Dispersal Patterns of One-Seed Juniper Seeds Contained in Mammal Scats and Bird Pellets

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Species
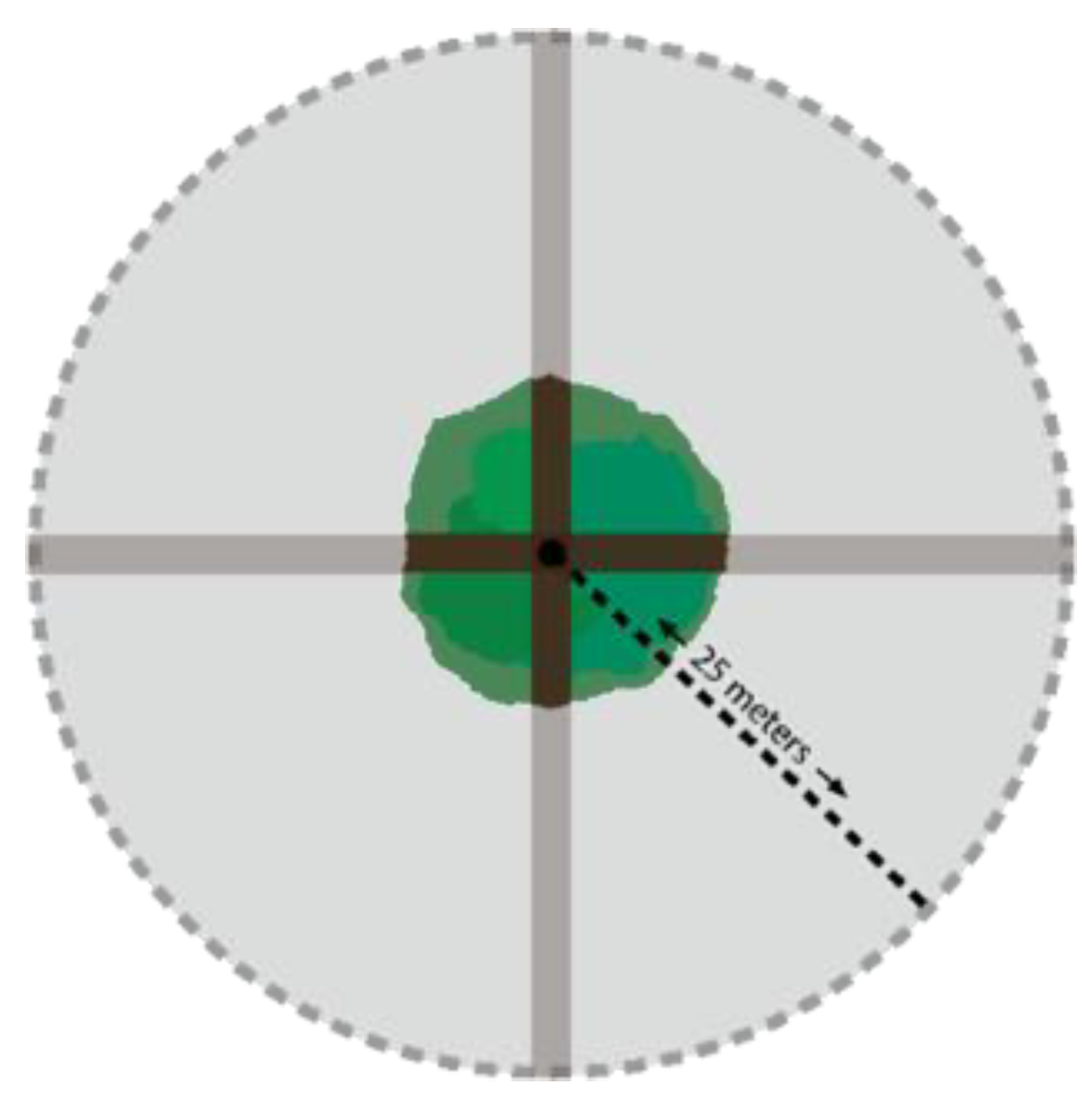
2.3. Study Design
2.4. Seed Counts
2.5. Statistical Analyses
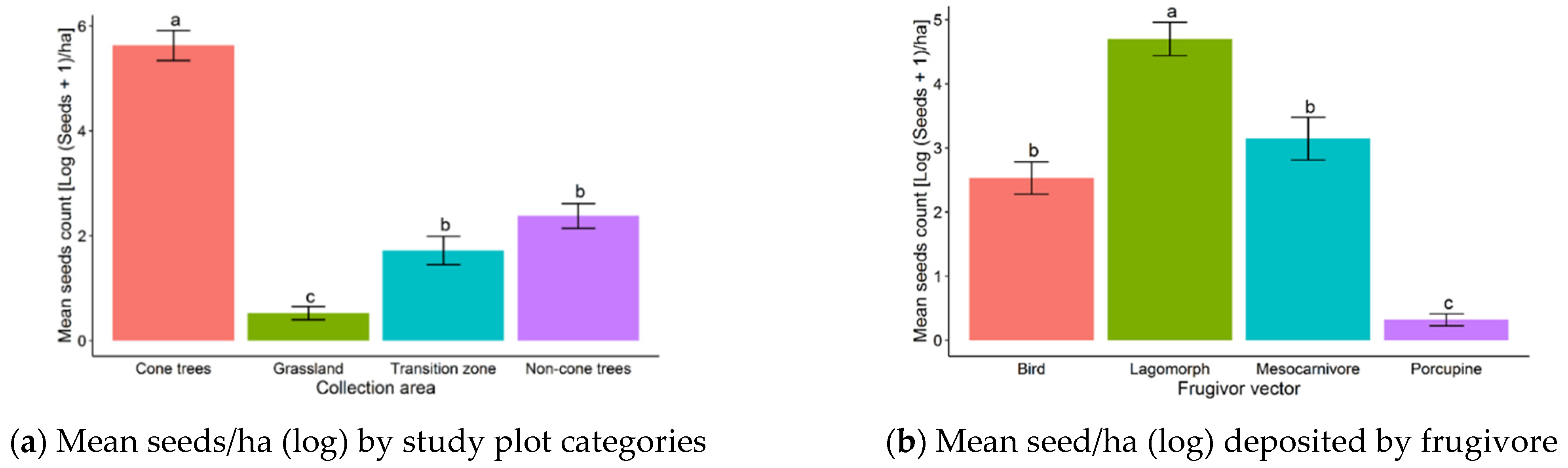
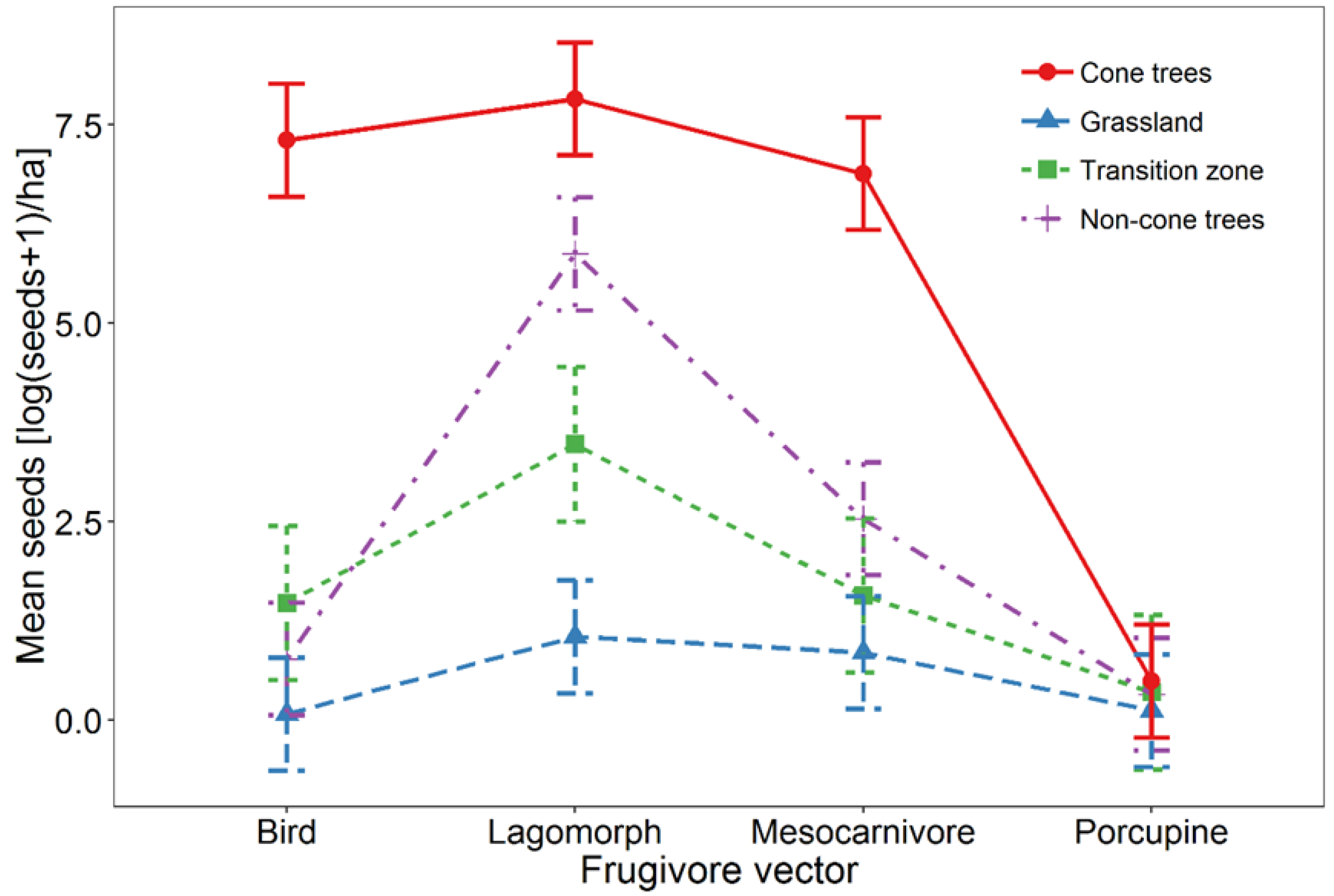
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Archer, S.R.; Andersen, E.M.; Predick, K.I.; Schwinning, S.; Steidl, R.J.; Woods, S.R. Woody plant encroachment: Causes and consequences. In Rangeland Systems; Briske, D., Ed.; Springer: College Station, TX, USA, 2017; pp. 25–84. [Google Scholar]
- Stephens, G.J.; Johnston, D.B.; Jonas, J.L.; Paschke, M.W. Understory Responses to Mechanical Treatment of Pinyon-Juniper in Northwestern Colorado. Rangel. Ecol. Manag. 2016, 69, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, B.P.; Fuhlendorf, S.D.; Walker, J.W.; Twidwell, D.; Ben Wu, X.; E Goodman, L.; Treadwell, M.; Birt, A. Saving imperiled grassland biomes by recoupling fire and grazing: A case study from the Great Plains. Front. Ecol. Environ. 2021, 20, 179–186. [Google Scholar] [CrossRef]
- Berg, M.; Sorice, M.G.; Wilcox, B.P.; Angerer, J.P.; Rhodes, E.C.; Fox, W.E. Demographic Changes Drive Woody Plant Cover Trends—An Example from the Great Plains. Rangel. Ecol. Manag. 2015, 68, 315–321. [Google Scholar] [CrossRef]
- Briske, D.D.; A Joyce, L.; Polley, H.W.; Brown, J.R.; Wolter, K.; A Morgan, J.; A McCarl, B.; Bailey, D.W. Climate-change adaptation on rangelands: Linking regional exposure with diverse adaptive capacity. Front. Ecol. Environ. 2015, 13, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Van Auken, O. Causes and consequences of woody plant encroachment into western North American grasslands. J. Environ. Manag. 2009, 90, 2931–2942. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Junipers of the World: The genus Juniperus, 4th ed.; Trafford Publishing: Bloomington, IN, USA, 2014. [Google Scholar]
- Johnsen, T.N. One-Seed Juniper Invasion of Northern Arizona Grasslands. Ecol. Monogr. 1962, 32, 187–207. [Google Scholar] [CrossRef]
- Pieper, R.D. Overstory-understory relations in pinyon-juniper woodlands in New Mexico. J. Range Manage. 1990, 43, 413–415. [Google Scholar] [CrossRef]
- Rosenstock, S.S.; Van Riper, C. Breeding Bird Responses to Juniper Woodland Expansion. Rangel. Ecol. Manag. 2001, 54, 226. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.F.; Gatewood, R.G. Proceedings: Ecology and Management of Pinyon-Juniper Communities within the Interior West, Provo, UT, USA, 15–18 September 1997; Monsen, S.B., Stevens, R., Eds.; Proc. RMRS-P-9; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999.
- Miller, R.F.; Svejcar, T.J.; Rose, J.A. Impacts of Western Juniper on Plant Community Composition and Structure. Rangel. Ecol. Manag. 2000, 53, 574. [Google Scholar] [CrossRef]
- Schott, M.R.; Pieper, R.D. Influence of Canopy Characteristics of One-Seed Juniper on Understory Grasses. Rangel. Ecol. Manag. 1985, 38, 328. [Google Scholar] [CrossRef]
- Roundy, B.A.; Young, K.; Cline, N.; Hulet, A.; Miller, R.E.; Tausch, R.J.; Chambers, J.C.; Rau, B. Pinion-juniper reduction increases soil water availability of the resource growth pool. Rangel. Ecol. Manage. 2014, 67, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Coppedge, B.R.; Engle, D.M.; Masters, R.E.; Gregory, M.S. Avian response to landscape change in fragmented southern great plains grasslands. Ecol. Applications. 2001, 11, 47–59. [Google Scholar] [CrossRef]
- Coppedge, B.R. Predicting juniper encroachment and CRP effects on avian community dynamics in southern nixed-grass prairie, USA. Biol. Conserv. 2004, 115, 431–441. [Google Scholar] [CrossRef]
- Coates, P.S.; Prochazka, B.G.; Ricca, M.A.; Gustafson, K.B.; Ziegler, P.; Casazza, M.L. Pinyon and juniper encroachment into sagebrush ecosystems impacts distribution and survival of greater sage-grouse. Rangel. Ecol. Manage. 2017, 70, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Romme, W.; Allen, J.D.; Bailey, W.L.; Baker, B.T.; Bestelmeyer, P.M.; Brown, K.S.; Eisenhart, M.L.; Floyd-Hanna, D.W.; Huffman, B.F.; Jacobs, R.F.; et al. Historical and modern disturbance regimes, stand structure, and landscape dynamics in pinyon-juniper vegetation of the western United States. Rangel. Ecol. Manag. 2009, 62, 203–222. [Google Scholar] [CrossRef] [Green Version]
- Stevens, B.F.; Erasmus, N.; Archibald, S.; Bond, W.J. Woody encroachment of 70 years in savannahs: Overgrazing, global change or extinction aftershock? Philos. Trans. R Soc. Lond B Biol. Sci. 2016, 371, 20150437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allred, B.W.; Fuhlendorf, S.D.; Smeins, F.E.; Taylor, C.A. Herbivore species and grazing intensity regulate community composition and an encroaching woody plant in semi-arid rangeland. Basic Appl. Ecology 2012, 13, 149–158. [Google Scholar] [CrossRef]
- Ganguli, A.C.; Engle, D.M.; Mayer, P.M.; Salo, L.F. Influence of resource availability on Juniperus virginiana expansion in a forest-prairie ecotone. Ecosphere 2016, 7, e01433. [Google Scholar] [CrossRef] [Green Version]
- Escribano-Avila, G.; Calviono-Cancela, M.; Pias, B.; Virgos, E.; Valladares, F.; Escudero, A. Diverse guilds provide complementary dispersal services in a woodland expansion process after land abandonment. J. Appl. Ecol. 2014, 51, 1701–1711. [Google Scholar] [CrossRef] [Green Version]
- Rumeu, B.; Elias, R.B.; Padilla, D.P. Differential seed dispersal systems of endemic junipers in two oceanic Macaronesian archipelagos: The influence of biogeographic and biological characteristics. Plant Ecology 2011, 212, 911–921. [Google Scholar] [CrossRef]
- Lezama-Delgado, E.; Sainos-Paredes, P.; López-Portillo, J.; Angeles, G.; Golubovc, J.; Martínez, A.J. Association of Juniperus deppeana (Cupressaceae: Pinales) seeds with Mexican cottontail rabbit (Sylvilagus cunicularius; Leporidae: Lagomorpha) latrines. J. Nat. Hist. 2016, 50, 2547–2555. [Google Scholar] [CrossRef]
- Salomonson, M.G. Adaptations for animal dispersal of one-seed juniper seeds. Oecologica 1978, 32, 333–339. [Google Scholar] [CrossRef] [PubMed]
- White, C.G.; Flinders, J.T.; Cates, R.G.; Blackwell, B.H.; Smith, H.D. Dietary use of Utah juniper berries by gray fox in eastern Utah. In Proceedings of the Ecology and Management of Pinyon-Juniper Communities within the Interior West: RMRS-P-9; Monson, S.B., Stevens, R., Eds.; Dept. of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999. [Google Scholar]
- Longland, W.S.; Dimitri, L.A. Are western juniper seeds dispersed through diplochory? Northwest Sci. 2016, 90, 235–243. [Google Scholar] [CrossRef]
- Dimitri, L.A.; Longland, W.S. Distribution of western juniper seeds across an ecotone and implications for dispersal. West. North Amer. Nat. 2017, 77, 212–222. [Google Scholar] [CrossRef]
- Chavez-Ramirez, F.; Slack, R.D. Carnivore Fruit-Use and Seed Dispersal of Two Selected Plant Species of the Edwards Plateau, Texas. Southwest. Nat. 1993, 38, 141. [Google Scholar] [CrossRef]
- Escribano-Ávila, G.; Pías, B.; Sanz-Pérez, V.; Virgós, E.; Escudero, A.; Valladares, F. Spanish juniper gain expansion opportunities by counting on a functionally diverse dispersal assemblage community. Ecol. Evol. 2013, 3, 3751–3763. [Google Scholar] [CrossRef] [Green Version]
- García-Cervigón, A.I.; Velázquez, E.; Wiegand, T.; Escudero, A.; Olano, J.M. Colonization in Mediterranean old-fields: The role of dispersal and plant-plant interactions. J. Veg. Sci. 2016, 28, 627–638. [Google Scholar] [CrossRef]
- Jordano, P.; García, C.; Godoy, J.A.; García-Castaño, L.L. Differential contribution of frugivores to complex seed dispersal patterns. PNAS 2007, 104, 3278–3282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traveset, A.; Verdu, M. A meta-analysis of the effect of gut treatment on seed germination. In Seed Dispersal and Frugivory: Ecology, Evolution and Conservation; Silva, W.R., Galetti, M., Eds.; CAB International: New York, NY, USA, 2002; pp. 339–350. [Google Scholar]
- Strand, E.K.; Robinson, A.P.; Bunting, S.C. Spatial patterns on the sagebrush steppe/Western juniper ecotone. Plant Ecol. 2007, 190, 159–173. [Google Scholar] [CrossRef]
- Holthuijzen, A.M.; Sharik, T.L. The avian seed dispersal of eastern red cedar. Can. J. Bot. 1985, 63, 1508–1515. [Google Scholar] [CrossRef]
- Herrera, C.M. Frugivory and seed dispersal by carnivorous mammals, and associated fruit characteristics, in undisturbed Mediterranean habitats. Oikos 1989, 55, 250–262. [Google Scholar] [CrossRef] [Green Version]
- NOAA, West. Reg. Climate Center. Available online: https://wrcc.dri.edu/Climate/west_coop_summaries.php (accessed on 17 November 2018).
- Forbes, A.C.; Allred, K.W. A field guide to the flora of New Mexico State University’s Corona Range and Livestock Research Center. NMSU Ag. Expt. Stn. Res. Rep. 2001, 745, 61. [Google Scholar]
- Santos, T.; Tellería, J.L.; Virgós, E. Dispersal of Spanish juniper Juniperus thurifera by birds and mammals in a fragmented landscape. Ecography 1999, 22, 193–204. [Google Scholar] [CrossRef]
- Tellería, J.L.; Carrascal, L.M.; Santos, T. Species abundance and migratory status affects large-scale fruit tracking in thrushes (Turdus spp.). J. Ornithol. 2014, 155, 157–164. [Google Scholar] [CrossRef]
- Jordano, P. Geographical ecology and variation of plant-seed disperser interactions: Southern Spanish junipers and frugivorous thrushes. In Vegetatio 107/108, Frugivory and Seed Dispersal: Ecological and Evolutionary Aspects; Fleming, T.H., Estrada, A., Eds.; Kluwer Academic Publishers: Dordrecht, Belgium, 1993; pp. 85–104. [Google Scholar]
- Elbroch, M. Mammal Tracks & Sign: A Guide to North American Species, 1st ed.; Stackpole Books: Mechanicsburg, PA, USA, 2003; 780p. [Google Scholar]
- Costello, C.M.; Jones, D.E.; Inman, R.M.; Inman, K.H.; Thompson, B.C.; Quigley, H.B. Relationship of variable mast production to American black bear reproductive parameters in New Mexico. Ursus 2003, 14, 1–16. [Google Scholar]
- R Core Team. 2017. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 12 February 2019).
- Salomonson, M.G.; Balda, R.P. Winter territoriality of Townsend’s Solitaires (Myadestes townsendi) in a pinon-juniper-ponderosa pine ecotone. Condor 1977, 79, 148–161. [Google Scholar] [CrossRef]
- Holthuijzen, A.M.; Sharik, T.L. The red cedar (Juniperus virginiana L.) seed shadow along a fenceline. Amer. Mid. Nat. 1985, 113, 200–202. [Google Scholar] [CrossRef]
- Stricklan, D.; Saud, P.; Cibils, A.F.; Steiner, R.L.; Cram, D.S.; Young, K.; Faist, A.M. Germination of one-seed juniper seeds distributed by different frugivore groups. Rangel. Ecol. Manage. 2020, 73, 433–440. [Google Scholar] [CrossRef]
- Schupp, E.W.; Gomez, J.M.; Jimenez, J.E.; Fuentes, M. Dispersal of Juniperus occidentalis (western juniper) seeds by frugivorous mammals on Juniper Mountain in southeastern Oregon. Great Basin Nat. 1997, 57, 74–78. [Google Scholar]
- Perea, R.; Delibes, M.; Polko, M.; Suárez-Esteban, A.; Fedriani, J.M. Context-dependent fruit–frugivore interactions: Partner identities and spatio-temporal variations. Oikos 2013, 122, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Schupp, E.W.; Heaton, H.J.; Gomez, J.M. Lagomorphs and the dispersal of seeds into communities dominated by exotic annual weeds. Great Basin Nat. 1997, 57, 253–258. [Google Scholar]
- Ilse, L.M.; Hellgren, E.C. Demographic and behavioral characteristics of North American porcupines (Erethizon dorsatum) in Pinyon-Juniper Woodlands of Texas. Amer. Mid. Nat. 2001, 146, 329–338. [Google Scholar] [CrossRef]
- Stricklan, D.; Flinders, J.T.; Cates, R.G. Factors affecting selection of winter food and roosting resources by porcupines in Utah. 1995. Great Basin Nat. 1995, 55, 29–36. [Google Scholar]
- Sweitzer, R.E. Predation or starvation: Consequences of foraging decisions by porcupines (Erethizon dorsatum). J. Mamm. 1996, 77, 1068–1077. [Google Scholar] [CrossRef] [Green Version]
- Schupp, E.W. Quantity, quality and the effectiveness of seed dispersal by animals. Vegetatio 1993, 107/108, 15–29. [Google Scholar] [CrossRef]
- Farris, E.; Canopoli, L.; Cucca, E.; Landi, S.; Maccioni, A.; Filigheddu, R. Foxes provide a direct dispersal service to Phoenician junipers in Mediterranean costal environments: Ecological and evolutionary implications. Plant Ecol. Evol. 2017, 150, 117–128. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Mean & SEM | Median | Maximum | % Presence |
|---|---|---|---|---|
| Cone Trees (under canopy) n = 60 | ||||
| Bird | 117,989 ± 19,387 | 71,474 | 844,473 | 100 |
| Lagomorph | 36,146 ± 7543 | 13,851 | 291,774 | 82 |
| Mesocarnivore | 107,200 ± 65,977 | 0 | 3,922,535 | 27 |
| Porcupine | 861 ± 391 | 0 | 19,934 | 15 |
| Cone Trees (beyond canopy) n = 60 | ||||
| Bird | 5338 ± 818 | 3193 | 26,464 | 93 |
| Lagomorph | 10,839 ± 2543 | 2543 | 119,068 | 93 |
| Mesocarnivore | 52,257 ± 10,031 | 20,503 | 375,777 | 63 |
| Porcupine | 52 ± 33 | 0 | 1818 | 08 |
| Non-Cone Tree (under canopy) n = 60 | ||||
| Bird | 896 ± 312 | 0 | 11,620 | 19 |
| Lagomorph | 5641 ± 1044 | 2312 | 38,710 | 64 |
| Mesocarnivore | 14,218 ± 8564 | 0 | 415,129 | 05 |
| Porcupine | 3029 ± 3029 | 0 | 181,734 | 02 |
| Non-Cone Tree (beyond canopy) n = 60 | ||||
| Bird | 154 ± 99 | 0 | 5798 | 12 |
| Lagomorph | 1983 ± 497 | 690 | 22,277 | 87 |
| Mesocarnivore | 10,054 ± 3005 | 0 | 117,654 | 25 |
| Porcupine | 39 ± 24 | 0 | 1234 | 05 |
| Transition Zone n = 32 * | ||||
| Bird | 128 ± 50 | 0 | 1100 | 25 |
| Lagomorph | 1260 ± 605 | 50 | 18,600 | 50 |
| Mesocarnivore | 1759 ± 947 | 0 | 23,200 | 19 |
| Porcupine | 13 ± 12 | 0 | 400 | 09 |
| Grassland Open Areas n = 60 | ||||
| Bird | 2 ± NA | 0 | 100 | 02 |
| Lagomorph | 90 ± 39 | 0 | 2000 | 18 |
| Mesocarnivore | 3325 ± 1836 | 0 | 82,100 | 08 |
| Porcupine | 23 ± NA | 0 | 1400 | 02 |
| Habitat | Scat/Pellet Presence (% Plots) | Frugivore Tests a | General Assoc. b | |||||
|---|---|---|---|---|---|---|---|---|
| B | L | M | P | Chi2 | p | CMH Statistic | p | |
| 2017 | ||||||||
| Cone trees | 92.6 | 88.9 | 85.2 | 7.4 | 62.7 | <0.01 | ||
| Non-cone trees | 15.2 | 90.9 | 15.2 | 3.0 | 75.1 | <0.01 | ||
| Open grassland | 0.0 | 26.7 | 0.9 | 0.0 | 21.5 | <0.01 | ||
| 2018 | ||||||||
| Cone trees | 90.9 | 96.9 | 45.5 | 9.1 | 70.8 | <0.01 | ||
| Non-cone tree | 7.4 | 81.5 | 37.0 | 7.4 | 44.7 | <0.01 | ||
| Transition zone c | 20.0 | 46.7 | 13.3 | 3.3 | 18.7 | <0.01 | ||
| Open grassland | 3.3 | 10.0 | 10.0 | 3.3 | 2.1 | 0.54 | ||
| Overall Frugivore by Habitat relationship d | 212.4 | <0.01 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stricklan, D.; Cibils, A.F.; Saud, P.; Steiner, R.L.; McIntosh, M.M.; Ganguli, A.C.; Cram, D.S.; Faist, A.M. Dispersal Patterns of One-Seed Juniper Seeds Contained in Mammal Scats and Bird Pellets. Forests 2022, 13, 1693. https://doi.org/10.3390/f13101693
Stricklan D, Cibils AF, Saud P, Steiner RL, McIntosh MM, Ganguli AC, Cram DS, Faist AM. Dispersal Patterns of One-Seed Juniper Seeds Contained in Mammal Scats and Bird Pellets. Forests. 2022; 13(10):1693. https://doi.org/10.3390/f13101693
Chicago/Turabian StyleStricklan, Dave, Andrés F. Cibils, Pradip Saud, Robert L. Steiner, Matthew M. McIntosh, Amy C. Ganguli, Douglas S. Cram, and Akasha M. Faist. 2022. "Dispersal Patterns of One-Seed Juniper Seeds Contained in Mammal Scats and Bird Pellets" Forests 13, no. 10: 1693. https://doi.org/10.3390/f13101693
APA StyleStricklan, D., Cibils, A. F., Saud, P., Steiner, R. L., McIntosh, M. M., Ganguli, A. C., Cram, D. S., & Faist, A. M. (2022). Dispersal Patterns of One-Seed Juniper Seeds Contained in Mammal Scats and Bird Pellets. Forests, 13(10), 1693. https://doi.org/10.3390/f13101693