
A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data
2.2. Indicator Development
3. Results
3.1. Forest-Associated Species Potentially at Risk of Losing Genetic Variation
3.2. Forest-Associated Species at Potential Risk of Losing Locally Adapted Genotypes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potter, K.M.; Jetton, R.M.; Bower, A.; Jacobs, D.F.; Man, G.; Hipkins, V.D.; Westwood, M. Banking on the future: Progress, challenges and opportunities for the genetic conservation of forest trees. New For. 2017, 48, 153–180. [Google Scholar] [CrossRef]
- Carroll, S.P.; Jorgensen, P.S.; Kinnison, M.T.; Bergstrom, C.T.; Denison, R.F.; Gluckman, P.; Smith, T.B.; Strauss, S.Y.; Tabashnik, B.E. Applying evolutionary biology to address global challenges. Science 2014, 346, 1245993-1–1245993-10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, A.A.; Sgro, C.M.; Kristensen, T.N. Revisiting adaptive potential, population size, and conservation. Trends Ecol. Evol. 2017, 32, 506–517. [Google Scholar] [CrossRef]
- Raffard, A.; Santoul, F.; Cucherousset, J.; Blanchet, S. The community and ecosystem consequences of intraspecific diversity: A meta-analysis. Biol. Rev. 2019, 94, 648–661. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Des Roches, S.; Pendleton, L.H.; Shapiro, B.; Palkovacs, E.P. Conserving intraspecific variation for nature’s contributions to people. Nat. Ecol. Evol. 2021, 5, 574–582. [Google Scholar] [CrossRef]
- Stange, M.; Barrett, R.D.H.; Hendry, A.P. The importance of genomic variation for biodiversity, ecosystems and people. Nat. Rev. Genet. 2021, 22, 89–105. [Google Scholar] [CrossRef]
- Leigh, D.M.; Hendry, A.P.; Vazquez-Dominguez, E.; Friesen, V.L. Estimated six per cent loss of genetic variation in wild populations since the industrial revolution. Evol. Appl. 2019, 12, 1505–1512. [Google Scholar] [CrossRef] [Green Version]
- FAO. The State of the World’s Forest Genetic Resources; Commission on Genetic Resources for Food and Agriculture, Food and Agriculture Organization of the United Nations: Rome, Italy, 2014; p. 276. [Google Scholar]
- Hamrick, J.L. Response of forest trees to global environmental changes. For. Ecol. Manag. 2004, 197, 323–335. [Google Scholar] [CrossRef]
- Alfaro, R.I.; Fady, B.; Vendramin, G.G.; Dawson, I.K.; Fleming, R.A.; Saenz-Romero, C.; Lindig-Cisneros, R.A.; Murdock, T.; Vinceti, B.; Navarro, C.M.; et al. The role of forest genetic resources in responding to biotic and abiotic factors in the context of anthropogenic climate change. For. Ecol. Manag. 2014, 333, 76–87. [Google Scholar] [CrossRef]
- Zhu, K.; Woodall, C.W.; Clark, J.S. Failure to migrate: Lack of tree range expansion in response to climate change. Glob. Chang. Biol. 2012, 18, 1042–1052. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Liebhold, A.M. Ecology of forest insect invasions. Biol. Invasions 2017, 19, 3141–3159. [Google Scholar] [CrossRef]
- Ghelardini, L.; Luchi, N.; Pecori, F.; Pepori, A.L.; Danti, R.; Della Rocca, G.; Capretti, P.; Tsopelas, P.; Santini, A. Ecology of invasive forest pathogens. Biol. Invasions 2017, 19, 3183–3200. [Google Scholar] [CrossRef]
- Potter, K.M.; Escanferla, M.E.; Jetton, R.M.; Man, G. Important insect and disease threats to United States tree species and geographic patterns of their potential impacts. Forests 2019, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Schaberg, P.G.; DeHayes, D.H.; Hawley, G.J.; Nijensohn, S.E. Anthropogenic alterations of genetic diversity within tree populations: Implications for forest ecosystem resilience. For. Ecol. Manag. 2008, 256, 855–862. [Google Scholar] [CrossRef]
- Jump, A.S.; Marchant, R.; Penuelas, J. Environmental change and the option value of genetic diversity. Trends Plant Sci. 2009, 14, 51–58. [Google Scholar] [CrossRef]
- Forest Europe. State of Europe’s Forests 2015. In Proceedings of the Ministerial Conference on the Protection of Forests in Europe, Madrid, Spain, 20–21 October 2015; p. 312. [Google Scholar]
- Montréal Process Liaison Office. The Montréal Process Criteria and Indicators for the Conservation and Sustainable Management of Temperate and Boreal Forests, 5th ed.; Available online: http://web.archive.org/web/20160211053015/http://montrealprocess.org/documents/publications/techreports/MontrealProcessSeptember2015.pdf (accessed on 10 November 2021).
- United States Department of Agriculture Forest Service. National Report on Sustainable Forests—2010; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2011; p. 134.
- United States Department of Agriculture Forest Service. National Report on Sustainable Forests—2003; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2004; p. 139.
- Graudal, L.; Aravanopoulos, F.; Bennadji, Z.; Changtragoon, S.; Fady, B.; Kjaer, E.D.; Loo, J.; Ramamonjisoa, L.; Vendramin, G.G. Global to local genetic diversity indicators of evolutionary potential in tree species within and outside forests. For. Ecol. Manag. 2014, 333, 35–51. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Leites, L.P.; St Clair, J.B.; Jaquish, B.C.; Saenz-Romero, C.; Lopez-Upton, J.; Joyce, D.G. Comparative responses to climate in the varieties of Pinus ponderosa and Pseudotsuga menziesii: Clines in growth potential. For. Ecol. Manag. 2014, 324, 138–146. [Google Scholar] [CrossRef]
- Tripiana, V.; Bourgeois, M.; Verhaegen, D.; Vigneron, P.; Bouvet, J.M. Combining microsatellites, growth, and adaptive traits for managing in situ genetic resources of Eucalyptus urophylla. Can. J. For. Res.-Rev. Can. Rech. For. 2007, 37, 773–785. [Google Scholar] [CrossRef] [Green Version]
- Boshier, D.H.; Boyle, T.J. Forest conservation genetics: Limitations and future directions. In Forest Conservation Genetics: Principles and Practice; Young, A.G., Boshier, D., Boyle, T.J., Eds.; CSIRO Publishing: Collingwood, Australia, 2000; pp. 289–297. [Google Scholar]
- Willyard, A.; Bower, A.; Hipkins, V.; Snelling, J.; DeWoody, J. Genetic diversity and population structure in Chrysolepis chrysophylla (golden chinquapin; Fagaceae): SSRs vs SNPs. Can. J. For. Res. 2020, 50, 788–799. [Google Scholar] [CrossRef]
- Callahan, C.M.; Rowe, C.A.; Ryel, R.J.; Shaw, J.D.; Madritch, M.D.; Mock, K.E. Continental-scale assessment of genetic diversity and population structure in quaking aspen (Populus tremuloides). J. Biogeogr. 2013, 40, 1780–1791. [Google Scholar] [CrossRef] [Green Version]
- Potter, K.M.; Hipkins, V.D.; Mahalovich, M.F.; Means, R.E. Nuclear genetic variation across the range of ponderosa pine (Pinus ponderosa): Phylogeographic, taxonomic, and conservation implications. Tree Genet. Genomes 2015, 11, 38. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.H.; Frankham, R. How closely correlated are molecular and quantitative measures of genetic variation? A meta-analysis. Evolution 2001, 55, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Shinneman, D.J.; Means, R.E.; Potter, K.M.; Hipkins, V.D. Exploring climate niches of ponderosa pine (Pinus ponderosa Douglas ex Lawson) haplotypes in the western United States: Implications for evolutionary history and conservation. PLoS ONE 2016, 11, e0151811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, A.M.; Potter, K.M. Macro-scale assessment of demographic and environmental variation within genetically derived evolutionary lineages of eastern hemlock (Tsuga canadensis), an imperiled conifer of the eastern United States. Biodivers. Conserv. 2017, 26, 2223–2249. [Google Scholar] [CrossRef]
- Hoban, S.; Campbell, C.D.; da Silva, J.M.; Ekblom, R.; Funk, W.C.; Garner, B.A.; Godoy, J.A.; Kershaw, F.; MacDonald, A.J.; Mergeay, J.; et al. Genetic diversity is considered important but interpreted narrowly in country reports to the Convention on Biological Diversity: Current actions and indicators are insufficient. Biol. Conserv. 2021, 261, 11. [Google Scholar] [CrossRef]
- Manion, P.D.; Griffin, D.H. Large landscape scale analysis of tree death in the Adirondack Park, New York. For. Sci. 2001, 47, 542–549. [Google Scholar] [CrossRef]
- Pike, C.; Potter, K.M.; Berrang, P.; Crane, B.; Baggs, J.; Leites, L.; Luther, T. New seed-collection zones for the Eastern United States: The Eastern Seed Zone Forum. J. For. 2020, 118, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.A.; Bartow, A.; Gisler, M.; Ward, K.; Young, A.S.; Kaye, T.N. Can an ecoregion serve as a seed transfer zone? Evidence from a common garden study with five native species. Restor. Ecol. 2010, 19, 268–276. [Google Scholar] [CrossRef]
- Potter, K.M.; Crane, B.S.; Hargrove, W.W. A United States national prioritization framework for tree species vulnerability to climate change. New For. 2017, 48, 275–300. [Google Scholar] [CrossRef]
- Potter, K.M.; Escanferla, M.E.; Jetton, R.M.; Man, G.; Crane, B.S. Prioritizing the conservation needs of United States tree species: Evaluating vulnerability to forest insect and disease threats. Glob. Ecol. Conserv. 2019, 18, 17. [Google Scholar] [CrossRef]
- Bower, A.D.; St Clair, B.; Erickson, V. Generalized provisional seed zones for native plants. Ecol. Appl. 2014, 24, 913–919. [Google Scholar] [CrossRef]
- Cleland, D.T.; Freeouf, J.A.; Keys, J.E.; Nowacki, G.J.; Carpenter, C.A.; McNab, W.H. Ecological Subregions: Sections and Subsections for the conterminous United States. In General Technical Report WO-76; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2007. Available online: https://www.fs.usda.gov/treesearch/pubs/48672 (accessed on 10 November 2021).
- Bailey, R.G. Description of the ecoregions of the United States; U.S. Department of Agriculture: Washington, DC, USA, 1995. Available online: https://www.fs.fed.us/land/ecosysmgmt/ (accessed on 10 November 2021).
- Cleland, D.T.; Avers, P.E.; McNab, W.H.; Jensen, M.E.; Bailey, R.G.; King, T.; Russell, W.E. National hierarchical framework of ecological units. In Ecosystem Management: Applications for Sustainable Forest and Wildlife Resources; Boyce, M.S., Haney, A., Eds.; Yale University Press: New Haven, CT, USA, 1997; pp. 181–200. [Google Scholar]
- Smith, W.B. Forest inventory and analysis: A national inventory and monitoring program. Environ. Pollut. 2002, 116, S233–S242. [Google Scholar] [CrossRef]
- Bechtold, W.A.; Patterson, P.L. The Enhanced Forest Inventory and Analysis Program: National Sampling Design and Estimation Procedures; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2005; p. 85.
- Burrill, E.A.; Wilson, A.M.; Turner, J.A.; Pugh, S.A.; Menlove, J.; Christiansen, G.; Conkling, B.L.; David, W. The Forest Inventory and Analysis Database: Database Description and User Guide Version 8.0 for Phase 2. Available online: https://www.fia.fs.fed.us/library/database-documentation/current/ver80/FIADB%20User%20Guide%20P2_8-0.pdf (accessed on 16 November 2021).
- Pugh, S.A.; Turner, J.A.; Burrill, E.A.; David, W. The Forest Inventory and Analysis Database: Population Estimation User Guide. Available online: https://www.fia.fs.fed.us/library/database-documentation/current/ver80/FIADB%20Population%20Estimation%20user%20guide_11_2018_final_revised_02_2019.pdf (accessed on 16 November 2021).
- Hoover, C.M.; Smith, J.E. Selecting Minimum Diameter for Forest Biomass and Carbon Estimation: How Low Should You Go? United States Department of Agriculture, Forest Service, Northern Research Station: Madison, WI, USA, 2020. [CrossRef]
- Janowiak, M.K.; Nagel, L.M.; Webster, C.R. Spatial scale and stand structure in northern hardwood forests: Implications for quantifying diameter distributions. For. Sci. 2008, 54, 497–506. [Google Scholar] [CrossRef]
- Rubin, B.D.; Manion, P.D.; Faber-Langendoen, D. Diameter distributions and structural sustainability in forests. For. Ecol. Manag. 2006, 222, 427–438. [Google Scholar] [CrossRef]
- ESRI. ArcMap, 10.7.1; Environmental Systems Research Institute Inc.: Redlands, CA, USA, 2019.
- Alessandrini, A.; Biondi, F.; Di Filippo, A.; Ziaco, E.; Piovesan, G. Tree size distribution at increasing spatial scales converges to the rotated sigmoid curve in two old-growth beech stands of the Italian Apennines. For. Ecol. Manag. 2011, 262, 1950–1962. [Google Scholar] [CrossRef]
- Halpin, C.R.; Lorimer, C.G.; Hanson, J.J.; Palik, B.J. Predicted long-term effects of group selection on species composition and stand structure in northern hardwood forests. For. Ecol. Manag. 2017, 400, 677–691. [Google Scholar] [CrossRef]
- Riitters, K.; Robertson, G. The United States’ Implementation of the Montreal Process Indicator of Forest Fragmentation. Forests 2021, 12, 727. [Google Scholar] [CrossRef]
- Ostrom, E. A General Framework for Analyzing Sustainability of Social-Ecological Systems. Science 2009, 325, 419–422. [Google Scholar] [CrossRef]
- Hastings, J.M.; Potter, K.M.; Koch, F.H.; Megalos, M.; Jetton, R.M. Prioritizing conservation seed banking locations for imperiled hemlock species using multi-attribute frontier mapping. New For. 2017, 48, 301–316. [Google Scholar] [CrossRef]
- Muller, R.N. Nutrient relations of the herbaceous layer in deciduous forest ecosystems. In The Herbaceous layer in Forests of Eastern North America; Gilliam, F.S., Roberts, M.R., Eds.; Oxford University Press: New York, NY, USA, 2003; pp. 15–37. [Google Scholar]
- Anderson, S.M.; Heath, L.S.; Emery, M.R.; Hicke, J.A.; Littell, J.S.; Lucier, A.; Masek, J.G.; Peterson, D.L.; Pouyat, R.; Potter, K.M.; et al. Developing a set of indicators to identify, monitor, and track impacts and change in forests of the United States. Clim. Chang. 2021, 165, 16. [Google Scholar] [CrossRef]
- Sauer, J.R.; Peterjohn, B.G.; Link, W.A. Observer differences in the North American Breeding Bird Survey. Auk 1994, 111, 50–62. [Google Scholar] [CrossRef]
- Keller, C.M.E.; Scallan, J.T. Potential roadside biases due to habitat changes along breeding bird survey routes. Condor 1999, 101, 50–57. [Google Scholar] [CrossRef]
- Hoban, S.; Bruford, M.; Jackson, J.D.; Lopes-Fernandes, M.; Heuertz, M.; Hohenlohe, P.A.; Paz-Vinas, I.; Sjogren-Gulve, P.; Segelbacher, G.; Vernesi, C.; et al. Genetic diversity targets and indicators in the CBD post-2020 Global Biodiversity Framework must be improved. Biol. Conserv. 2020, 248, 11. [Google Scholar] [CrossRef]
- Laikre, L.; Hoban, S.; Bruford, M.W.; Segelbacher, G.; Allendorf, F.W.; Gajardo, G.; Rodriguez, A.G.; Hedrick, P.W.; Heuertz, M.; Hohenlohe, P.A.; et al. Post-2020 goals overlook genetic diversity. Science 2020, 367, 1083–1085. [Google Scholar] [CrossRef] [Green Version]

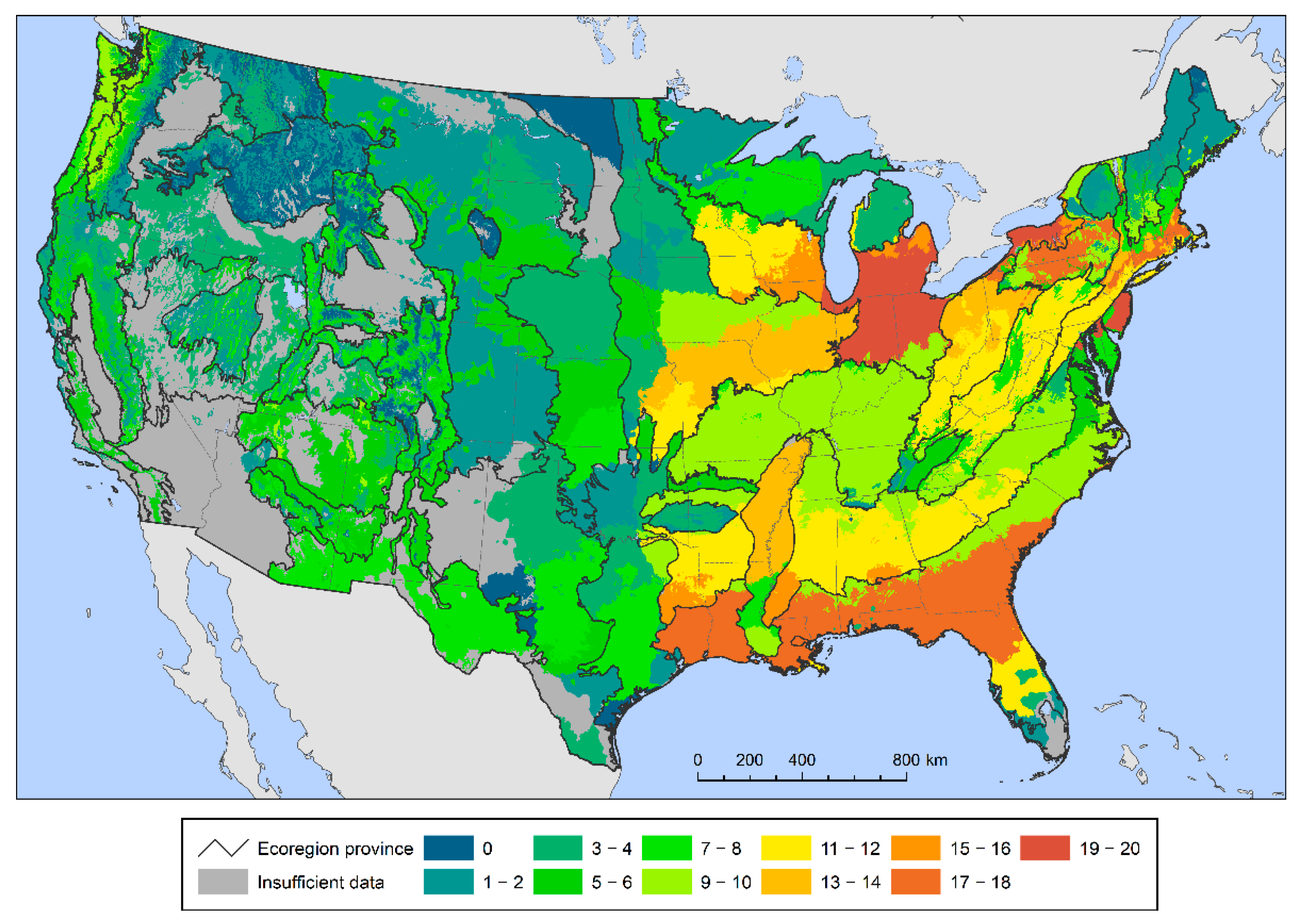


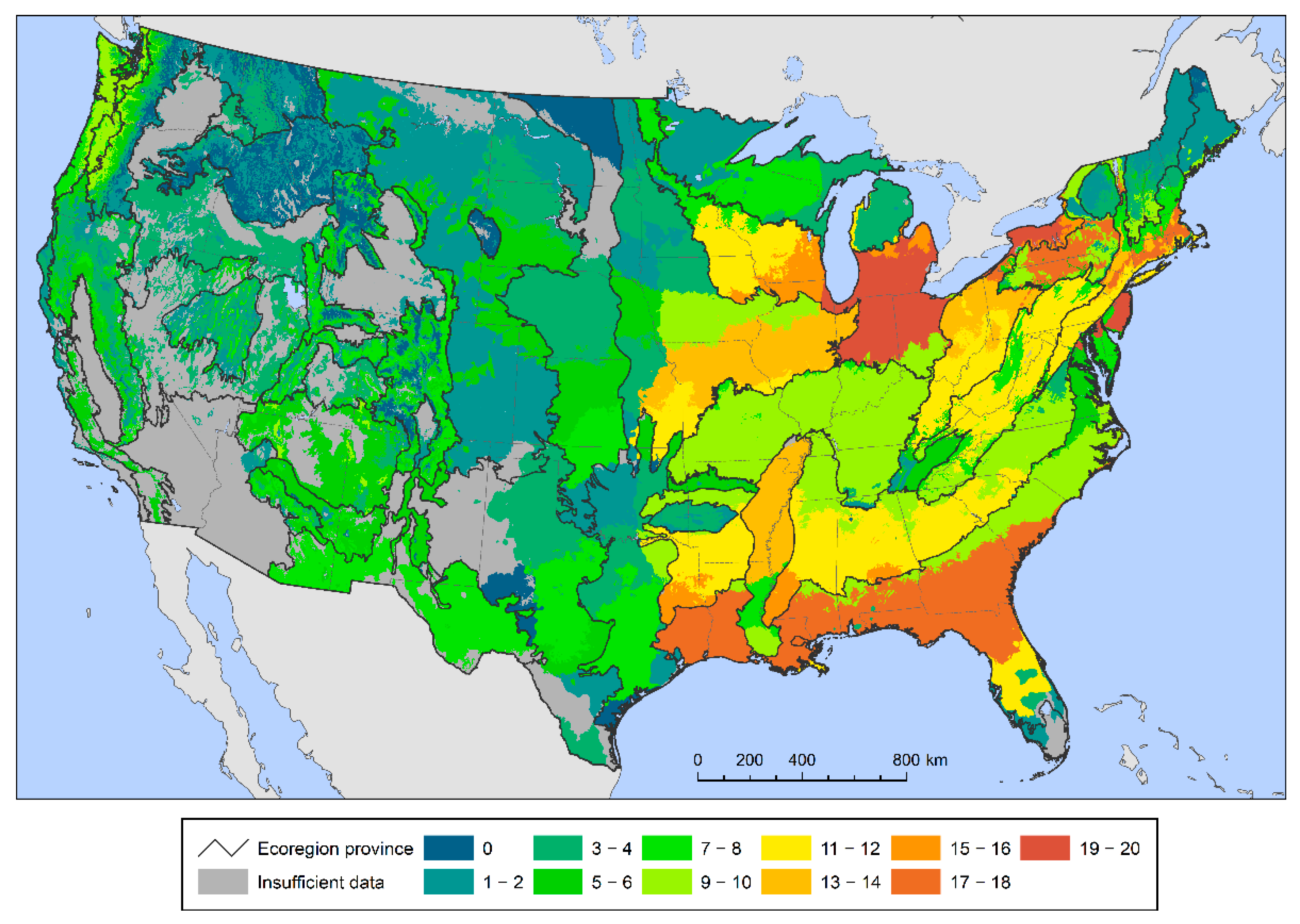
{kind=link}
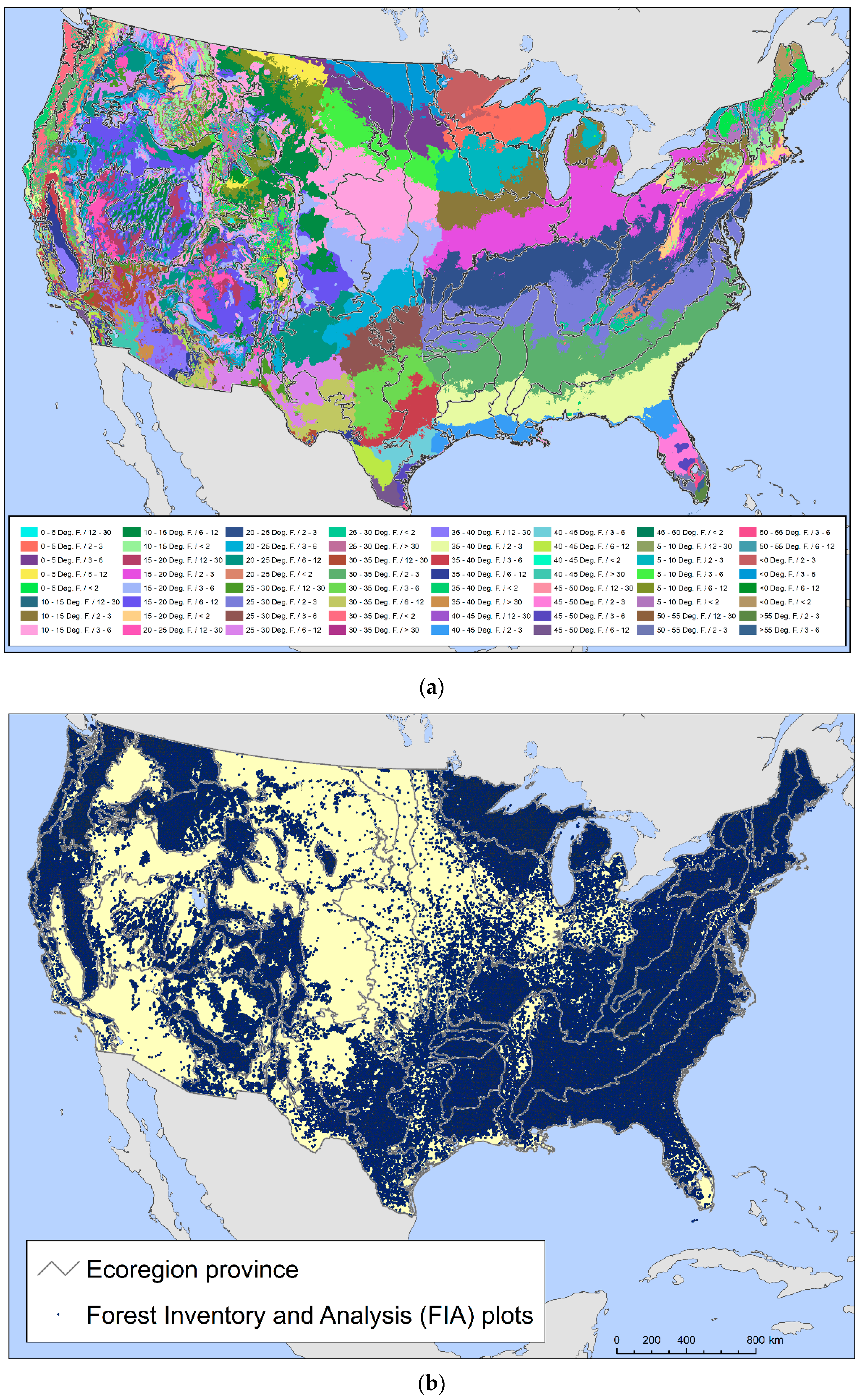
{kind=link}
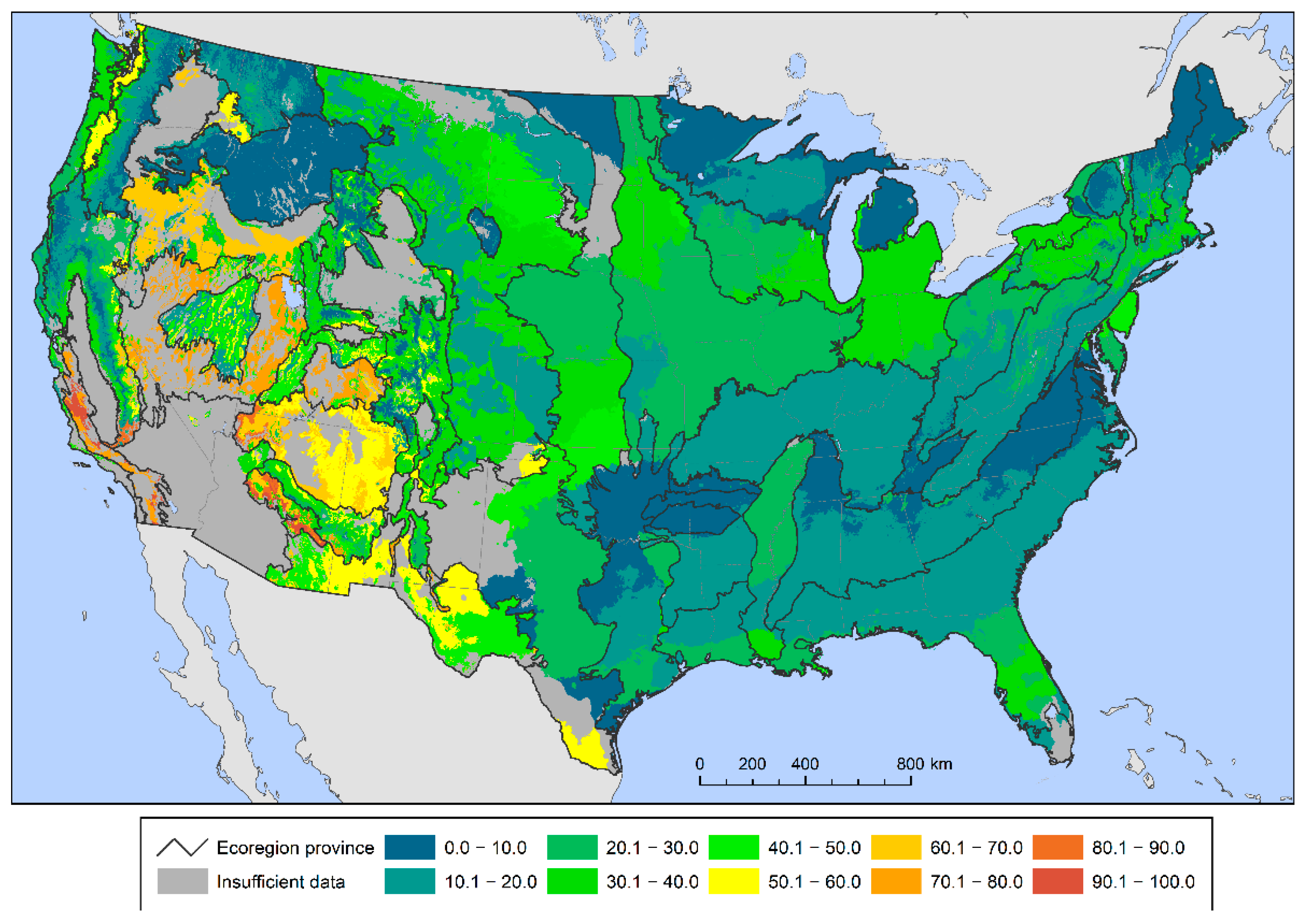{kind=link}
| Criterion 1: Conservation of Biological Diversity. |
|---|
| 1.1 Ecosystem diversity |
| 1.1.a Area and percent of forest by forest ecosystem type, successional stage, age class, and forest ownership or tenure |
| 1.1.b Area and percent of forest in protected areas by forest ecosystem type, and by age class or successional stage |
| 1.1.c Fragmentation of forests |
| 1.2 Species diversity |
| 1.2.a Number of native forest associated species |
| 1.2.b Number and status of native forest associated species at risk, as determined by legislation or scientific assessment |
| 1.2.c Status of on-site and off-site efforts focused on conservation of species diversity |
| 1.3 Genetic diversity |
| 1.3.a Number and geographic distribution of forest associated species at risk of losing genetic variation and locally adapted genotypes |
| 1.3.b Population levels of selected representative forest associated species to describe genetic diversity |
| 1.3.c Status of on-site and off-site efforts focused on conservation of genetic diversity |
| Estimated Stems | |||||
|---|---|---|---|---|---|
| Species | Plots | Small | Large | % Small | Region |
| Pinus muricata | 11 | . | 3,201,331 | 0.0 | CA |
| Platanus racemosa | 12 | . | 568,110 | 0.0 | CA |
| Pseudotsuga macrocarpa | 14 | . | 876,130 | 0.0 | CA |
| Sequoiadendron giganteum | 6 | . | 642,261 | 0.0 | CA |
| Quercus laceyi | 42 | 3,531,744 | 13,855,531 | 20.3 | TX |
| Pinus radiata | 7 | 805,720 | 2,256,087 | 26.3 | CA |
| Ebenopsis ebano | 7 | 3,143,280 | 7,776,171 | 28.8 | TX |
| Pinus longaeva | 33 | 4,036,177 | 8,117,168 | 33.2 | CA/RM |
| Quercus lobata | 96 | 6,738,616 | 11,186,381 | 37.6 | CA |
| Ulmus serotina | 7 | 1,323,092 | 2,164,378 | 37.9 | SE |
| Bursera simaruba | 5 | 453,118 | 714,342 | 38.8 | SE |
| Quercus engelmannii | 8 | 501,619 | 774,952 | 39.3 | CA |
| Quercus oblongifolia | 32 | 2,945,510 | 4,439,051 | 39.9 | SW |
| Pinus balfouriana | 23 | 3,667,384 | 4,781,293 | 43.4 | CA |
| Juniperus osteosperma | 5192 | 1,705,042,053 | 1,852,085,663 | 47.9 | SW |
| Juniperus californica | 93 | 20,479,125 | 18,888,399 | 52.0 | CA |
| Pinus leiophylla | 13 | 2,291,034 | 2,086,389 | 52.3 | SW |
| Pinus resinosa | 2358 | 651,585,247 | 592,268,113 | 52.4 | NE/MW |
| Juniperus monosperma | 2381 | 915,076,545 | 784,121,991 | 53.9 | SW |
| Nyssa aquatica | 543 | 274,436,885 | 212,879,335 | 56.3 | SE |
| Morus microphylla | 5 | 909,545 | 680,855 | 57.2 | TX/SW |
| Quercus douglasii | 473 | 229,864,057 | 154,538,031 | 59.8 | CA |
| Pinus pungens | 196 | 33,893,343 | 21,511,606 | 61.2 | SE |
| Pinus rigida | 917 | 333,647,542 | 188,912,475 | 63.8 | NE/SE |
| Pinus jeffreyi | 785 | 303,719,657 | 162,367,673 | 65.2 | CA |
| Platanus wrightii | 5 | 846,542 | 432,163 | 66.2 | SW |
| Condalia hookeri | 278 | 154,649,389 | 75,831,092 | 67.1 | TX |
| Pinus elliottii | 3376 | 3,803,976,324 | 1,784,082,874 | 68.1 | SE |
| Nyssa ogeche | 55 | 19,908,868 | 9,206,676 | 68.4 | SE |
| Juniperus deppeana | 1181 | 478,038,092 | 218,766,045 | 68.6 | SW |
| Carya ovalis | 68 | 8,514,642 | 3,815,860 | 69.1 | NE/SE/MW |
| Tilia americana heterophylla | 37 | 7,200,971 | 3,074,222 | 70.1 | SE/MW/NE |
| Alnus rubra | 1991 | 956,957,735 | 393,555,656 | 70.9 | NW/CA |
| Salix bebbiana | 51 | 31,448,863 | 12,327,732 | 71.8 | RM/MW/NE |
| Betula populifolia | 751 | 989,927,141 | 379,812,097 | 72.3 | NE |
| Halesia diptera | 25 | 21,505,462 | 8,121,802 | 72.6 | SE |
| Pinus serotina | 386 | 204,557,934 | 75,901,847 | 72.9 | SE |
| Maclura pomifera | 1283 | 500,698,545 | 185,376,403 | 73.0 | TX/SE/MW |
| Pinus palustris | 2019 | 1,343,159,776 | 495,078,052 | 73.1 | SE |
| Pinus remota | 32 | 31,546,173 | 11,605,019 | 73.1 | TX |
| Taxodium distichum | 1227 | 698,154,187 | 247,087,517 | 73.9 | SE/TX |
| Malus coronaria | 55 | 48,212,777 | 16,987,961 | 73.9 | MW/NE/SE |
| Pinus aristata | 140 | 80,046,751 | 27,964,956 | 74.1 | RM |
| Prosopis velutina | 362 | 243,842,284 | 84,664,761 | 74.2 | SW |
| Cercocarpus ledifolius | 912 | 752,320,781 | 260,407,121 | 74.3 | RM/NW/CA |
| Populus fremontii | 40 | 19,766,406 | 6,832,552 | 74.3 | CA/SW/TX |
| Seed Zones | |||||
|---|---|---|---|---|---|
| Species | Plots | n | at Risk | % at Risk | Region |
| Juniperus californica | 93 | 6 | 6 | 100.0 | CA |
| Juniperus osteosperma | 5192 | 56 | 56 | 100.0 | SW |
| Pinus pungens | 196 | 5 | 5 | 100.0 | SE |
| Quercus lobata | 96 | 5 | 5 | 100.0 | CA |
| Pinus resinosa | 2358 | 30 | 28 | 93.3 | NE/MW |
| Juniperus monosperma | 2381 | 40 | 37 | 92.5 | SW |
| Quercus douglasii | 473 | 13 | 12 | 92.3 | CA |
| Nyssa aquatica | 543 | 10 | 9 | 90.0 | SE |
| Condalia hookeri | 278 | 7 | 6 | 85.7 | TX |
| Juniperus deppeana | 1181 | 19 | 16 | 84.2 | SW |
| Pinus jeffreyi | 785 | 21 | 17 | 81.0 | CA |
| Prosopis velutina | 362 | 12 | 9 | 75.0 | SW |
| Taxodium distichum | 1227 | 15 | 11 | 73.3 | SE/TX |
| Pinus elliottii | 3376 | 11 | 8 | 72.7 | SE |
| Pinus aristata | 140 | 7 | 5 | 71.4 | RM |
| Pinus palustris | 2019 | 10 | 7 | 70.0 | SE |
| Pinus echinata | 5143 | 31 | 21 | 67.7 | SE |
| Populus deltoides | 994 | 40 | 27 | 67.5 | MW/NE/SE/TX |
| Alnus rubra | 1991 | 21 | 14 | 66.7 | CA/NW |
| Pinus banksiana | 1339 | 9 | 6 | 66.7 | MW/NE |
| Pinus cembroides | 71 | 6 | 4 | 66.7 | TX |
| Betula populifolia | 751 | 20 | 13 | 65.0 | NE |
| Pinus rigida | 917 | 17 | 11 | 64.7 | NE/SE |
| Juniperus coahuilensis | 358 | 14 | 9 | 64.3 | SW/TX |
| Carya laciniosa | 267 | 8 | 5 | 62.5 | MW/SE |
| Maclura pomifera | 1283 | 26 | 16 | 61.5 | TX/SE/MW |
| Alnus rhombifolia | 100 | 7 | 4 | 57.1 | CA/NW |
| Pinus sabiniana | 293 | 14 | 8 | 57.1 | CA |
| Juniperus occidentalis | 1579 | 29 | 16 | 55.2 | CA/NW |
| Planera aquatica | 272 | 11 | 6 | 54.5 | SE |
| Platanus occidentalis | 2630 | 34 | 18 | 52.9 | MW/NE/SE/TX |
| Quercus agrifolia | 287 | 17 | 9 | 52.9 | CA |
| Cercocarpus ledifolius | 912 | 29 | 15 | 51.7 | RM/NW/CA |
| Pinus ponderosa | 11,350 | 115 | 59 | 51.3 | CA/NW/SW/RM |
| Fraxinus caroliniana | 121 | 6 | 3 | 50.0 | SE |
| Magnolia macrophylla | 251 | 10 | 5 | 50.0 | SE |
| Pinus taeda | 18,674 | 38 | 19 | 50.0 | SE |
| Sequoia sempervirens | 304 | 8 | 4 | 50.0 | CA |
| Taxodium ascendens | 863 | 8 | 4 | 50.0 | SE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potter, K.M.; Riitters, K. A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss. Forests 2022, 13, 19. https://doi.org/10.3390/f13010019
Potter KM, Riitters K. A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss. Forests. 2022; 13(1):19. https://doi.org/10.3390/f13010019
Chicago/Turabian StylePotter, Kevin M., and Kurt Riitters. 2022. "A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss" Forests 13, no. 1: 19. https://doi.org/10.3390/f13010019
APA StylePotter, K. M., & Riitters, K. (2022). A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss. Forests, 13(1), 19. https://doi.org/10.3390/f13010019