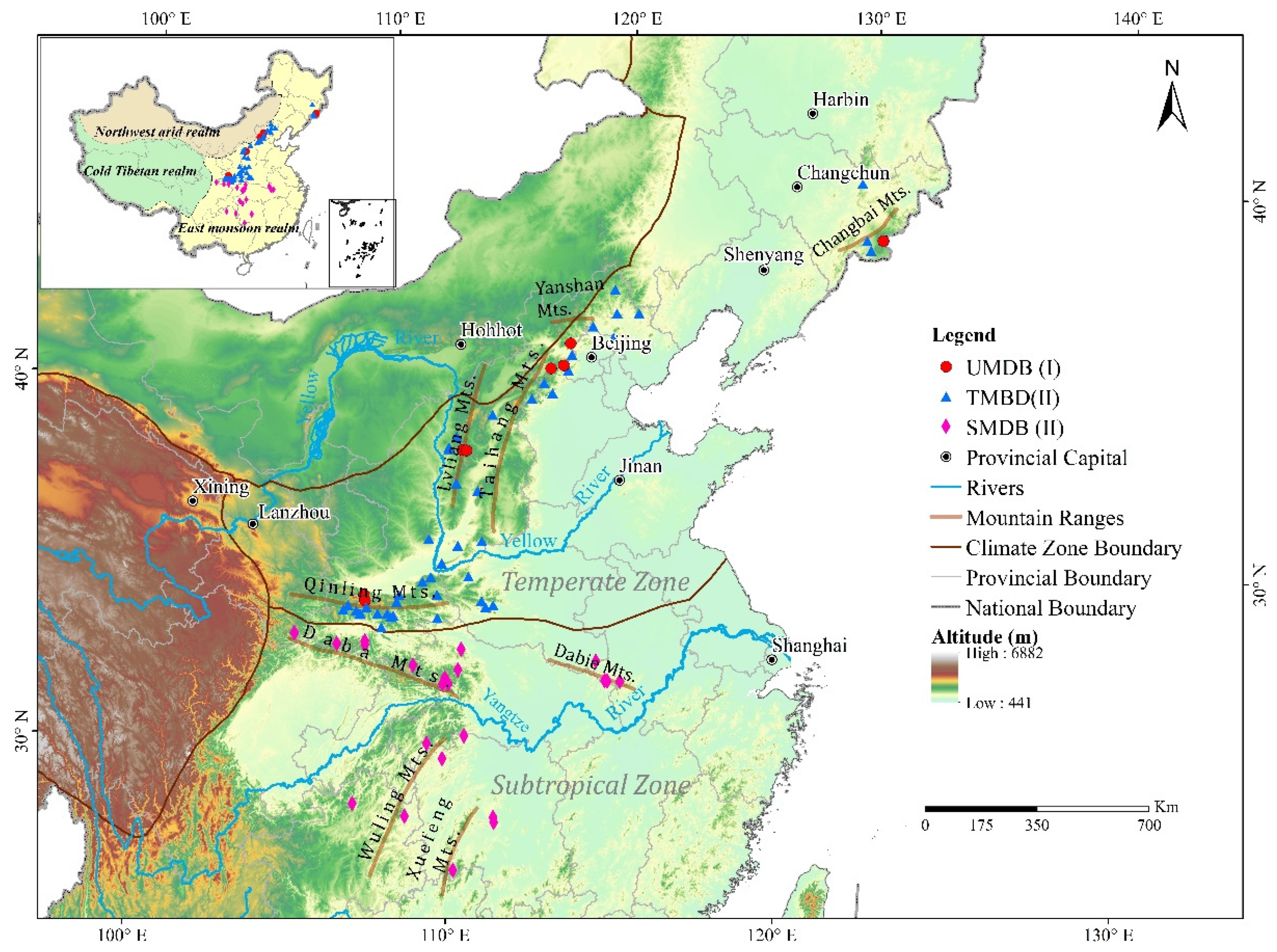
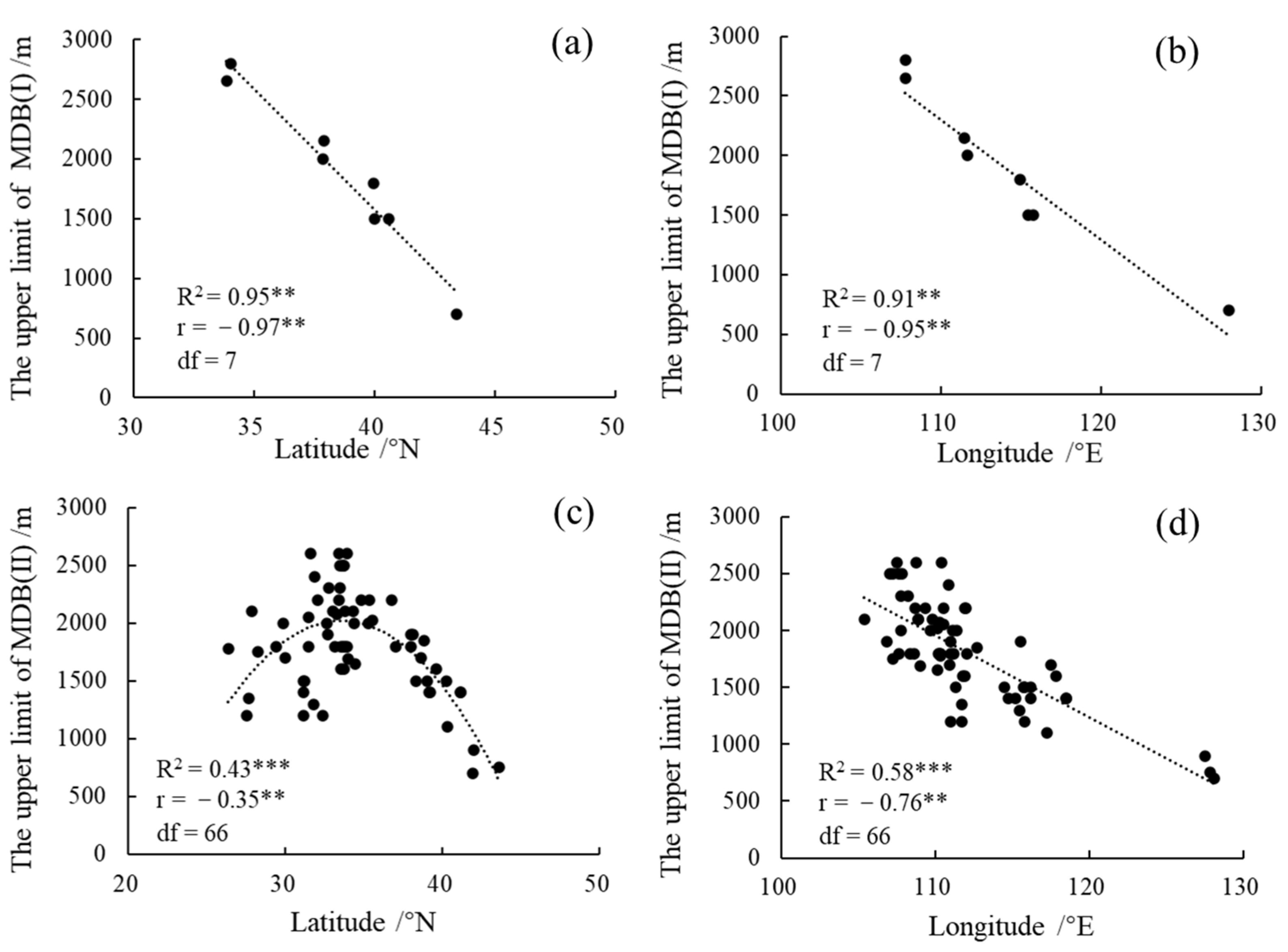
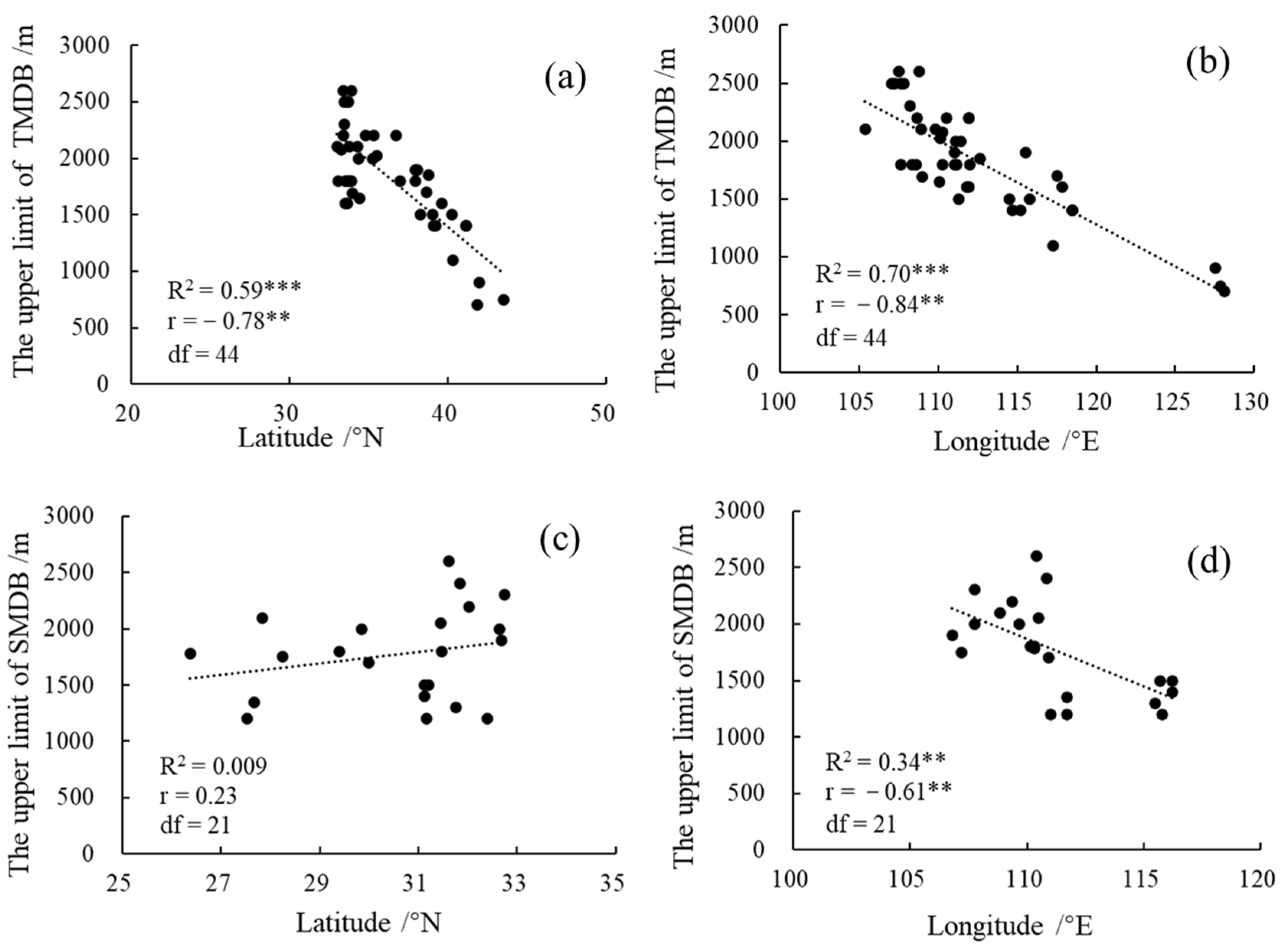
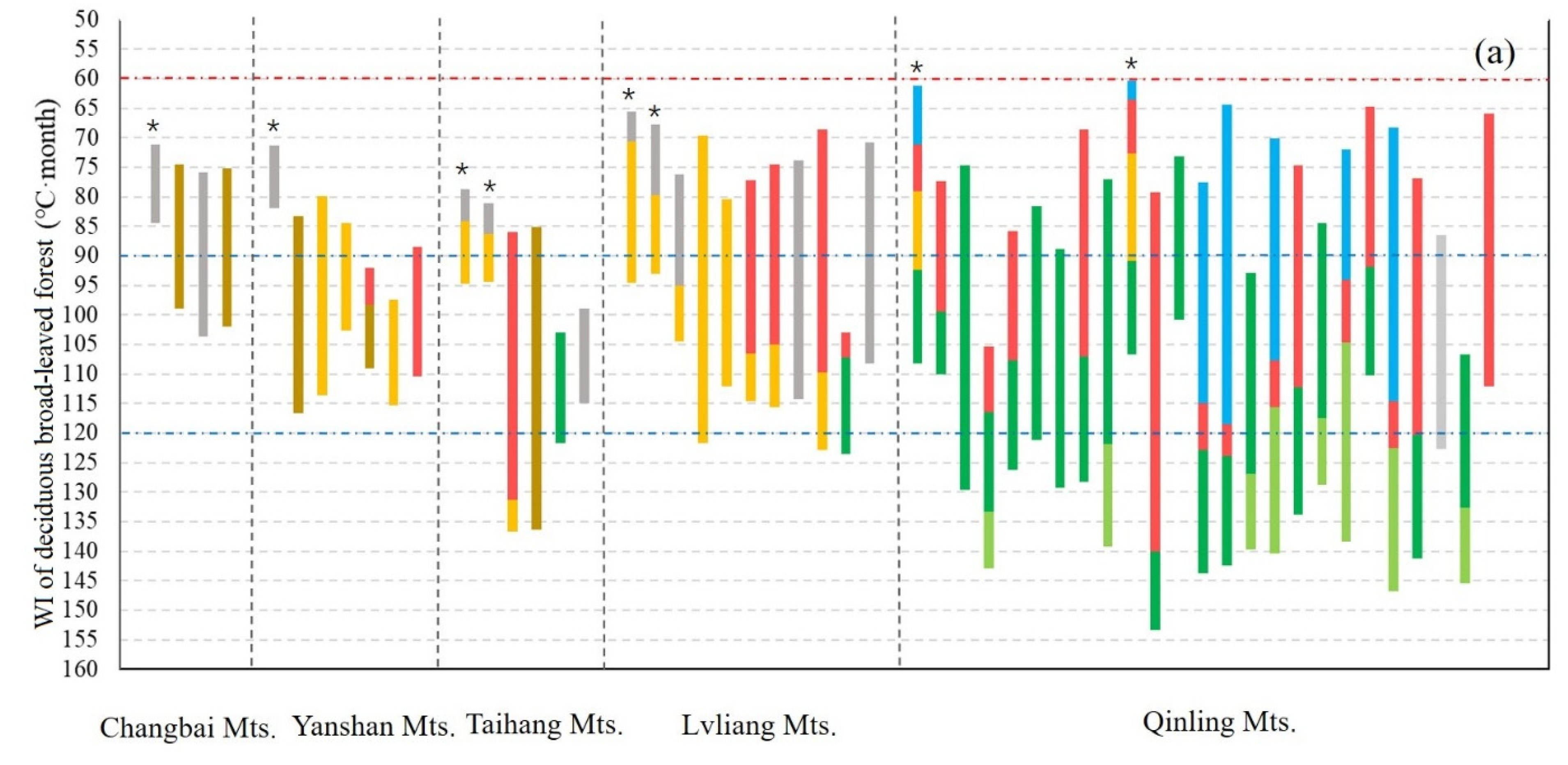
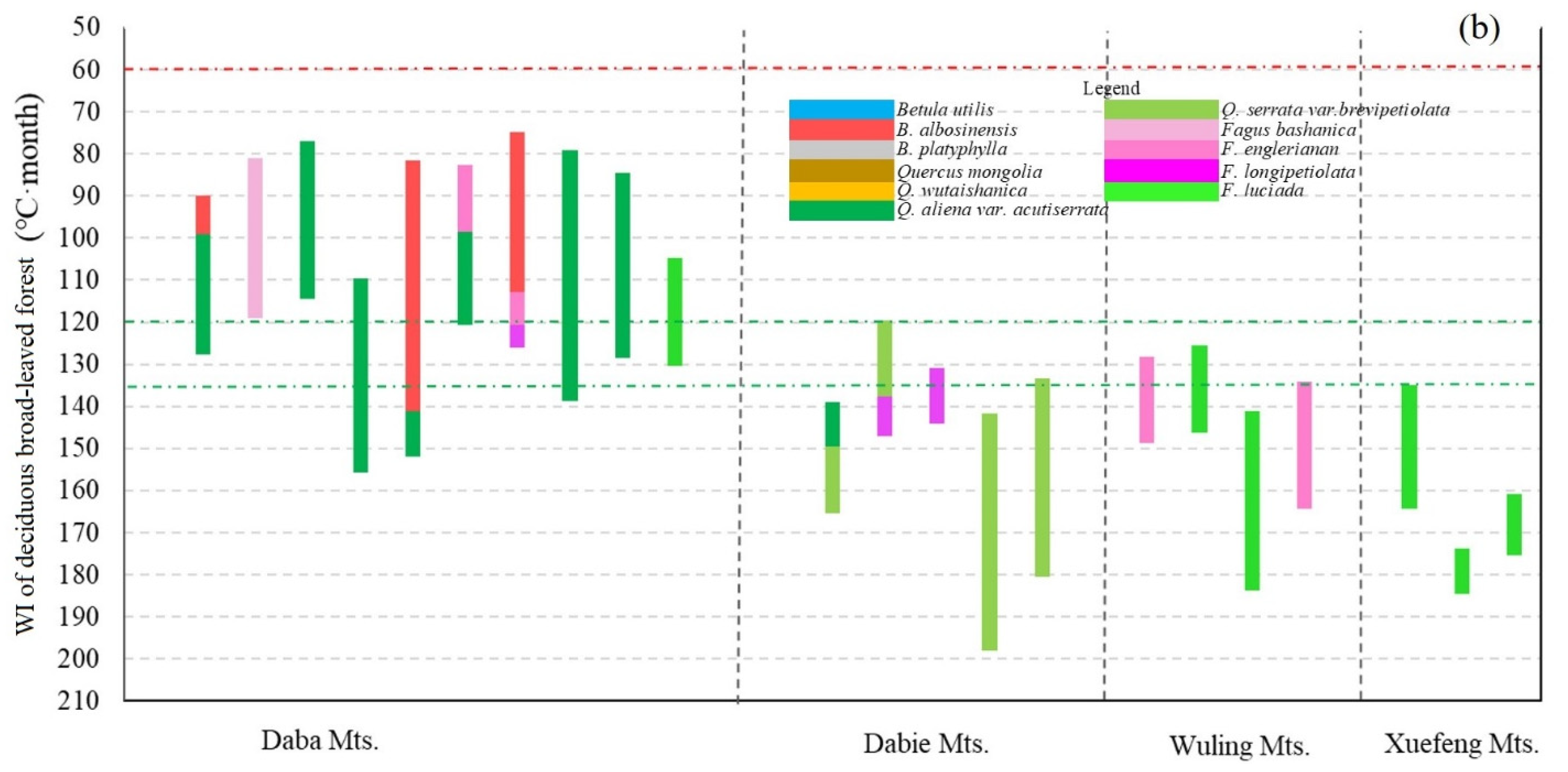
3.1. Geographical Distribution of the Upper Limit of MDB in the East Realm of China
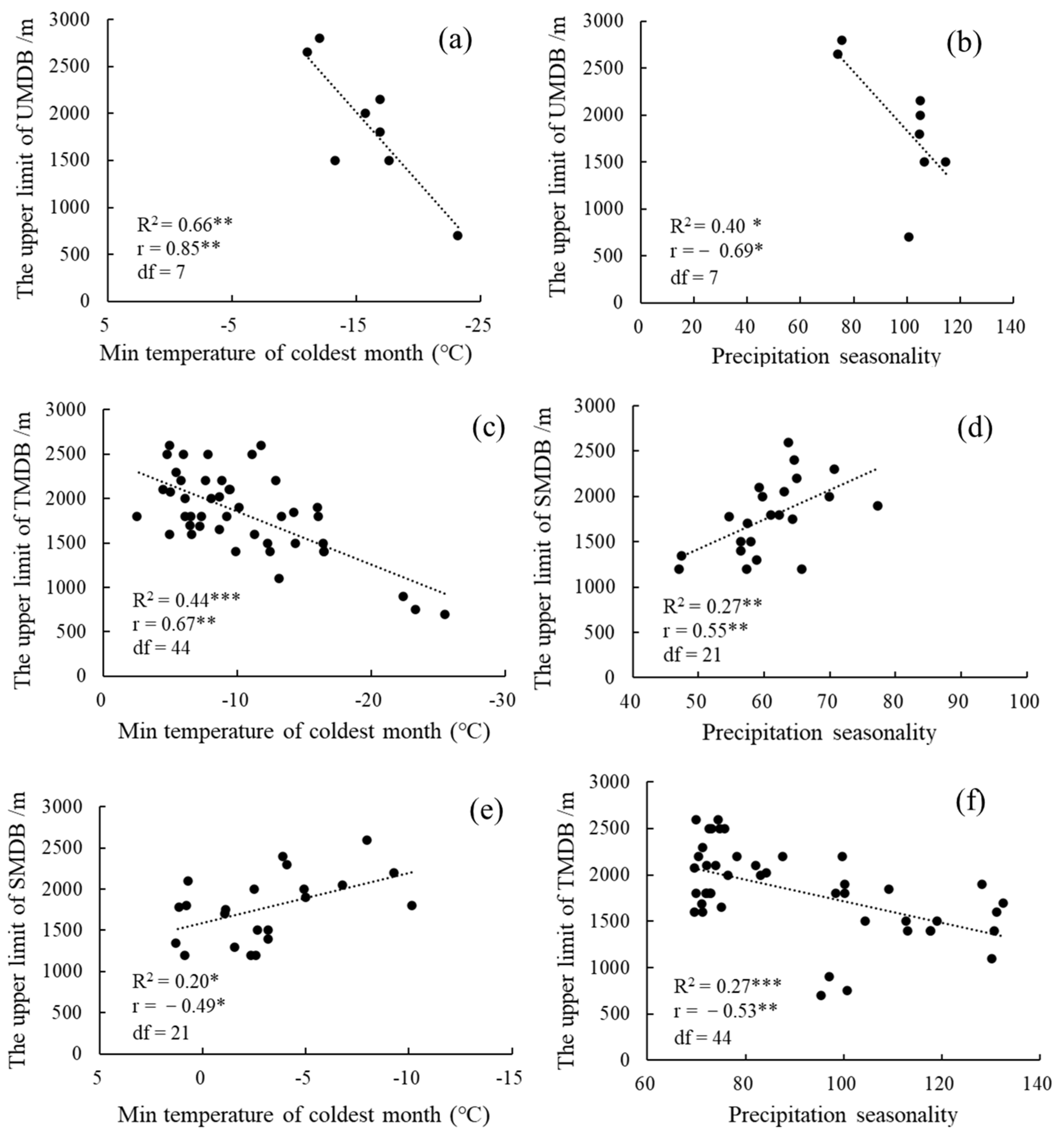
3.2. The Influence of Climate Factors on the Upper Limit of MDB
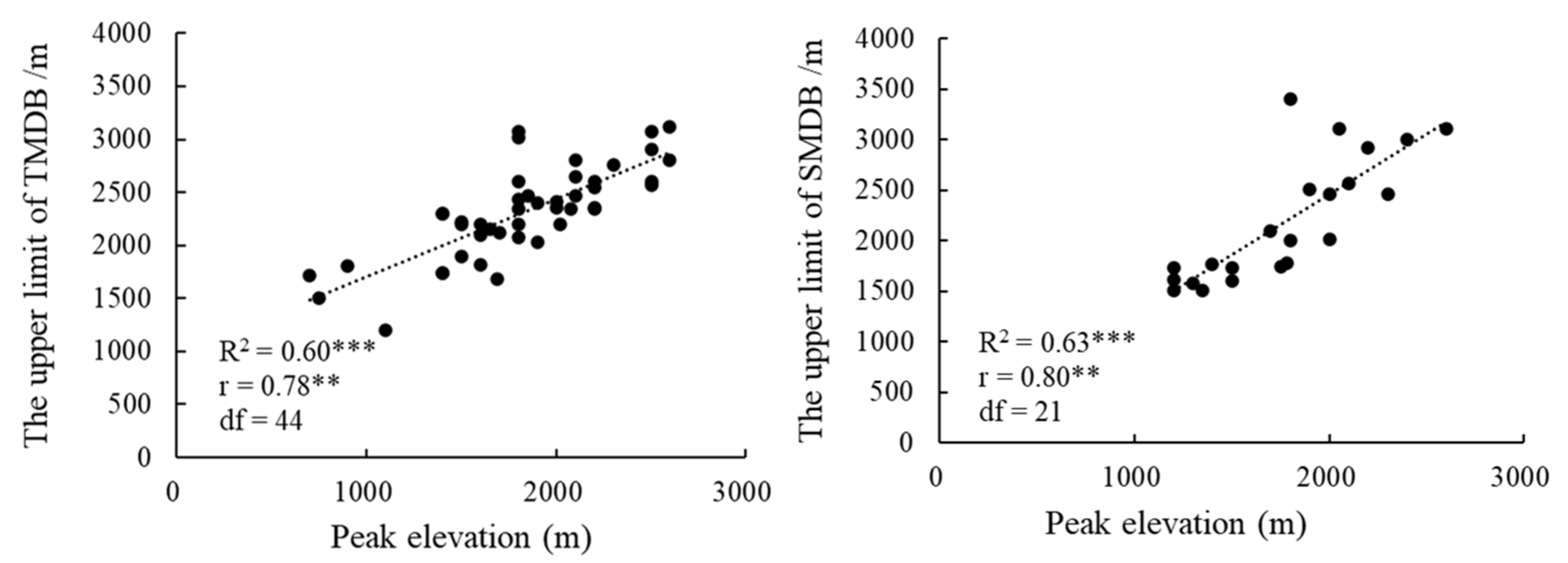
3.3. The Influence of Peak Elevation on the Upper Limit of MDB
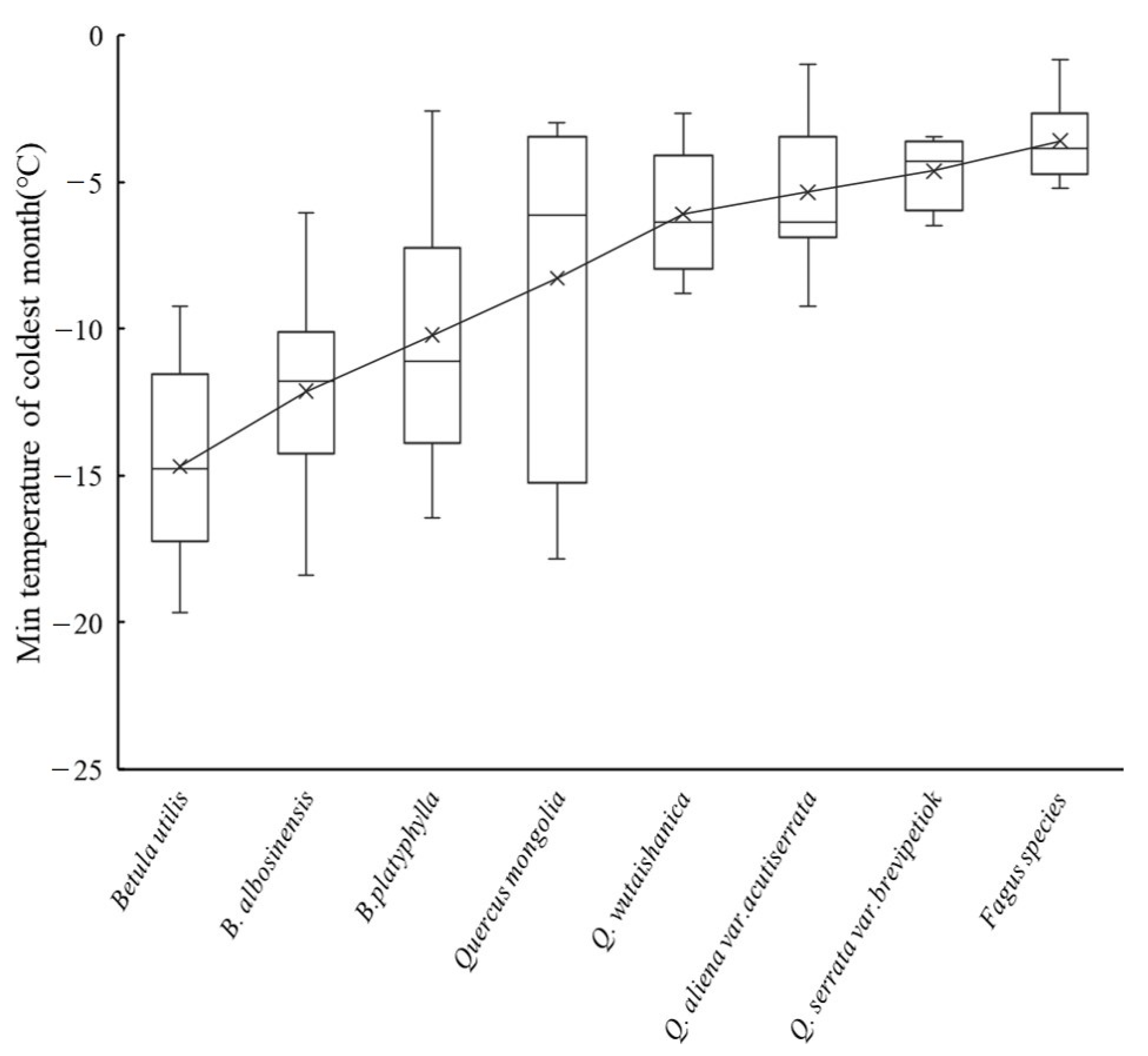
3.4. The Influence of the Cold Tolerance of Dominant Species on the Upper Limit of MDB
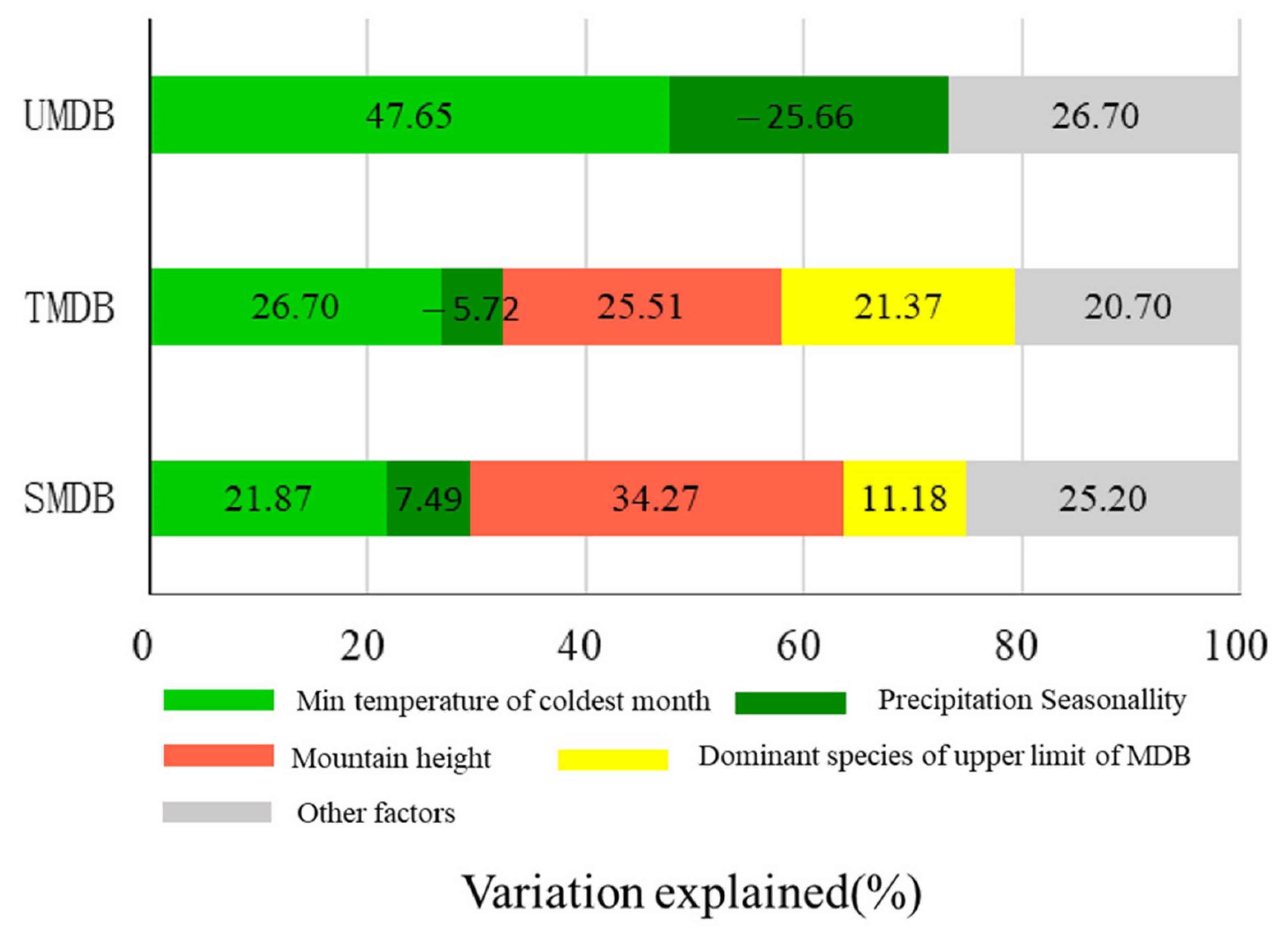
3.5. The Relative Contribution of Environmental Factors to the Upper Limit of MDB
4. Discussion
4.1. Dominant Factors Influencing the Upper Limit of Deciduous Broad-Leaved Forest on the Mountains with or without Climatic Treeline
4.2. Dominant Factors Influencing the Upper Limit of MDB on Temperate and Subtropical Mountains without Climatic Treeline
4.3. Difference of Dominant Species for the Upper Limit of Deciduous Broad-Leaved Forest on the Temperate and Subtropical Mountains
4.4. WI of the Upper Limit of MDB in the East Monsoon Realm of China
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, Z.Y. Physical Geography of China: Plant Geography; Science Press: Beijing, China, 1983; pp. 20–45. [Google Scholar]
- Wen, D. Carbon Sequestration in Chinese Forest Ecosystems: Pattern, Potential, and Controlling Mechanisms; University of Chinese Academy of Sciences: Beijing, China, 2016; pp. 4–14. [Google Scholar]
- Rahmat, A. Hydrological Characteristics under Deciduous Broadleaf and Evergreen Coniferous Forests in Central Japan. Int. J. Geomate 2019, 16, 217–224. [Google Scholar] [CrossRef]
- Dai, J.H.; Cui, H.T. A review on the studies of alpine timberline. Sci. Geogr. Sin. 1999, 19, 243–249. [Google Scholar]
- Ohsawa, M. An Interpretation of Latitudinal Patterns of Forest Limits in South and East Asian Mountains. J. Ecol. 1990, 78, 326–339. [Google Scholar] [CrossRef]
- Irl, S.D.H.; Anthelme, F.; Harter, D.E.V.; Jentsch, A.; Lotter, E.; Steinbauer, M.J.; Beierkuhnlein, C. Patterns of island treeline elevation—A global perspective. Ecography 2015, 39, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.L.; Guo, R.Q.; Sun, S.C. Variations in the vertical vegetation zonation of subtropical Chinese mountains: The importance of climatic seasonality. Acta Ecol. Sin. 2010, 30, 3912–3922. [Google Scholar]
- Xu, H.C. Natural Forests in North China; China Science and Technology Press: Beijing, China, 2003; pp. 1–10. [Google Scholar]
- Chen, L.Z. Flora and Vegetation Geography of China; Science Press: Beijing, China, 2014; pp. 344–349. [Google Scholar]
- Rumpf, S.B.; Hülber, K.; Klonner, G.; Moser, D.; Schütz, M.; Wessely, J.; Willner, W.; Zimmermann, N.E.; Dullinger, S. Range dynamics of mountain plants decrease with elevation. Proc. Natl. Acad. Sci. USA 2018, 115, 1848–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, M.R.F.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Körner, C.; Basler, D.; Hoch, G.; Kollas, C.; Lenz, A.; Randin, C.F.; Vitasse, Y.; Zimmermann, N. Where, why and how? Explaining the low-temperature range limits of temperate tree species. J. Ecol. 2016, 104, 1076–1088. [Google Scholar] [CrossRef]
- Sakai, A. Freezing tolerance of evergreen and deciduous broad-leaved trees in Japan with reference to tree regions. Low Temp. Sci. Ser. B Biol. Sci. 1979, 30, 1–9. [Google Scholar]
- Sakai, A.; Larcher, P.D.W. Frost Survival of Plants; Springer Science & Business Media: Berlin, Germany, 1987; pp. 62–85. [Google Scholar] [CrossRef]
- Lenz, A.; Hoch, G.; Vitasse, Y.; Körner, C. European deciduous trees exhibit similar safety margins against damage by spring freeze events along elevational gradients. New Phytol. 2013, 200, 1166–1175. [Google Scholar] [CrossRef]
- Stephenson, N.L. Climatic control of vegetation distribution: The role of the water balance. Am. Nat. 1990, 135, 649–670. [Google Scholar] [CrossRef]
- China Forest Editorial Board. Chinese Forest (Broadleaved Forest); China Forestry Press: Beijing, China, 2000; pp. 1838–1875. [Google Scholar]
- Ge, J. Climatic seasonality is linked to the occurrence of the mixed evergreen and deciduous broad-leaved forests in China. Ecosphere 2019, 10, e02862. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer Science & Business Media: Berlin, Germany, 2012; pp. 23–25. [Google Scholar]
- Odland, A. Effect of latitude and mountain height on the timberline (Betula pubescens ssp. czerpanovii) elevation along the central Scandinavian mountain range. Fenn. Int. J. Geogr. 2015, 193, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.P.; Tan, J.; Yao, Y.H. Digital Integration and Patterns of Mountains Altitudinal Belts; China Environmental Science Press: Beijing, China, 2009; pp. 200–223. [Google Scholar]
- Zhang, B.P.; Yao, Y.H. Studies on Mass Elevation Effect; China Environment Press: Beijing, China, 2015; pp. 9–27. [Google Scholar]
- Fang, J.Y. Rediscussion about the forest vegetation zonation in eastern China. Acta Bot. Sin. 2001, 43, 522–533. [Google Scholar] [CrossRef]
- Kang, M.Y.; Zhu, Y. Discussion and analysis on the geo-ecological boundary in Qinling Range. Acta Ecol. Sin. 2006, 27, 2774–2784. [Google Scholar]
- Jobbágy, E.; Jackson, R. Global controls of forest line elevation in the northern and southern hemisphere. Glob. Ecol. Biogeogr. 2000, 9, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Ghalambor, C.K.; Huey, R.B.; Martin, P.R.; Tewksbury, J.J.; Wang, G. Are mountain passes higher in the tropics? Janzen’s hypothesis revisited. Integr. Comp. Biol. 2006, 46, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Clim. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Elsen, P.R.; Monahan, W.B.; Merenlender, A.M. Global patterns of protection of elevational gradients in mountain ranges. Proc. Natl. Acad. Sci. USA 2018, 115, 6004–6009. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.Y.; Yoda, K. Climate and vegetation in China (I). Changes in the altitudinal lapse rate of temperature and distribution of sea-level temperature. Ecol. Res. 1988, 3, 37–51. [Google Scholar] [CrossRef]
- Peng, B.Z.; Chen, F. Progress in the study of vertical mountain zonation in China. Sci. Geogr. Sin. 1999, 4, 303–308. [Google Scholar] [CrossRef]
- Sun, J. Mountain altitudinal belt: A review. Ecol. Environ. Sci. 2014, 23, 1544–1550. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales: Treeline and environmental change. Glob. Ecol. Biogeogr. 2005, 14, 395–410. [Google Scholar] [CrossRef]
- Grabherr, G.; Nagy, L.; Thompson, D. An Outline of Europe’s Alpine Areas. In Alpine Biodiversity in Europe; Springer: Berlin/Heidelberg, Germany, 2003; pp. 3–12. [Google Scholar] [CrossRef]
- Zhang, Q.; Du, J.H.; Tian, H.; Huang, Y.X. The temporal-spatial characteristics of the pseudo-timberline in Shimentain National Nature Reserve and its adjacent areas based on 3S technology. Trop. Geogr. 2018, 38, 364–371. [Google Scholar] [CrossRef]
- Lei, M.D. Vegetation in Shanxi, China; Science Press: Beijing, China, 1999; pp. 410–440. [Google Scholar]
- Wang, X.P.; Zhang, L.; Fang, J.Y. Relationship between alpine timberline and climate in China. Acta Geogr. Sin. 2004, 59, 871–879. [Google Scholar]
- Li, Z.G.; Dang, K.L.; Li, D.W. Integrated Scientific Investigation of Tianhua Mountains in Shanxi Province; Shanxi Science and Technology Press: Shanxi, China, 2005; pp. 426–450. [Google Scholar]
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 261–280. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, S.; Di Biase, L.; Chiarucci, A. Recognizing and interpreting vegetational belts: New wine in the old bottles of a von Humboldt’s legacy. J. Biogeogr. 2019, 46, 1643–1651. [Google Scholar] [CrossRef]
- Woodward, F.I.; Williams, B.G. Climate and plant distribution at global and local scales. Vegetatio 1987, 69, 189–197. [Google Scholar] [CrossRef]
- Shen, Z.-H.; Fang, J.-Y.; Chiu, C.-A.; Chen, T.-Y.; Liu, H. The geographical distribution and differentiation of Chinese beech forests and the association with Quercus. Appl. Veg. Sci. 2014, 18, 23–33. [Google Scholar] [CrossRef]
- Amissah, L.; Mohren, G.M.J.; Bongers, F.; Hawthorne, W.D.; Poorter, L. Rainfall and temperature affect tree species distribution in Ghana. J. Trop. Ecol. 2014, 30, 435–446. [Google Scholar] [CrossRef]
- Fang, J.Y.; Guo, Q.H.; Liu, G.H. Distribution patterns of Chinese Beech (Fagus L.) species in relation to topography. Acta Bot. Sin. 1999, 41, 766–774. [Google Scholar] [CrossRef]
- Fang, J.; Lechowicz, M.J. Climatic limits for the present distribution of beech (fagus L.) species in the world. J. Biogeogr. 2006, 33, 1804–1819. [Google Scholar] [CrossRef]
- Cai, Q.; Welk, E.; Ji, C.; Fang, W.; Sabatini, F.M.; Zhu, J.; Zhu, J.; Tang, Z.; Attorre, F.; Campos, J.A.; et al. The relationship between niche breadth and range size of beech (Fagus) species worldwide. J. Biogeogr. 2021, 48, 1240–1253. [Google Scholar] [CrossRef]
- Hong, B.G.; An, S.Q. Preliminary studies on the geographical distribution of Fagus in China. J. Integr. Plant Biol. 1993, 35, 229–233. [Google Scholar]
- Guo, K.; Werger, M.J.A. Effect of prevailing monsoons on the distribution of beeches in continental East Asia. For. Ecol. Manag. 2010, 259, 2197–2203. [Google Scholar] [CrossRef]
- Wu, Z.Y. Vegetation of China; Science Press: Beijing, China, 1980; pp. 250–280. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DV | Model | Adjusted R2 | ΔR2 | ΔF | Sig. ΔF | Variables | B | β | T | p |
|---|---|---|---|---|---|---|---|---|---|---|
| UMDB(I) | 1 | 0.733 | 0.809 | 10.619 | 0.016 | Intercept | 883.075 | 6.035 | 0.002 ** | |
| MinT | 40.224 | 0.663 | 2.914 | 0.033 * | ||||||
| PS | −10.266 | −0.357 | −1.571 | 0.177 | ||||||
| TMDB(II) | 1 | 0.477 | 0.501 | 21.084 | <0.001 | Intercept | 2827.668 | 12.815 | <0.001 *** | |
| MinT | 47.523 | 0.538 | 4.206 | <0.001 *** | ||||||
| PS | −5.588 | −0.257 | −2.008 | 0.051 | ||||||
| 2 | 0.717 | 0.235 | 36.492 | <0.001 | Intercept | 801.344 | 2.150 | 0.037 * | ||
| MinT | 34.165 | 0.387 | 3.969 | <0.001 *** | ||||||
| PS | −0.713 | −0.033 | −0.324 | 0.748 | ||||||
| H | 0.624 | 0.586 | 6.041 | <0.001 *** | ||||||
| 3 | 0.793 | 0.076 | 16.217 | <0.001 | Intercept | 935.613 | 2.924 | <0.006 ** | ||
| MinT | 35.860 | 0.406 | 4.871 | <0.001 *** | ||||||
| PS | −1.882 | −0.087 | −0.989 | 0.329 | ||||||
| H | 0.413 | 0.388 | 4.017 | <0.001 *** | ||||||
| RCT | 84.293 | 0.325 | 4.027 | <0.001 *** | ||||||
| SMDB(II) | 1 | 0.289 | 0.357 | 5.265 | 0.015 | Intercept | 214.362 | 0.286 | 0.778 | |
| MinT | −32.936 | −0.265 | −1.201 | 0.245 | ||||||
| PS | 23.959 | 0.408 | 1.849 | 0.080 | ||||||
| 2 | 0.689 | 0.377 | 25.401 | <0.001 | Intercept | −416.691 | −0.815 | 0.426 | ||
| MinT | 51.103 | 0.411 | 2.074 | 0.053 * | ||||||
| PS | 14.449 | 0.246 | 1.646 | 0.117 | ||||||
| H | 0.669 | 0.991 | 5.040 | <0.001 *** | ||||||
| 3 | 0.748 | 0.063 | 5.249 | 0.035 | Intercept | −491.184 | −1.065 | 0.302 | ||
| MinT | 82.399 | 0.663 | 3.165 | 0.006 ** | ||||||
| PS | 13.306 | 0.227 | 1.682 | 0.111 | ||||||
| H | 0.702 | 1.039 | 5.834 | <0.001 *** | ||||||
| RCT | 68.807 | 0.339 | 2.291 | 0.035 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhang, B.; Yao, Y. The Spatial Pattern of the Upper Limit of Montane Deciduous Broad-Leaved Forests and Its Geographical Interpretation in the East Monsoon Realm of China. Forests 2021, 12, 1225. https://doi.org/10.3390/f12091225
Wang J, Zhang B, Yao Y. The Spatial Pattern of the Upper Limit of Montane Deciduous Broad-Leaved Forests and Its Geographical Interpretation in the East Monsoon Realm of China. Forests. 2021; 12(9):1225. https://doi.org/10.3390/f12091225
Chicago/Turabian StyleWang, Jing, Baiping Zhang, and Yonghui Yao. 2021. "The Spatial Pattern of the Upper Limit of Montane Deciduous Broad-Leaved Forests and Its Geographical Interpretation in the East Monsoon Realm of China" Forests 12, no. 9: 1225. https://doi.org/10.3390/f12091225