
Mid-Rotation Impacts of Stand Tending with Glyphosate on Plant Diversity in the Boreal Forest of West-Central Alberta


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
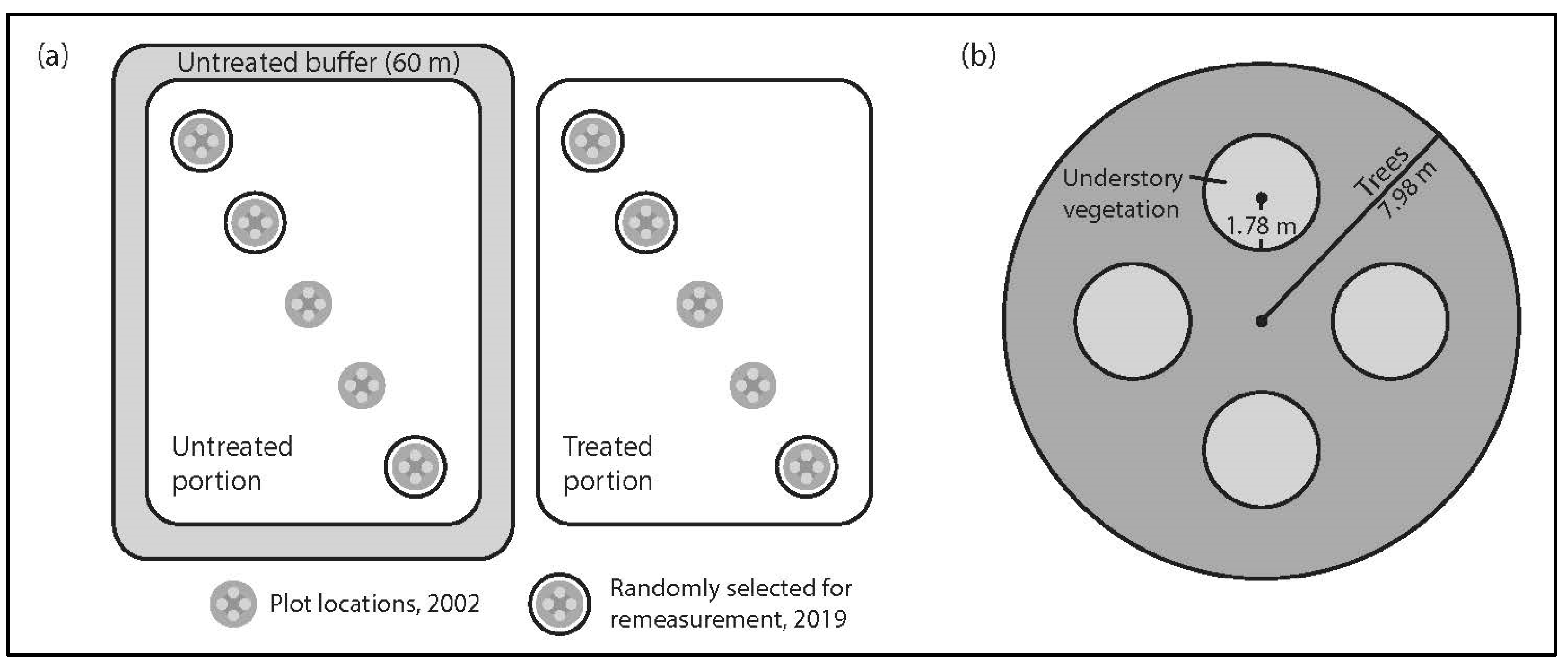
2.2. Experimental Design
- Installations were placed in cutblocks that were uniform, including
- Uniformity in the site across treated and untreated areas;
- Uniformity in harvesting and silvicultural treatment across treated and untreated areas, with the exception of herbicide application;
- The untreated portion was at least 1 ha in size;
- The untreated portion was buffered from herbicide treatment with buffer width dependent on the method of herbicide application;
- Buffer areas were not included in the measurement portion of the untreated reference area.
2.3. Field Sampling
2.4. Statistical Analysis
2.4.1. Overstory Vascular Plant Diversity and Dominance
2.4.2. Understory Plant Diversity
3. Results
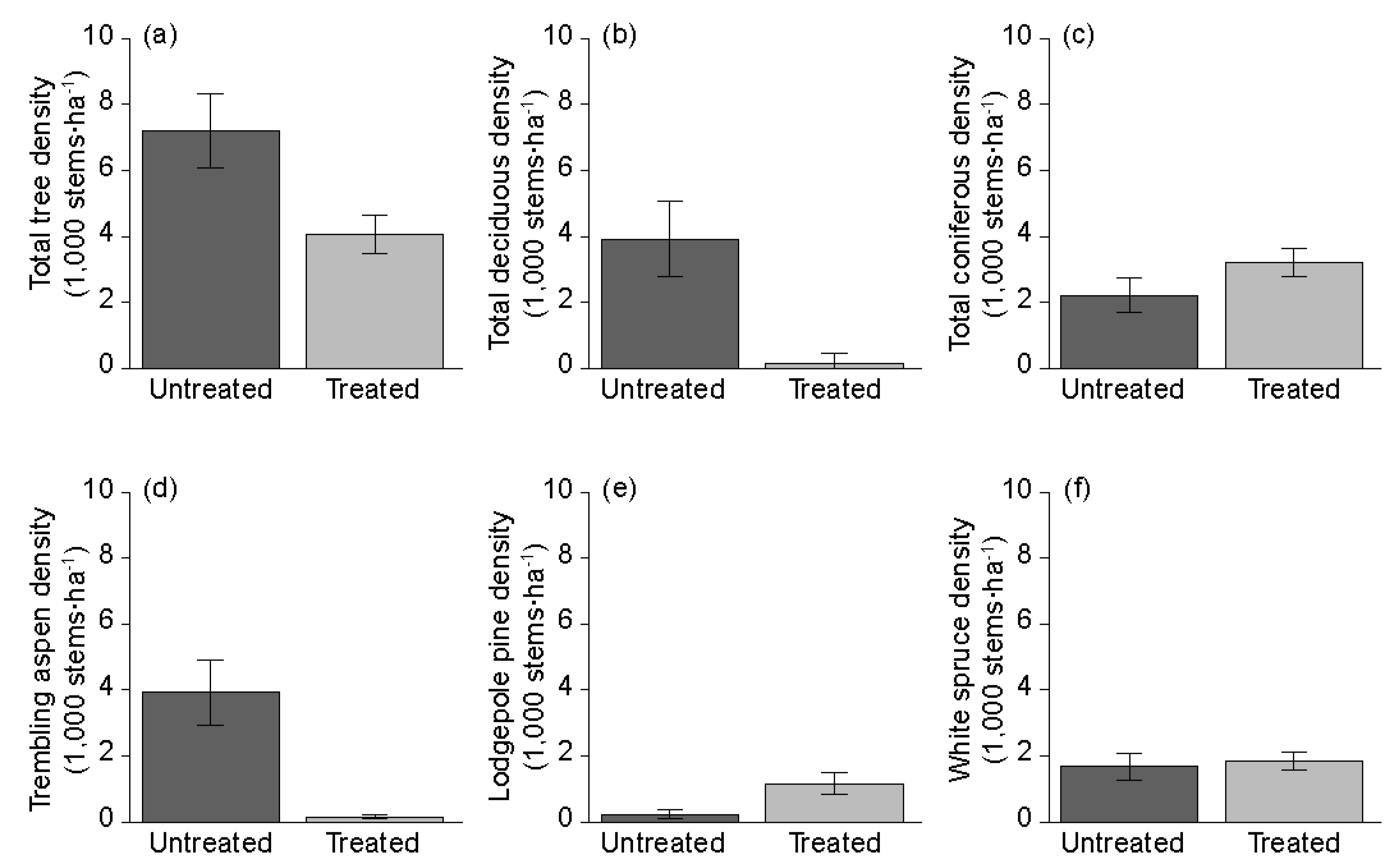
3.1. Overstory Plant Diversity and Dominance
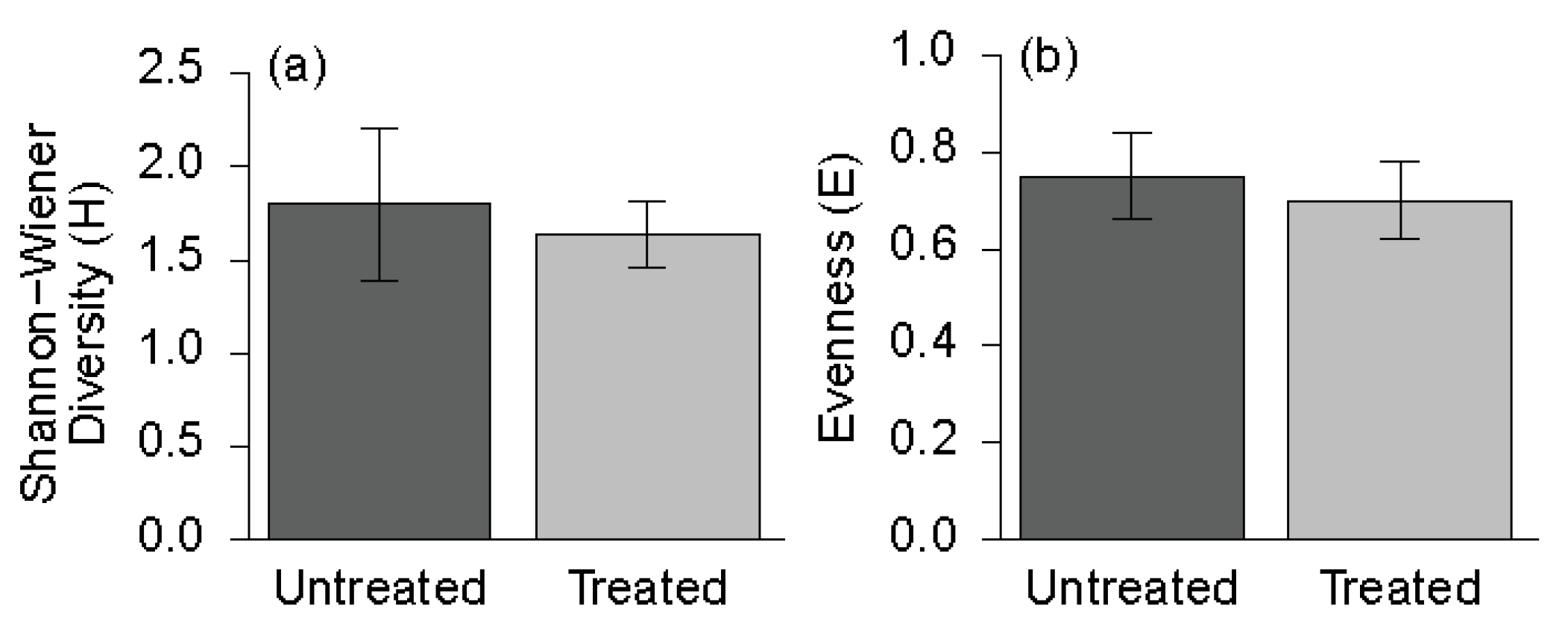
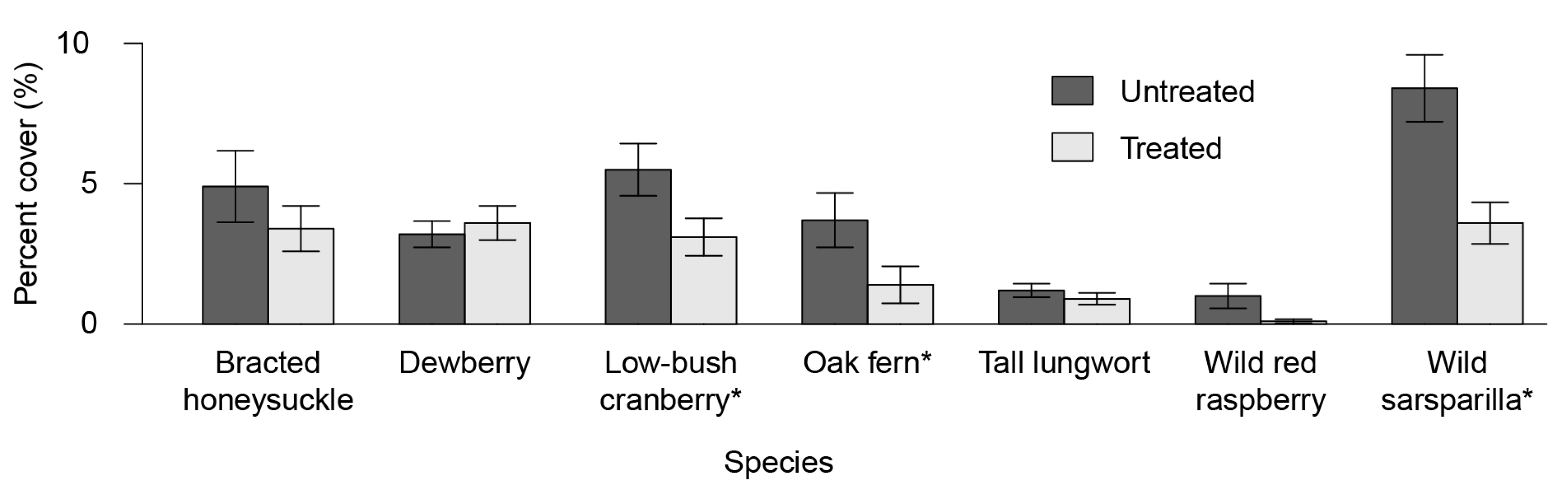
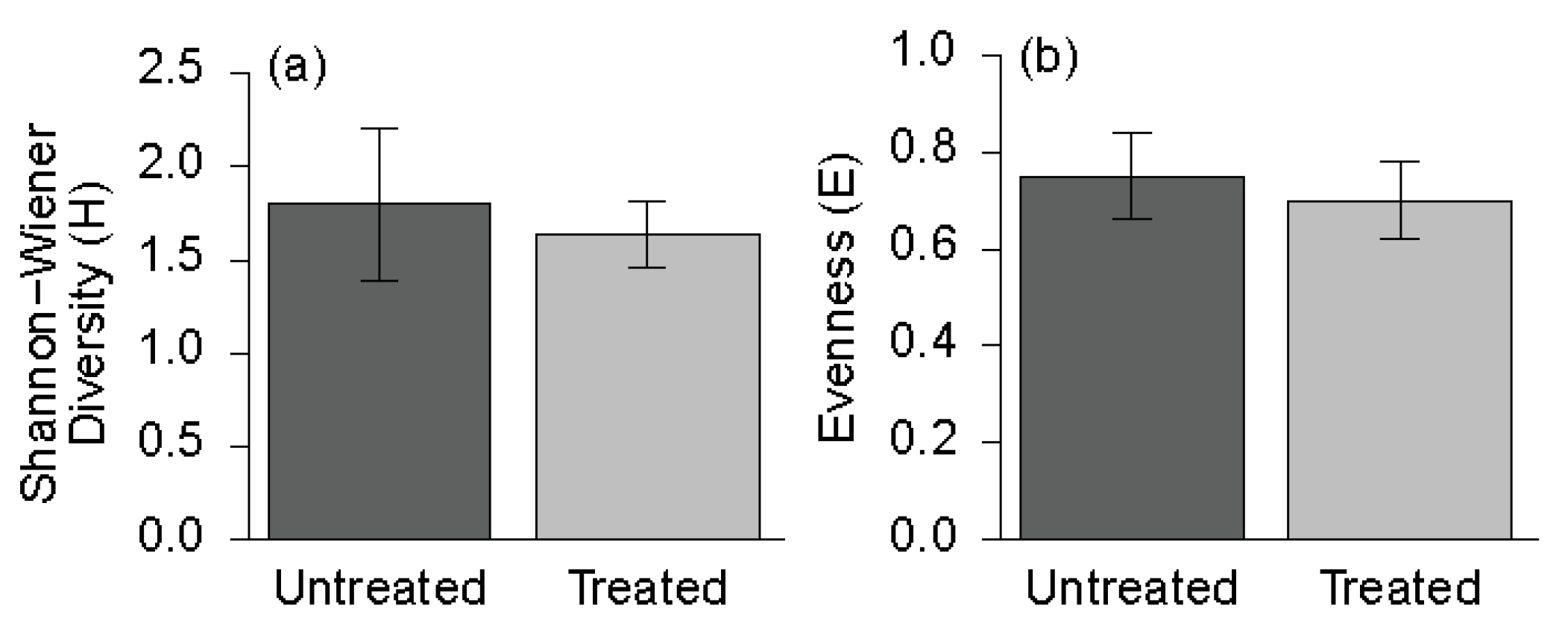
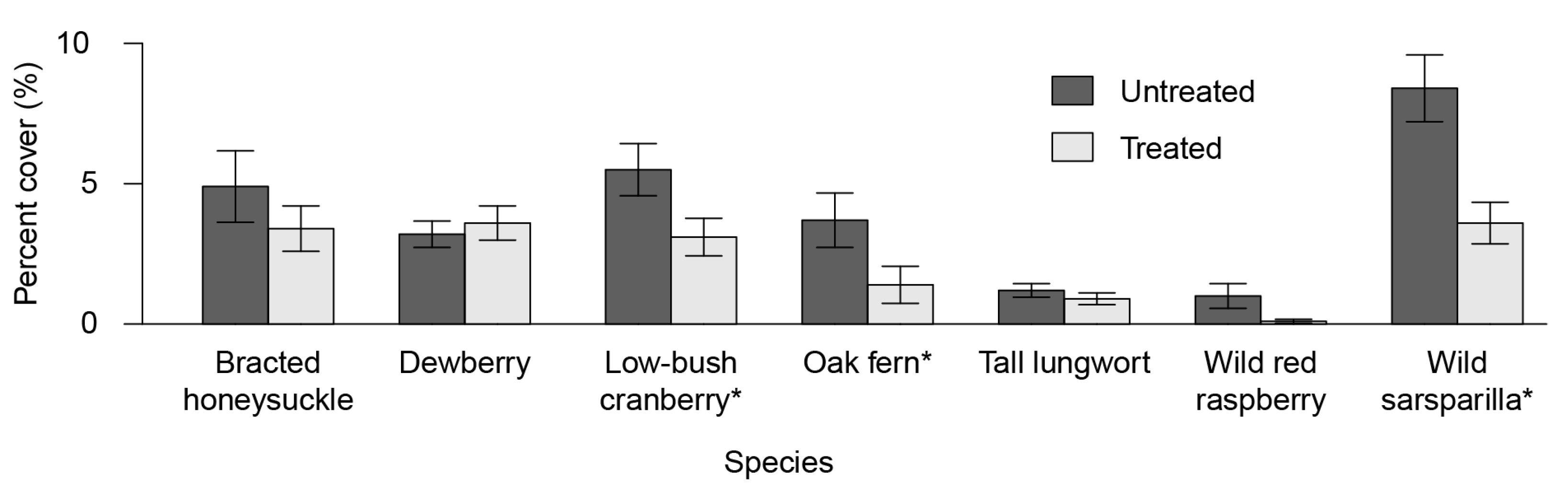
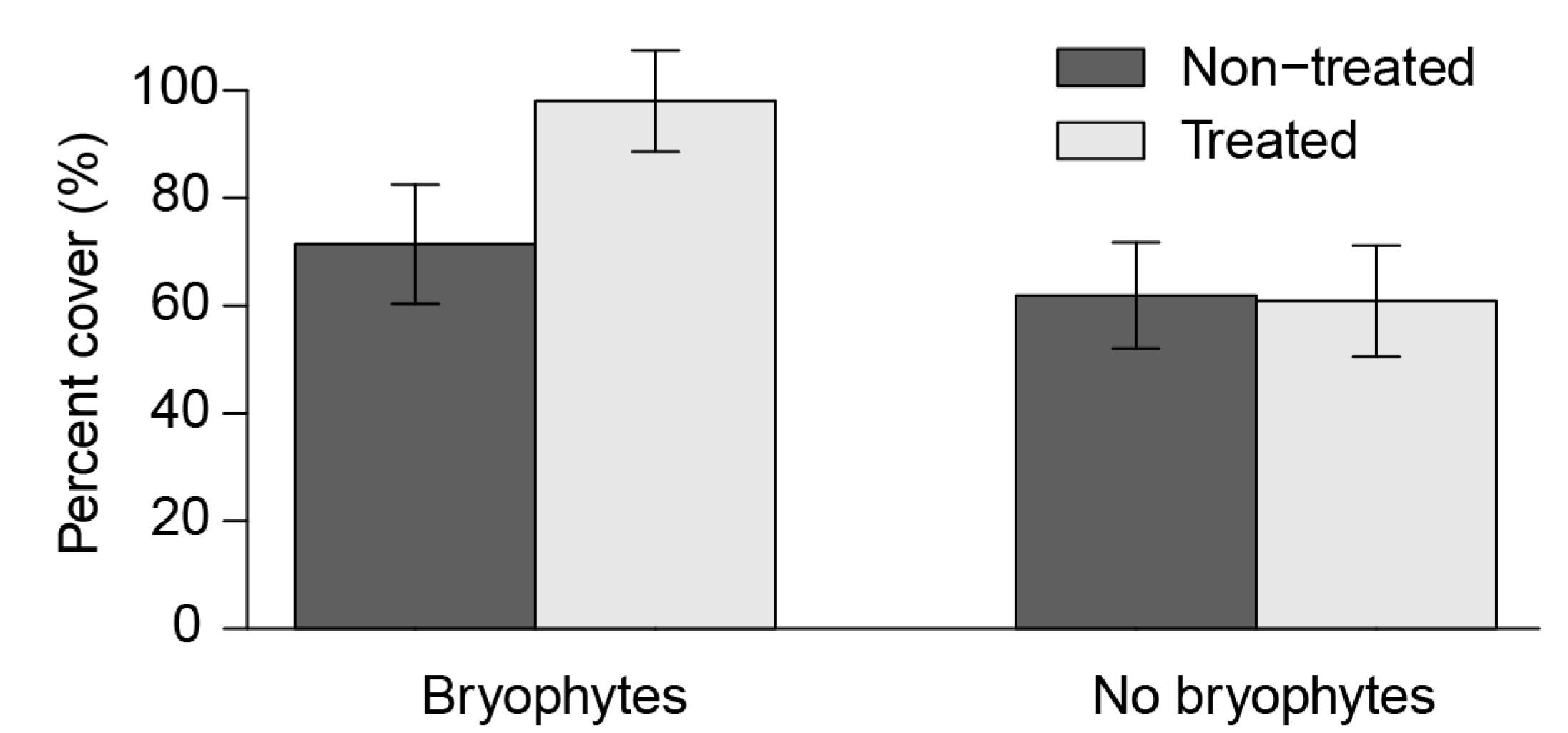
3.2. Understory Plant Diversity
4. Discussion
4.1. Overstory Vascular Plants
4.2. Understory Vegetation
4.3. Study Limitations
5. Conclusions
- Establish a series of glyphosate demonstration sites that contrast untreated with treated areas and establish and measure ongoing monitoring plots;
- Identify both chemical herbicide and other non-herbicide alternatives to glyphosate and establish trials to compare these options to glyphosate;
- Engage with key stakeholders, especially Indigenous stakeholders and/or collaborators, when selecting treatments and designing the installations suggested in items 1 and 2. In particular, seek Indigenous engagement (and potentially study partners) when assessing herbicide impact on shrub and herbaceous plants as many of these species are culturally important. Understory plant presence/abundance data collected in this evaluation might form the basis of an assessment of impact on species of Indigenous importance.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | English Name |
|---|---|
| Achillea millefolium | Common yarrow |
| Actaea rubra | Red and white baneberry |
| Alnus crispa | Green alder |
| Alnus tenuifolia | River alder |
| Aquilegia brevistyla | Blue columbine |
| Aralia nudicaulis | Wild sarsaparilla |
| Aster conspicuus | Showy aster |
| Astragalus americanus | American milk-vetch |
| Athyrium filix-femina | Lady fern |
| Calamagrostis canadensis | Marsh reed grass |
| Circaea alpina | Small enchanters-nightshade |
| Cornus canadensis | Bunchberry |
| Disporum trachycarpum | Fairybells |
| Dryopteris austriaca | Shield fern |
| Elymus innovatus | Hairy wild rye |
| Epilobium angustitolium | Fireweed |
| Equisetum arvense | Common horsetail |
| Equisetum sylvaticum | Woodland horsetail |
| Fragaria vesca | Woodland strawberry |
| Fragaria virginiana | Wild strawberry |
| Galium boreale | Northern bedstraw |
| Galium triflorum | Sweet-scented bedstraw |
| Geum aleppicum | Yellow avens |
| Geum rivale | Purple avens |
| Gymnocarpium dryopteris | Oak fern |
| Heracleum lanatum | Cow-parsnip |
| Lathyrus ochroleucus | Creamy peavine |
| Ledum groenlandicum | Common labrador tea |
| Linnaea borealis | Twinflower |
| Lonicera dioica | Twining honeysuckle |
| Lonicera involucrata | Bracted honeysuckle |
| Lycopodium annotinum | Stiff club-moss |
| Maianthemum canadense | Wild lily-of-the-valley |
| Mertensia paniculata | Tall lungwort |
| Mitella nuda | Common mitrewort |
| Oryszopsis asperifolia | Rough-leaved rice grass |
| Petasites palmatus | Palmate-leaved coltsfoot |
| Petasites sagittatus | Arrow-leaved coltsfoot |
| Potentilla fruticosa | Shrubby cinquefoil |
| Pyrola asarifolia | Common pink wintergreen |
| Pyrola chlorantha | Green wintergreen |
| Pyrola secunda | One-sided wintergreen |
| Ribes glandulosum | Skunk currant |
| Ribes hudsonianum | Northern black currant |
| Ribes lacustre | Black gooseberry |
| Ribes oxyacanthoides | Northern gooseberry |
| Ribes triste | Wild red currant |
| Rosa acicularis | Prickly rose |
| Rubus idaeus | Wild red raspberry |
| Rubus parviflorus | Thimbleberry |
| Rubus pedatus | Dwarf bramble |
| Rubus pubescens | Dewberry |
| Salix bebbiana | Beaked willow |
| Salix spp. | Unknown willow species |
| Shepherdia canadensis | Canada buffaloberry |
| Smilacina stellata | Star-flowered false Solomons-seal |
| Sorbus scopulina | Western mountain ash |
| Spiraea betulifolia | White meadowsweet |
| Streptopus amplexifolius | Twisted-stalk |
| Symphoricarpos albus | Common snowberry |
| Taraxacum officinale | Common dandelion |
| Trifolium pratense | Red clover |
| Vaccinium myrtilloides | Common blueberry |
| Vaccinium vitis-idaea | Bog cranberry |
| Viburnum edule | Low bush-cranberry |
| Viburnum opulus | High bush-cranberry |
| Vicia americana | Wild vetch |
| Viola canadensis | Western Canada violet |
| Viola renifolia | Kidney-leaved violet |
| Feather moss | Feather moss |
| Wood moss | Wood moss |
| Leaf lichen | Leaf lichen |
| Unknown grass species | Unknown grass species |
References
- Wagner, R.G.; Little, K.M.; Richardson, B.; McNabb, K. The role of vegetation management in enhancing productivity of the world’s forests. Forestry 2006, 79, 57–79. [Google Scholar] [CrossRef]
- Boateng, J.O.; Heineman, J.L.; McLarnon, J.; Bedford, L. Twenty year response of white spruce to mechanical site preparation and early chemical release in the boreal region of northeastern British Columbia. Can. J. For. Res. 2006, 36, 2386–2399. [Google Scholar] [CrossRef]
- Cole, E.C.; Newton, M.; Youngblood, A. Regenerating white spruce, paper birch and willow in central Alaska. Can. J. For. Res. 1999, 29, 993–1001. [Google Scholar] [CrossRef]
- CCFM: National Forest Database. Canadian Council of Forest Ministers. 2022. Available online: http://nfdp.ccfm.org/en/data/harvest.php (accessed on 13 July 2022).
- Wagner, R.G.; Flynn, J.; Gregory, R. Public perceptions of risk and acceptability of forest vegetation management alternatives in Ontario. For. Chron. 1998, 74, 720–727. [Google Scholar] [CrossRef]
- Buse, L.J.; Wagner, R.G.; Perrin, B. Public attitudes to forest herbicide use and the implications for public involvement. For. Chron. 1995, 71, 596–600. [Google Scholar] [CrossRef]
- Mihajlovich, M.; Odsen, S.; Chicoine, D. Review of herbicide use for forest management in Alberta, 1995–2009. For. Chron. 2012, 88, 328–339. [Google Scholar] [CrossRef]
- Lautenschlager, R.A.; Sullivan, T.P. Effects of herbicide treatments on biotic components in regenerating northern forests. For. Chron. 2002, 78, 695–731. [Google Scholar] [CrossRef]
- Comeau, P.G.; Filipescu, C.N.; Kabzems, R.; DeLong, C. Early growth of white spruce underplanted beneath spaced and unspaced aspen stands in northeastern British Columbia. Can. J. For. Res. 2004, 34, 2277–2283. [Google Scholar] [CrossRef]
- Kabzems, R.D.; Harper, G.; Fielder, P. Growing space management in boreal mixedwood forests. West. J. Appl. For. 2011, 26, 82–90. [Google Scholar] [CrossRef]
- Kayahara, G.J.; Chen, H.Y.H.; Klinka, K.; Coates, K.D. Relations of Terminal Growth and Specific Leaf Area to Available Light in Naturally Regenerated Seedlings of Lodgepole Pine and Interior Spruce in Central British Columbia; Research Report No. 9; Research Branch, B.C. Ministry of Forests, 31 Bastion Square: Victoria, BC, Canada. 1995. Available online: https://www.for.gov.bc.ca/hfd/pubs/docs/rr/rr09.pdf (accessed on 13 July 2022).
- Bell, F.W.; Ter-Mikaelian, M.T.; Wagner, R.G. Relative competitiveness of nine early-successional boreal forest species associated with planted jack pine and black spruce seedlings. Can. J. For. Res. 2000, 30, 790–800. [Google Scholar] [CrossRef]
- Olson, M.G.; Wagner, R.G.; Brissette, J.C. Forty years of spruce-fir stand development following herbicide application and precommercial thinning in central Maine, USA. Can. J. For. Res. 2012, 41, 1–11. [Google Scholar] [CrossRef]
- Comeau, P.G.; Fraser, E.C. Plant community diversity and tree growth following single and repeated glyphosate herbicide applications to a white spruce plantation. Forests 2018, 9, 107. [Google Scholar] [CrossRef]
- Boateng, J.O.; Haeussler, S.; Bedford, L. Boreal plant community diversity 10 years after glyphosate treatment. West. J. Appl. For. 2000, 15, 15–26. [Google Scholar] [CrossRef]
- Haeussler, S.; Bartemucci, P.; Bedford, L. Succession and resilience in boreal mixedwood plant communities 15–16 years after silvicultural site preparation. For. Ecol. Mgmt. 2004, 199, 349–370. [Google Scholar] [CrossRef]
- Pitt, D.G.; Mihajlovich, M.; Proudfoot, L.M. Juvenile stand responses and potential outcomes of conifer release efforts on Alberta’s spruce–aspen mixedwood sites. For. Chron. 2004, 80, 583–597. [Google Scholar] [CrossRef]
- Natural Regions Committee. Natural Regions and Subregions of Alberta; Downing, D.J., Pettapiece, W.W., Eds.; Government of Alberta: Edmonton, AB, Canada, 2006; Available online: https://www.albertaparks.ca/media/2942026/nrsrcomplete_may_06.pdf (accessed on 13 July 2022).
- Beckingham, J.D.; Corns, I.G.W.; Archibald, J.H. Field Guide to the Ecosites of West-Central Alberta; Special Report 9; Natural Resources Canada, Canadian Forest Service; Northern Forestry Centre: Edmonton, AB, Canada, 1996; 540p. [Google Scholar]
- Staudhammer, C.L.; LeMay, V.M. Introduction and evaluation of possible indices of stand structural diversity. Can. J. For. Res. 2001, 31, 1105–1115. [Google Scholar] [CrossRef]
- SAS. Statistical Analysis Software. Users’ Guide Statistics, version 9.4; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 13 July 2022).
- Broman, K.W. Broman: Karl Broman’s R Code. R Package Version 0.72-4. 2021. Available online: https://CRAN.R-project.org/package=broman (accessed on 13 July 2022).
- Heip, C.H.R.; Herman, P.M.J.; Soetaert, K. Indices of diversity and evenness. Océanis 1998, 24, 61–87. [Google Scholar]
- Whittaker, R.H. Vegetation of the Siskiyou mountains, Oregon and California. Ecol. Monogr. 1960, 30, 280–338. [Google Scholar] [CrossRef]
- Jurasinski, G.; Retzer, V.; Beierkuhnlein, C. Inventory, differentiation, and proportional diversity: A consistent terminology for quantifying species diversity. Oecologia 2009, 159, 15–26. [Google Scholar] [CrossRef]
- Hunter, M., Jr. Fundamentals of Conservation Biology, 2nd ed.; Blackwell Science: Malden, MA, USA, 2002. [Google Scholar]
- Dempster, W.R.; Meredith, S. A discussion of best management practices for reforestation following harvesting of lodgepole pine in the Alberta Foothills. For. Chron. 2014, 90, 763–770. [Google Scholar] [CrossRef]
- Reforestation Standard of Alberta. Government of Alberta. 2021. Available online: https://open.alberta.ca/publications/7010852 (accessed on 13 July 2022).
- Bataineh, M.M.; Wagner, R.G.; Olson, M.G.; Olson, E.K. Mid-rotation response of ground vegetation to herbicide and precommercial thinning in the Acadian Forest of Maine, USA. For. Ecol. Mgmt. 2014, 313, 132–143. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Armstrong, G.W.; Stadt, K.J.; Marenholtz, E.H. Forest regeneration standards: Are they limiting management options for Alberta’s boreal mixedwoods? For Chron. 2008, 84, 76–82. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Pinno, B.D.; Beverly, J.L.; Thomas, B.R.; Nock, C. Reforestation policy has constrained options for managing risks on public forests. Can. J. For. Res. 2020, 50, 855–861. [Google Scholar] [CrossRef]


| 2019 (Site ID) | 2002 (Field ID) | Site Preparation | Propagule Type | Herbicide Applied | Remedial Treatment | Note |
|---|---|---|---|---|---|---|
| 1 | 130-75 | Power disk trench | Plant (Sw/Pl) | 1997 | Patch mound, patch plough, fill plant, glyphosate | The remedial treatments did not impact the untreated plot. |
| 2 | 120-37 | Plough | Plant (Sw) | 1990 | None | Follow up motor manual tending 2002. |
| 5 | WP-1012 | Disk trench | Plant (Sw) | 1994 | None | |
| 6 | 690-38 | Spot mound | Plant (Pl/Sw/Sb) | 1999 | Glyphosate | |
| 7 | HC-1096 | Disk trench | Plant (Sw) | 1996 | None | |
| 8 | W06-1048A | Mound | Plant (Sw/Pl) | 1999 | None | Aerial spray buffers along block boundary were treated with triclopyr in 2006. |
| 9 | W06_1042 | None | Plant (Sw) | 1998 | None | Aerial spray buffers along block boundary were treated with triclopyr in 2006. |
| 10 | S14036 | Power disk trench | Plant (Sw, Pl) | 1999 | Fill plant | |
| NA | W06_1002 | Untreated area was compromised by a pipeline. | ||||
| NA | 270-58 | Untreated area treated with glyphosate post 2003. | ||||
| NA | 152 | Untreated area treated with glyphosate post 2003. | ||||
| NA | 179 | Untreated area treated with glyphosate post 2003. |
| Tree Species/Group | Average Stem Density (Stems·ha−1) | |||
|---|---|---|---|---|
| Untreated | Treated | Relative % Change ((Treated – Untreated)/Untreated × 100) | p-Value | |
| All tree species | 7198 (1121) | 4060 (580) | −43.60 | 0.047 |
| Deciduous trees | 4990 (1161) | 850 (291) | −82.97 | 0.008 |
| Coniferous trees | 2208 (523) | 3210 (445) | 45.38 | 0.234 |
| Trembling aspen | 3927 (983) | 154 (62) | −96.08 | 0.008 |
| Lodgepole pine | 221 (138) | 1163 (333) | 426.24 | 0.008 |
| White spruce | 1688 (405) | 1852 (267) | 9.72 | 0.805 |
| Species | Partial R-Squared | p-Value | No. of Subplots Observed (Out of 192) |
|---|---|---|---|
| Forest Floor Bryophytes | 0.569 | <0.00001 | 124 (65%) |
| Dewberry (Rubus pubescens) | 0.122 | 0.016 | 116 (60%) |
| Blue columbine (Aquilegia brevistyla) | 0.090 | 0.043 | 1 (0.5%) |
| Shrubby cinquefoil (Potentilla fruticose) | 0.095 | 0.040 | 1 (0.5%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihajlovich, M.; Nash, J.C.; Odsen, S.; Roth, B.E. Mid-Rotation Impacts of Stand Tending with Glyphosate on Plant Diversity in the Boreal Forest of West-Central Alberta. Forests 2022, 13, 1585. https://doi.org/10.3390/f13101585
Mihajlovich M, Nash JC, Odsen S, Roth BE. Mid-Rotation Impacts of Stand Tending with Glyphosate on Plant Diversity in the Boreal Forest of West-Central Alberta. Forests. 2022; 13(10):1585. https://doi.org/10.3390/f13101585
Chicago/Turabian StyleMihajlovich, Milo, John C. Nash, Sonya Odsen, and Brian E. Roth. 2022. "Mid-Rotation Impacts of Stand Tending with Glyphosate on Plant Diversity in the Boreal Forest of West-Central Alberta" Forests 13, no. 10: 1585. https://doi.org/10.3390/f13101585
APA StyleMihajlovich, M., Nash, J. C., Odsen, S., & Roth, B. E. (2022). Mid-Rotation Impacts of Stand Tending with Glyphosate on Plant Diversity in the Boreal Forest of West-Central Alberta. Forests, 13(10), 1585. https://doi.org/10.3390/f13101585