
Warming Responses of Leaf Morphology Are Highly Variable among Tropical Tree Species

 , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Planting Design and Plant Material
2.3. Environmental Conditions
2.4. Leaf Sampling and Morphological Traits Measurements
2.5. Leaf Shape and Size Estimation
2.6. Determination of LMA, LD and LVA
2.7. Leaf Nutrients Analysis
2.8. Comparison of Juvenile Planted Trees with Mature and Natural Regenerated Trees
2.9. Statistics
3. Results
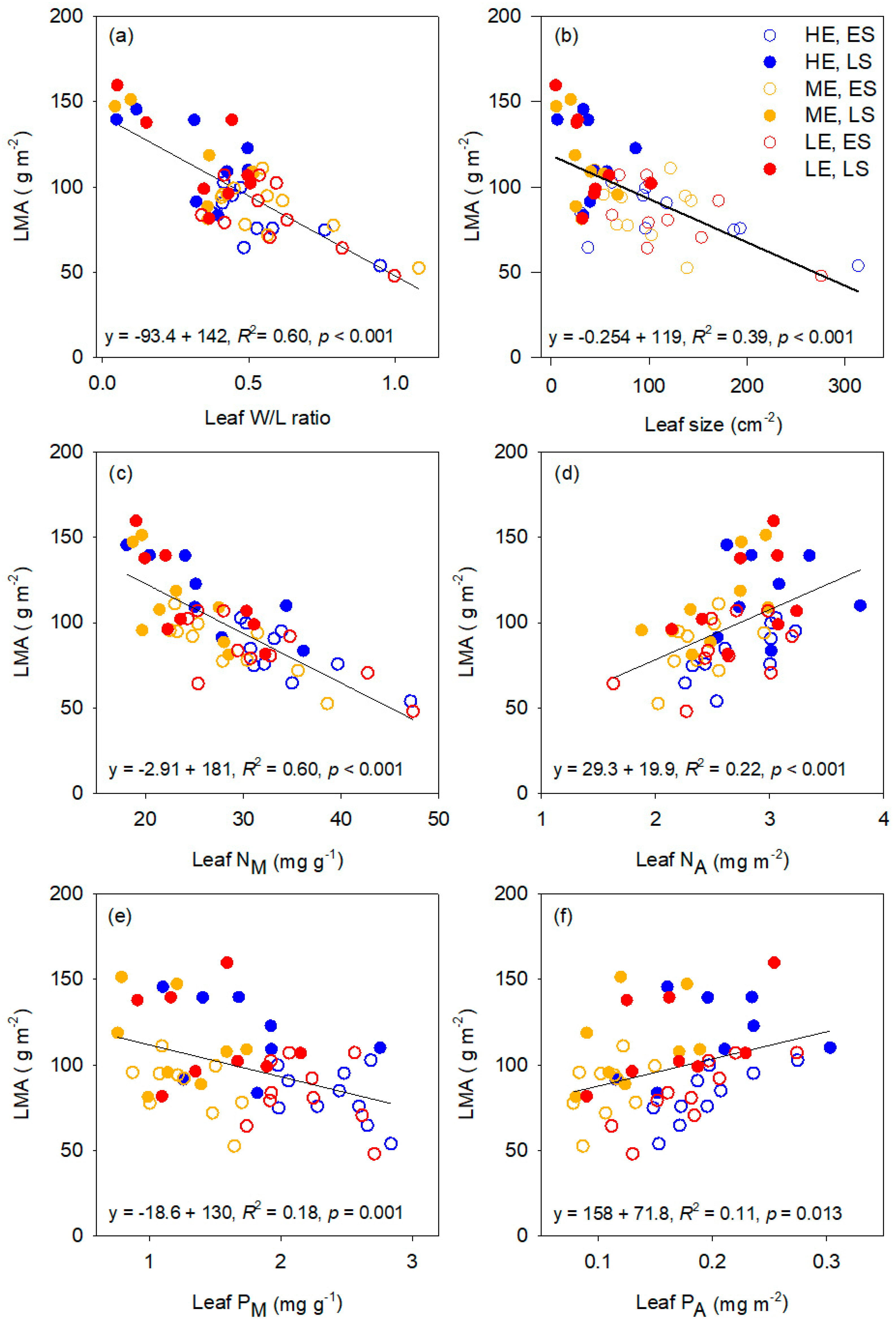
3.1. Leaf Trait Interrelationships and Variation between Species
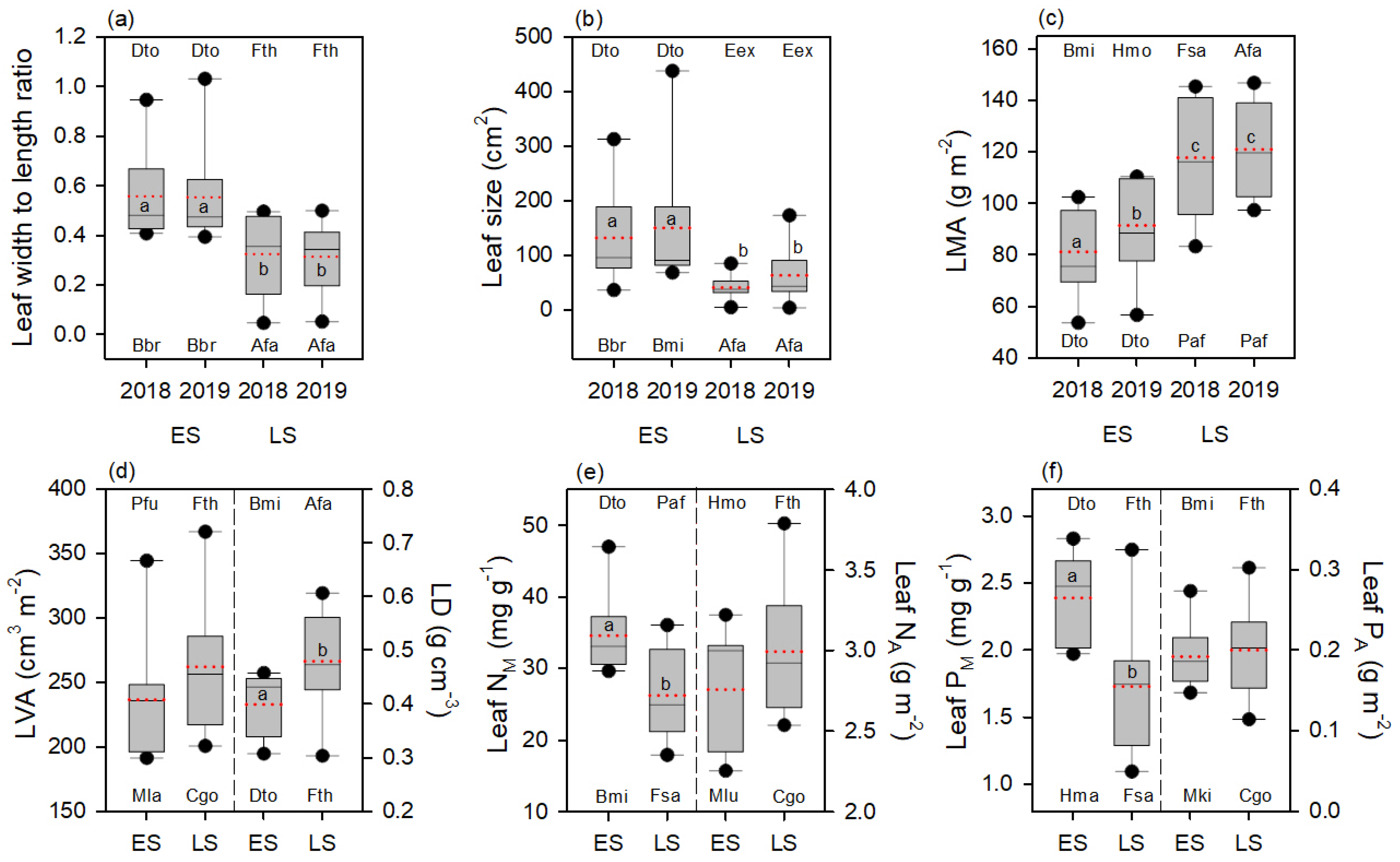
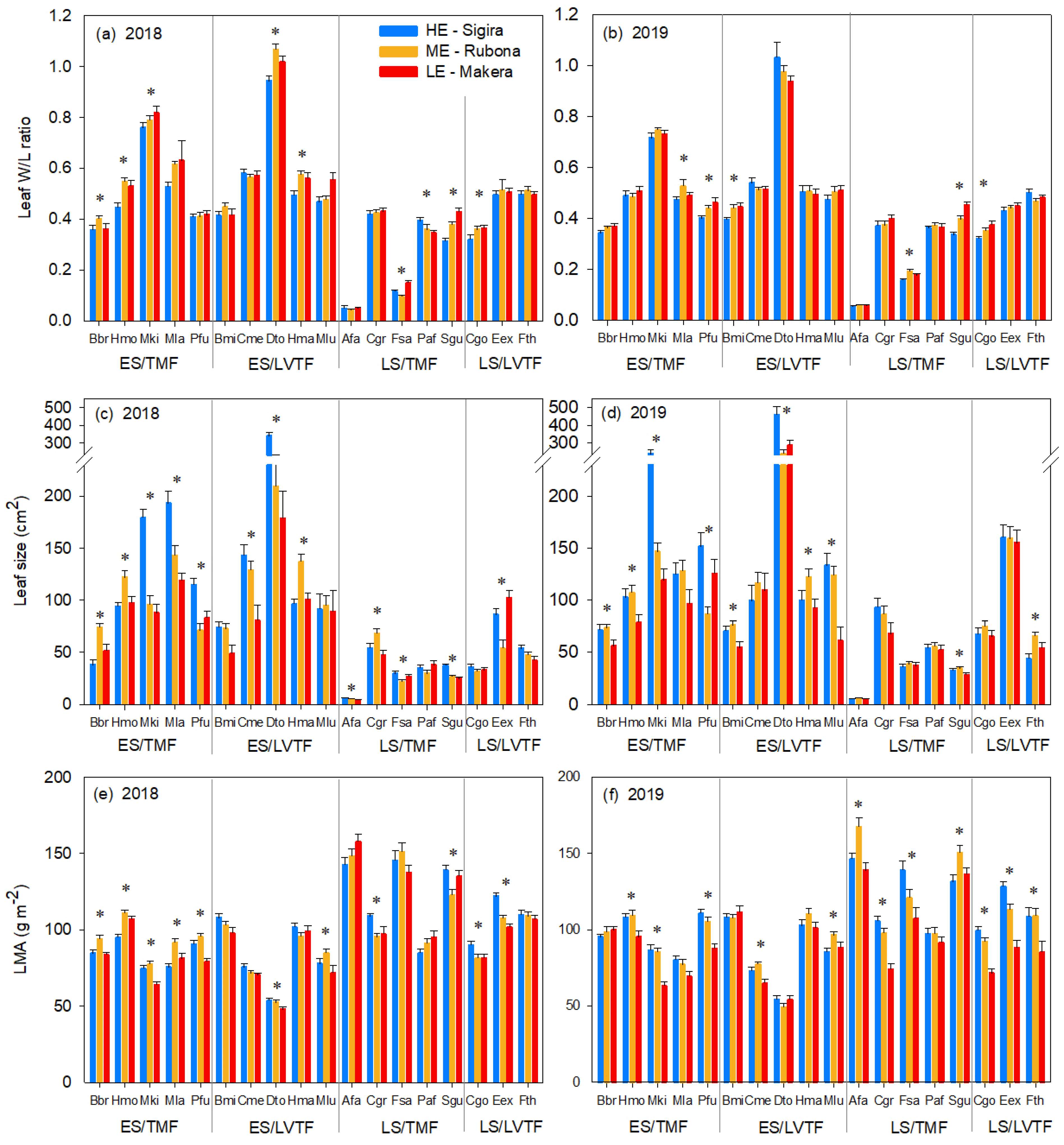
3.2. Leaf Shape, LMA and Leaf Size Responses to Sites and Warming
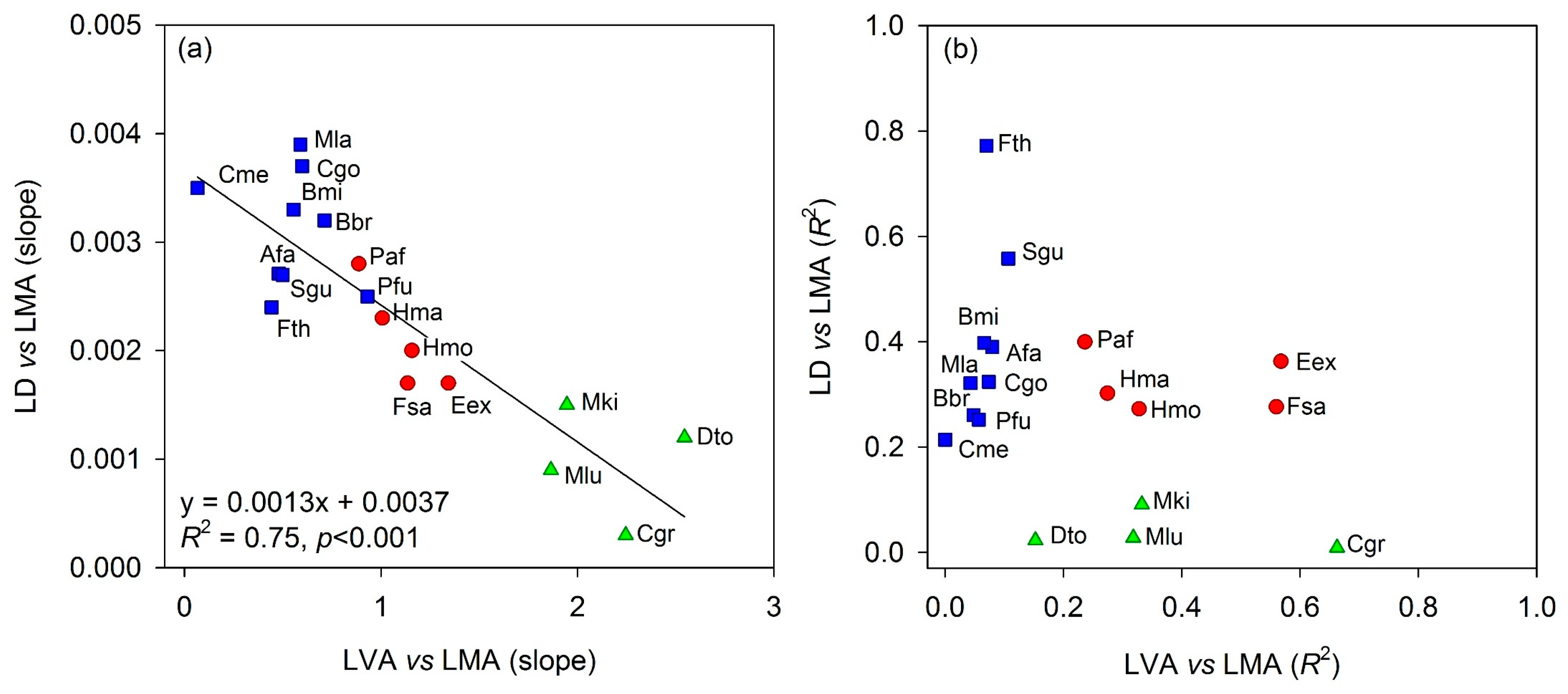
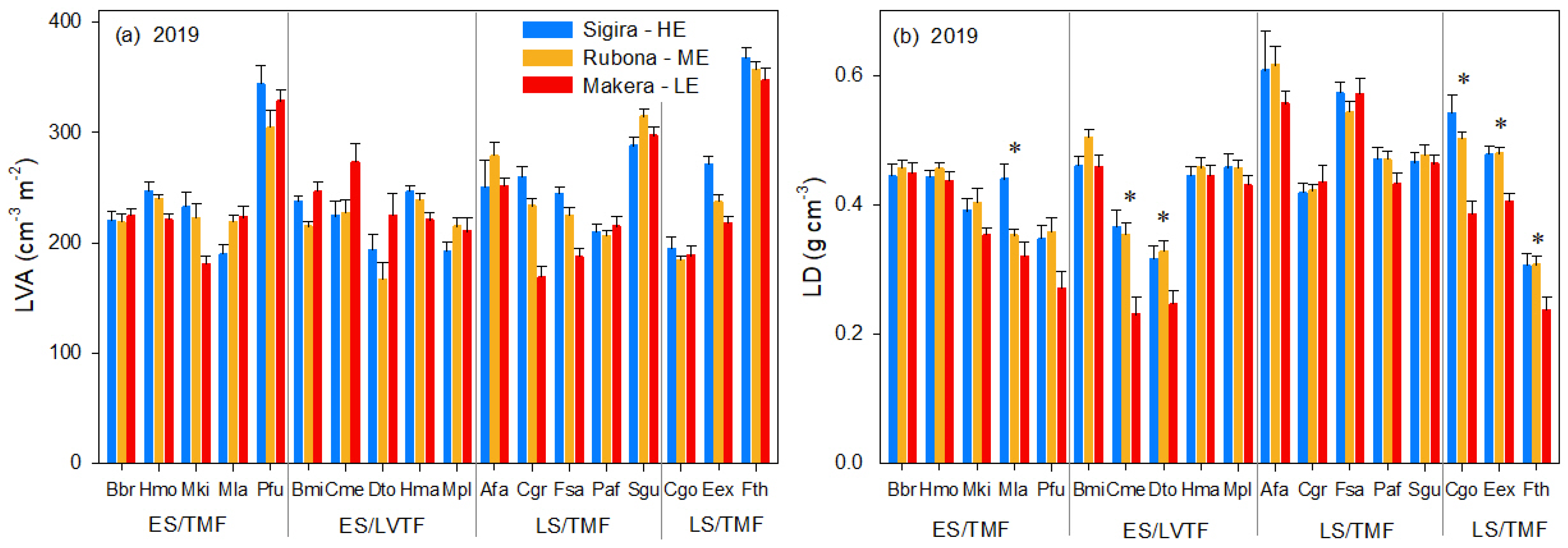
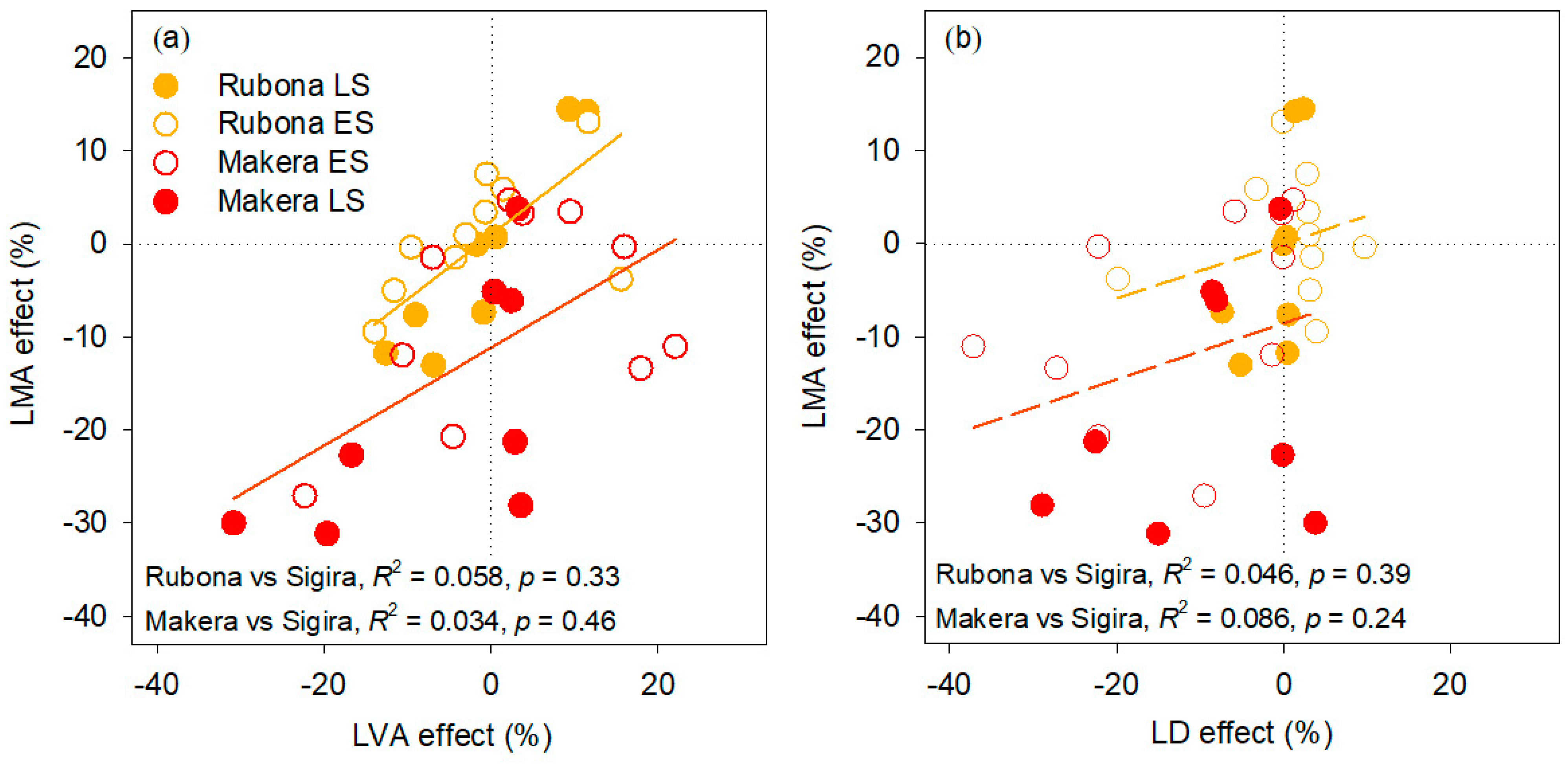
3.3. Effect of Warming on LVA and LD, and Their Contribution to the Effect on LMA
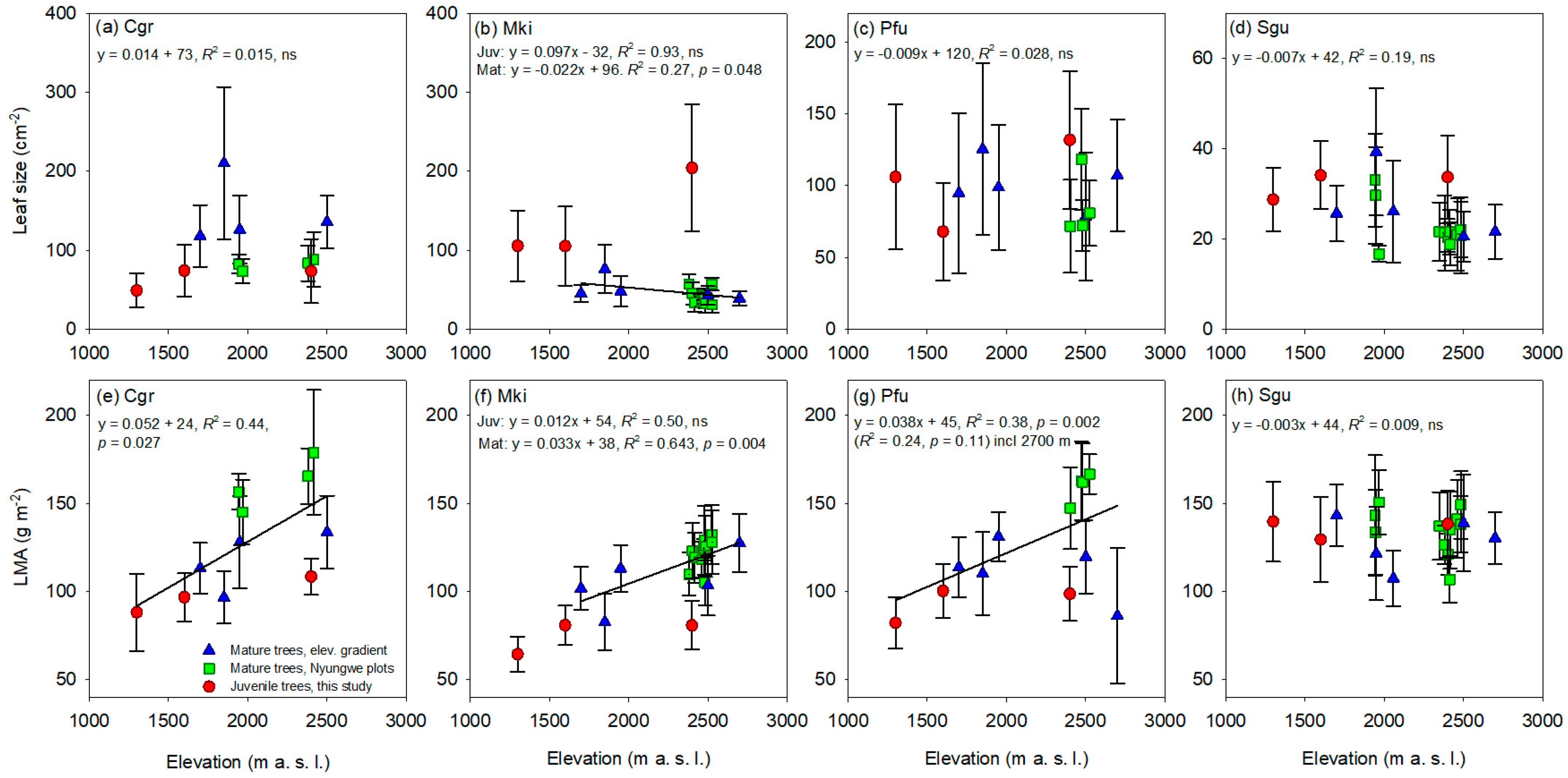
3.4. Leaf Size and LMA in Juvenile and Mature Trees at Different Elevations
4. Discussion
4.1. Characterisation of Leaf Traits in Different Species and Successional Groups
4.2. LMA Responses to Warming
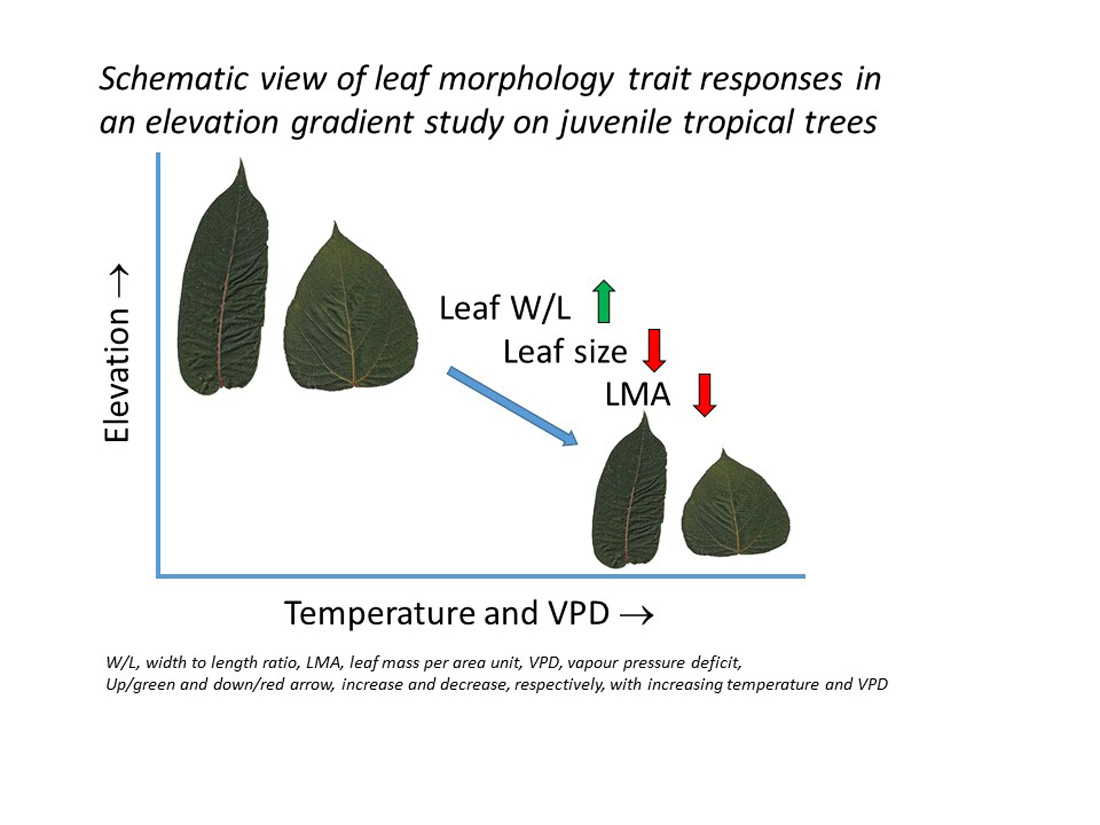
4.3. Leaf Size and Leaf Width to Length Response to Warming
4.4. Responses to Warming in Species of Different Successional Groups
4.5. LMA and Leaf Size in Juvenile and Mature Trees at Different Elevations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, J. The Cost of Leafing: Understanding the trade-offs involved for plants making leaves promises fresh insights on every scale from the plant to the planet. Nature 2006, 444, 539–541. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Reich, P.B.; Schmid, B.; Shrestha, N.; Feng, X.; Lyu, T.; Maitner, B.S.; Xu, X.; Li, Y.; Zou, D.; et al. Leaf size of woody dicots predicts ecosystem primary productivity. Ecol. Lett. 2020, 23, 1003–1013. [Google Scholar] [CrossRef]
- Marron, N.; Villar, M.; Dreyer, E.; Delay, D.; Boudouresque, E.; Petit, J.M.; Delmotte, F.M.; Guehl, J.M.; Brignolas, F. diversity of leaf traits related to productivity in 31 Populus deltoides x Populus nigra clones. Tree Physiol. 2005, 25, 425–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meziane, D.; Shipley, B. Direct and indirect relationships between specific leaf area, leaf nitrogen and leaf gas exchange. effects of irradiance and nutrient supply. Ann. Bot. 2001, 88, 915–927. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.R.C.; Jordan, G.J.; Brodribb, T.J. Acclimation to humidity modifies the link between leaf size and the density of veins and stomata. Plant Cell Environ. 2014, 37, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, K.Y. Plant responses to climate warming: Physiological adjustments and implications for plant functioning in a future, warmer world. Am. J. Bot. 2019, 106, 1049–1051. [Google Scholar] [CrossRef] [Green Version]
- Fauset, S.; Freitas, H.C.; Galbraith, D.R.; Sullivan, M.J.P.; Aidar, M.P.M.; Joly, C.A.; Phillips, O.L.; Vieira, S.A.; Gloor, M.U. Differences in leaf thermoregulation and water use strategies between three co-occurring Atlantic forest tree species. Plant Cell Environ. 2018, 41, 1618–1631. [Google Scholar] [CrossRef] [Green Version]
- Fyllas, N.M.; Bentley, L.P.; Shenkin, A.; Asner, G.P.; Atkin, O.K.; Díaz, S.; Enquist, B.J.; Farfan-Rios, W.; Gloor, E.; Guerrieri, R.; et al. Solar radiation and functional traits explain the decline of forest primary productivity along a tropical elevation gradient. Ecol. Lett. 2017, 20, 730–740. [Google Scholar] [CrossRef]
- Wright, I.A.N.J.; Westoby, M.; Reich, P.B. Wright. Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span. J. Ecol. 2002, 90, 534–543. [Google Scholar] [CrossRef] [Green Version]
- Bussotti, F. Functional leaf traits, plant communities and acclimation processes in relation to oxidative stress in trees: A critical overview. Glob. Chang. Biol. 2008, 14, 2727–2739. [Google Scholar] [CrossRef]
- Gornish, E.S.; Prather, C.M. Foliar functional traits that predict plant biomass response to warming. J. Veg. Sci. 2014, 25, 919–927. [Google Scholar] [CrossRef]
- Neyret, M.; Bentley, L.P.; Oliveras, I.; Marimon, B.S.; Marimon-Junior, B.H.; Almeida de Oliveira, E.; Barbosa Passos, F.; Castro Ccoscco, R.; dos Santos, J.; Matias Reis, S.; et al. Examining variation in the leaf mass per area of dominant species across two contrasting tropical gradients in light of community assembly. Ecol. Evol. 2016, 6, 5674–5689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussotti, F.; Pancrazi, M.; Matteucci, G.; Gerosa, G. Leaf morphology and chemistry in Fagus sylvatica (Beech) trees as affected by site factors and ozone: Results from conecofor permanent monitoring plots in Italy. Tree Physiol. 2005, 25, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Doughty, C.E.; Santos-Andrade, P.E.; Shenkin, A.; Goldsmith, G.R.; Bentley, L.P.; Blonder, B.; Díaz, S.; Salinas, N.; Enquist, B.J.; Martin, R.E.; et al. Tropical forest leaves may darken in response to climate change. Nat. Ecol. Evol. 2018, 1918–1924. [Google Scholar] [CrossRef]
- Blonder, B.; Vasseur, F.; Violle, C.; Shipley, B.; Enquist, B.J.; Vile, D. Testing models for the leaf economics spectrum with leaf and whole-plant traits in Arabidopsis thaliana. AoB Plants 2015, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shurbs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Osnas, J.L.D.; Katabuchi, M.; Kitajima, K.; Wright, S.J.; Reich, P.B.; van Bael, S.; Kraft, N.J.B.; Samaniego, M.J.; Pacala, S.W.; Lichstein, J.W. Divergent drivers of leaf trait variation within species, among species, and among functional groups. Proc. Natl. Acad. Sci. USA 2018, 115, 5480–5485. [Google Scholar] [CrossRef] [Green Version]
- Mujawamariya, M.; Wittemann, M.; Manishimwe, A.; Ntirugulirwa, B.; Zibera, E.; Nsabimana, D.; Wallin, G.; Uddling, J.; Dusenge, M.E. Complete or overcompensatory thermal acclimation of leaf dark respiration in African tropical trees. New Phytol. 2021, 229, 2548–2561. [Google Scholar] [CrossRef]
- Feng, Y.L.; Cao, K.F.; Zhang, J.L. Photosynthetic characteristics, dark respiration, and leaf mass per unit area in seedlings of four tropical tree species grown under three irradiances. Photosynthetica 2004, 42, 431–437. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Villar, R.; Ruiz-Robleto, J.; Ubera, J.L.; Poorter, H. Exploring variation in leaf mass per area (LMA) from leaf to cell: An anatomical analysis of 26 woody species. Am. J. Bot. 2013, 100, 1969–1980. [Google Scholar] [CrossRef] [PubMed]
- Coble, A.P.; Cavaleri, M.A. Vertical leaf mass per area gradient of mature sugar maple reflects both height-driven increases in vascular tissue and light-driven increases in palisade layer thickness. Tree Physiol. 2017, 37, 1337–1351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, L.; Shen, W. Dramatic altitudinal variations in leaf mass per area of two plant growth forms at extreme heights. Ecol. Indic. 2020, 110, 105890. [Google Scholar] [CrossRef]
- Lambers, H.I.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar] [CrossRef]
- Choong, M.F.; Lucas, P.W.; Ong, J.S.Y.; Pereira, B.; Tan, H.T.W.; Turner, I.M. Leaf fracture toughness and sclerophylly: Their correlations and ecological implications. New Phytol. 1992, 121, 597–610. [Google Scholar] [CrossRef]
- Cavaleri, M.A.; Reed, S.C.; Smith, W.K.; Wood, T.E. Urgent need for warming experiments in tropical forests. Glob. Chang. Biol. 2015, 21, 2111–2121. [Google Scholar] [CrossRef]
- Chapman, C.A.; Galán-Acedo, C.; Gogarten, J.F.; Hou, R.; Lawes, M.J.; Omeja, P.A.; Sarkar, D.; Sugiyama, A.; Kalbitzer, U. A 40-year evaluation of drivers of African rainforest change. For. Ecosyst. 2021, 8, 66. [Google Scholar] [CrossRef]
- Malhi, Y.; Silman, M.; Salinas, N.; Bush, M.; Meir, P.; Saatchi, S. Introduction: Elevation gradients in the tropics: Laboratories for ecosystem ecology and global change research. Glob. Chang. Biol. 2010, 16, 3171–3175. [Google Scholar] [CrossRef]
- Cavaleri, M.A.; Oberbauer, S.F.; Clark, D.B.; Clark, D.A.; Ryan, M.G. Height is more important than light in determining leaf morphology in a tropical forest. Ecology 2010, 91, 1730–1739. [Google Scholar] [CrossRef]
- Milla, R.; Reich, P.B. Multi-trait interactions, not phylogeny, fine-tune leaf size reduction with increasing altitude. Ann. Bot. 2011, 107, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Osnas, J.L.D.; Lichstein, J.W.; Reich, P.B.; Pacala, S.W. Global leaf trait relationships: Mass, area, and the leaf economics spectrum. Science 2013, 340, 741–744. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Matson, P.A.; Turner, D.R. Elevational and age gradients in Hawaiian montane rainforest: Foliar and soil nutrients. Oecologia 1988, 77, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, K.; Aiba, S.I. Ecosystem structure and productivity of tropical rain forests along altitudinal gradients with contrasting soil phosphorus pools on Mount Kinabalu, Borneo. J. Ecol. 2002, 90, 37–51. [Google Scholar] [CrossRef]
- Körner, C. The nutritional status of plants from high altitudes—A Worldwide Comparison. Oecologia 1989, 81, 379–391. [Google Scholar] [CrossRef]
- Tanner, E.V.J.; Vitousek, P.M.; Cuevas, E. Experimental investigation of nutrient limitation of forest growth on wet tropical mountains. Ecology 1998, 79, 10–22. [Google Scholar] [CrossRef]
- van De Weg, M.; Meir, P.; Grace, J.; Atkin, O.K. Altitudinal variation in leaf mass per unit area, leaf tissue density and foliar nitrogen and phosphorus content along an Amazon-Andes gradient in Peru. Plant Ecol. Divers. 2009, 2, 243–254. [Google Scholar] [CrossRef]
- Tserej, O.; Feeley, K.J. Variation in leaf temperatures of tropical and subtropical trees are related to leaf thermoregulatory traits and not geographic distributions. Biotropica 2021, 53, 868–878. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Weiser, M.D.; McDowell, N.G.; Zhou, J.; Kaspari, M.; Helliker, B.R.; Enquist, B.J. The energetic and carbon economic origins of leaf thermoregulation. Nat. Plants 2016, 2, 1–9. [Google Scholar] [CrossRef]
- Vårhammar, A.; Wallin, G.; McLean, C.M.; Dusenge, M.E.; Medlyn, B.E.; Hasper, T.B.; Nsabimana, D.; Uddling, J. Photosynthetic temperature responses of tree species in Rwanda: Evidence of pronounced negative effects of high temperature in montane rainforest climax species. New Phytol. 2015, 206, 1000–1012. [Google Scholar] [CrossRef]
- Leigh, A.; Sevanto, S.; Close, J.D.; Nicotra, A.B. The influence of leaf size and shape on leaf thermal dynamics: Does theory hold up under natural conditions? Plant Cell Environ. 2017, 40, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Ntawuhiganayo, E.B.; Uwizeye, F.K.; Zibera, E.; Dusenge, M.E.; Ziegler, C.; Ntirugulirwa, B.; Nsabimana, D.; Wallin, G.; Uddling, J. Traits controlling shade tolerance in tropical montane trees. Tree Physiol. 2020, 40, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichlos, S.A.; Wilson, C.R.; Lusk, C.H. Differential effects of elevation on leaf size of overstorey and understorey species in a temperate rainforest. N. Zeal. J. Bot. 2019, 57, 39–49. [Google Scholar] [CrossRef]
- Li, Y.; Zou, D.; Shrestha, N.; Xu, X.; Wang, Q.; Jia, W.; Wang, Z. Spatiotemporal variation in leaf size and shape in response to climate. J. Plant Ecol. 2020, 13, 87–96. [Google Scholar] [CrossRef]
- Peppe, D.J.; Royer, D.L.; Cariglino, B.; Oliver, S.Y.; Newman, S.; Leight, E.; Enikolopov, G.; Fernandez-Burgos, M.; Herrera, F.; Adams, J.M.; et al. Sensitivity of leaf size and shape to climate: Global patterns and paleoclimatic applications. New Phytol. 2011, 190, 724–739. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.L. Change and determinism in tropical forest succession. In Tropical forest Community Ecology; Carson, W.P., Schnitzer, S.A., Schwarz, E., Eds.; Wiley Blackwell: Oxford, UK, 2008; pp. 384–408. [Google Scholar]
- Coelho, P.A.; Santos, P.F.; de Paiva Paula, E.; Apgaua, D.M.G.; Madeira, B.G.; Menino, G.C.D.; Nunes, Y.R.F.; Santos, R.M.; Tng, D.Y.P. Tree succession across a seasonally dry tropical forest and forest-savanna ecotone in northern Minas Gerais, Brazil. J. Plant Ecol. 2016, 10, rtw091. [Google Scholar] [CrossRef]
- Fan, K.; Tao, J.; Zang, L.; Yao, J.; Huang, J.; Lu, X.; Ding, Y.; Xu, Y.; Zang, R. Changes in plant functional groups during secondary succession in a tropical montane rain forest. Forests 2019, 10, 1134. [Google Scholar] [CrossRef] [Green Version]
- Lohbeck, M.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Poorter, L.; Bongers, F. Functional trait strategies of trees in dry and wet tropical forests are similar but differ in their consequences for succession. PLoS ONE 2015, 10, e0123741. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Vargas, G.; Perroni, Y.; López-Acosta, J.C.; Noa-Carrazana, J.C.; Sánchez-Velásquez, L.R. Do the distribution patterns of plant functional traits change during early secondary succession in tropical montane cloud forests? Acta Oecol. 2019, 95, 26–35. [Google Scholar] [CrossRef]
- Sterck, F.; Markesteijn, L.; Schieving, F.; Poorter, L. Functional traits determine trade-offs and niches in a tropical forest community. Proc. Natl. Acad. Sci. USA 2011, 108, 20627–20632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado-Junior, J.A.; Schiavini, I.; Vale, V.S.; Arantes, C.S.; van der Sande, M.T.; Lohbeck, M.; Poorter, L. Conservative species drive biomass productivity in tropical dry forests. J. Ecol. 2016, 104, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Cheesman, A.W.; Winter, K. Growth response and acclimation of CO2 exchange characteristics to elevated temperatures in tropical tree seedlings. J. Exp. Bot. 2013, 64, 3817–3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slot, M.; Winter, K. High tolerance of tropical sapling growth and gas exchange to moderate warming. Funct. Ecol. 2018, 32, 599–611. [Google Scholar] [CrossRef]
- Valladares, F.; Wright, S.J.; Lasso, E.; Kitajima, K.; Pearcy, R.W. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology 2000, 81, 1925–1936. [Google Scholar] [CrossRef]
- Thomas, S.C.; Ickes, K. Ontogenetic Changes in leaf size in Malaysian rain forest trees. Biotropica 1995, 27, 427–434. [Google Scholar] [CrossRef]
- Ring, U.; Albrecht, C.; Schrenk, F. The East African rift system: Tectonics, climate and biodiversity. In Mountains, Climate and Biodiversity; Hoorn, C., Perrigo, A., Antonelli, A., Eds.; Wiley: Hoboken, NJ, USA, 2018; pp. 391–412. [Google Scholar]
- Camberlin, P. Climate of eastern Africa. In Oxford Research Encyclopedias; Oxford University Press: Oxford, UK, 2018. [Google Scholar] [CrossRef]
- IPCC—Intergovernmental Panel on Climate Change. Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5°C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; World Meteorological Organization: Geneva, Switzerland, 2018. [Google Scholar]
- IPCC—Intergovernmental Panel on Climate Change. Climate Change 2014—Impacts. Adaptation and Vulnerability: Part B: Regional Aspects: Working Group II Contribution to the IPCC Fifth Assessment Report; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef] [Green Version]
- Bonal, D.; Burban, B.; Stahl, C.; Wagner, F.; Hérault, B. The response of tropical rainforests to drought—lessons from recent research and future prospects. Ann. For. Sci. 2016, 73, 27–44. [Google Scholar] [CrossRef] [Green Version]
- Malhi, Y.; Doughty, C.; Galbraith, D. The allocation of ecosystem net primary productivity in tropical forests. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3225–3245. [Google Scholar] [CrossRef] [Green Version]
- Cuni-Sanchez, A.; Sullivan, M.; Platts, P.; Lewis, S.; Marchant, R.; Imani, G.; Hubau, W.; Abiem, I.; Adhikari, H.; Albrecht, T.; et al. High above-ground carbon stock of African tropical montane forests. Nature 2021, 596, 536–542. [Google Scholar] [CrossRef]
- Nyirambangutse, B.; Zibera, E.; Uwizeye, F.K.; Nsabimana, D.; Bizuru, E.; Pleijel, H.; Uddling, J.; Wallin, G. Carbon stocks and dynamics at different successional stages in an Afromontane tropical forest. Biogeosciences 2017, 14, 1285–1303. [Google Scholar] [CrossRef] [Green Version]
- Kindt, R.; Lillesø, J.P.B.; van Breugel, P.; Bingham, M.; Demissew, S.; Dudley, C.; Friis, I.; Gachathi, F.; Kalema, J.; Mbago, F.; et al. Correspondence in forest species composition between the vegetation map of Africa and higher resolution maps for seven African countries. Appl. Veg. Sci. 2014, 17, 162–171. [Google Scholar] [CrossRef]
- Ellis, B.; Daly, D.C.; Hickery, L.J.; Johnson, K.R.; Mitchell, J.D.; Wilf, P.; Wing, S.L. Manual of Leaf Architecture; Cornell University Press: Ithaca, NY, USA, 2009; p. 190. [Google Scholar]
- Shi, P.; Liu, M.; Yu, X.; Gielis, J.; Ratkowsky, D.A. Proportional relationship between leaf area and the product of leaf length and width of four types of special leaf shapes. Forests 2019, 10, 178. [Google Scholar] [CrossRef] [Green Version]
- Laurans, M.; Martin, O.; Nicolini, E.; Vincent, G. Functional traits and their plasticity predict tropical trees regeneration niche even among species with intermediate light requirements. J. Ecol. 2012, 100, 1440–1452. [Google Scholar] [CrossRef] [Green Version]
- Mujawamariya, M.; Manishimwe, A.; Ntirugulirwa, B.; Zibera, E.; Ganszky, D.; Ntawuhiganayo Bahati, E.; Nyirambangutse, B.; Nsabimana, D.; Wallin, G.; Uddling, J. Climate sensitivity of tropical trees along an elevation gradient in rwanda. Forests 2018, 9, 647. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Niklas, K.J.; Wan, Y.; Hölscher, D.; Hui, C.; Ding, Y.; Shi, P. Leaf shape influences the scaling of leaf dry mass vs. area: A test case using bamboos. Ann. For. Sci. 2020, 77, 11. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Tissue, D.T.; Li, X.; Liu, S.; Chu, G.; Zhou, G.; Li, Y.; Zheng, M.; Meng, Z.; Liu, J. Long-term effects of 7-year warming experiment in the field on leaf hydraulic and economic traits of subtropical tree species. Glob. Chang. Biol. 2020, 26, 7144–7157. [Google Scholar] [CrossRef] [PubMed]
- Vemmos, S.N.; Petri, E.; Stournaras, V. Seasonal changes in photosynthetic activity and carbohydrate content in leaves and fruit of three fig cultivars (Ficus carica L.). Sci. Hortic. 2013, 160, 198–207. [Google Scholar] [CrossRef]
- Bhusal, N.; Bhusal, S.J.; Yoona, T.M. Comparisons of physiological and anatomical characteristics between two cultivars in bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- Mujawamariya, M. Climate Change Sensitivity of Photosynthesis and Respiration in Tropical Trees. Ph.D. Thesis, University of Rwanda, Kigali, Rwanda, University of Gothenburg, Gothenburg, Sweden, 2021. [Google Scholar]
- de la Riva, E.G.; Olmo, M.; Poorter, H.; Ubera, J.S.; Villar, R. Leaf mass per area (LMA) and its relationship with leaf structure and anatomy in 34 mediterranean woody species along a water availability gradient. PLoS ONE 2016, 11, e0148788. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü.; Kull, O.; Tenhunen, J.D. Variability in leaf morphology and chemical composition as a function of canopy light environment in coexisting deciduous trees. Int. J. Plant Sci. 1999, 160, 837–848. [Google Scholar] [CrossRef]
- Sancho-Knapik, D.; Escudero, A.; Mediavilla, S.; Scoffoni, C.; Zailaa, J.; Cavender-Bares, J.; Álvarez-Arenas, T.G.; Molins, A.; Alonso-Forn, D.; Ferrio, J.P.; et al. Deciduous and evergreen oaks show contrasting adaptive responses in leaf mass per area across environments. New Phytol. 2021, 230, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Wang, M.; Lyu, M.; Niklas, K.J.; Zhong, Q.; Li, M.; Cheng, D. stem and leaf growth rates define the leaf size vs. number trade-off. AoB Plants 2019, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rozendaal, D.M.A.; Hurtado, V.H.; Poorter, L. Plasticity in Leaf Traits of 38 tropical tree species in response to light; relationships with light demand and adult stature. Funct. Ecol. 2006, 20, 207–216. [Google Scholar] [CrossRef]
- Ishida, A.; Yazaki, K.; Ang, L.H. Ontogenetic transition of leaf physiology and anatomy from seedlings to mature trees of a rain forest pioneer tree, Macaranga gigantea. Tree Physiol. 2005, 25, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.C.; Winner, W.E. Photosynthetic differences between saplings and adult trees: An integration of field results by meta-analysis. Tree Physiol. 2002, 22, 117–127. [Google Scholar] [CrossRef]
- Brito-Rocha, E.; Schilling, A.C.; Dos Anjos, L.; Piotto, D.; Dalmolin, A.C.; Mielke, M.S. regression models for estimating leaf area of seedlings and adult individuals of neotropical rainforest tree species. Braz. J. Biol. 2016, 76, 983–989. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site: | Sigira (HE) | Rubona (ME) | Makera (LE) |
|---|---|---|---|
| Site characteristics | |||
| Elevation (m a.s.l.) | 2400 | 1600 | 1300 |
| Latitude | S 2°30′54″ | S 2°28′30″ | S 2°6′31″ |
| Longitude | E 29°23′44″ | E 29°46′49″ | E 30°51′16″ |
| Potential Natural Vegetation 1 | Montane forest | Lake Victoria Transitional rain forest | Evergreen and semi-evergreen bushland and thicket |
| Weather | |||
| MAP (mm year−1) | 2144 ± 61 | 1672 ± 136 | 1106 ± 33 |
| MAT (°C) | 15.2 ± 0.1 | 20.0 ± 0.0 | 20.6 ± 0.1 |
| T air day (°C) | 17.1 ± 0.2 | 22.4 ± 0.1 | 24.0 ± 0.3 |
| T air night (°C) | 13.3 ± 0.1 | 17.5 ± 0.1 | 16.9 ± 0.03 |
| T air 99%ile (°C) | 23.1 ± 0.4 | 28.4 ± 0.4 | 31.2 ± 0.4 |
| T air 1%ile (°C) | 10.9 ± 0.3 | 13.4 ± 0.2 | 10.9 ± 0.8 |
| VPD day (kPa) | 0.51 ± 0.03 | 1.03 ± 0.01 | 1.14 ± 0.03 |
| VPD 99%ile (kPa) | 1.5 ± 0.004 | 2.5 ± 0.11 | 3.0 ± 0.16 |
| PPFD day (µmol m−2 s−1) | 611 ± 66 | 764 ± 62 | 740 ± 31 |
| PPFD 99%ile (µmol m−2 s−1) | 1683 ± 134 | 1884 ± 135 | 1841 ± 104 |
| Wind speed (m s−1) | 0.6 | 0.7 | 0.3 |
| Wind speed gusts (m s−1) | 1.2 | 1.2 | 0.7 |
| Wind speed gusts 99%ile (m s−1) | 3.5 | 4.0 | 3.0 |
| Soil properties | |||
| T soil (°C) | 16.7 | 22.4 | 22.4 |
| SWC (m3 m−3) | 0.29 | 0.19 | 0.20 |
| SWC 1%ile (m3 m−3) | 0.12 | 0.09 | 0.10 |
| SBD (g cm−3) | 1.0 ± 0.0 | 1.6 ± 0.1 | 1.4 ± 0.1 |
| Texture | Clay | Sandy clay | Clay loam |
| pH (Water) | 4.2 ± 0.2 | 5.0 ± 0.1 | 6.1 ± 0.1 |
| pH (KCl) | 3.4 ± 0.1 | 3.9 ± 0.0 | 5.3 ± 0.2 |
| Available P (µg g−1) | 12.1 ± 3.9 | 14.1 ± 0.7 | 8.2 ± 2.5 |
| NH4+ (µg g−1) | 11.3 ± 2.9 | 10.1 ± 2.3 | 7.1 ± 1.0 |
| NO3− (µg g−1) | 28.3 ± 1.9 | 7.2 ± 3.0 | 24.1 ± 4.6 |
| Tot N (mg g−1) | 2.73 ± 0.35 | 2.00 ± 0.39 | 1.26 ± 0.04 |
| Tot P (mg g−1) | 0.65 ± 0.02 | 0.14 ± 0.04 | 0.27 ± 0.02 |
| Org C (mg g−1) | 38.6 ± 1.1 | 27.1 ± 3.1 | 27.3 ± 2.4 |
| Code | Scientific Name and Author 1 | Family Name 1 | FT 2 | SG | Distribution 3 | Plant Functional Type 4 | Leaf Type & Shape 5 | K |
|---|---|---|---|---|---|---|---|---|
| Afa | Afrocarpus falcatus, (Thunb.) C.N.Page | Podocarpaceae | TMF | LS | Eastern and Southern Africa | Evergreen | Simple alternate, Oblong linear | 0.85 |
| Bbr | Bridelia brideliifolia, (Pax) Fedde | Euphorbiaceae/Phyllanthaceae | TMF | ES | Eastern and Southern Africa | Semi-deciduous | Simple alternate, Elliptic | 0.76 |
| Bmi | Bridelia micrantha, (Hochst.) Baill | Euphorbiaceae/Phyllanthaceae | LVTF | ES | Sub-Sahara Africa | Semi-deciduous | Simple alternate, Elliptic | 0.75 |
| Cgo | Chrysophyllum gorungosanum, Engl | Sapotaceae | LVTF | LS | Pantropical Africa mountains | Evergreen | Simple alternate, Obovate | 0.67 |
| Cgr | Carapa grandiflora, Sprague | Meliaceae | TMF | LS | Albertine Rift and west Africa mountains | Evergreen | Pinnately Compound, Oblong | 0.75 |
| Cme | Croton megalocarpus, Hutch. | Euphorbiaceae | LVTF | ES | Pantropical Africa | Semi-deciduous | Simple opposite, Ovate | 0.71 |
| Dto | Dombeya torrida, (J.F.Gmel.) Bamps | Sterculiaceae/Malvaceae | LVTF | ES | East Africa mountains | Semi-deciduous | Simple alternate, Elliptic lobate | 0.72 |
| Eex | Entandrophragma excelsum, (Dawe & Sprague) Sprague | Meliaceae | LVTF | LS | East Africa mountains | Evergreen | Pinnately Compound, Elliptic | 0.75 |
| Fsa | Faurea saligna, Harv | Proteaceae | TMF | LS | Eastern and Southern Africa | Evergreen | Simple alternate, Oblong acute | 0.69 |
| Fth | Ficus thonningii, Blume | Moraceae | LVTF | LS * | Pantropical | Semi-deciduous | Simple alternate, Elliptic | 0.71 |
| Hma | Harungana madagascariensis, Lam. ex Poir | Hypericaceae | LVTF | ES | Pantropical Africa | Semi-deciduous | Simple opposite, Ovate | 0.70 |
| Hmo | Harungana montana, Spirlet | Hypericaceae | TMF | ES | Albertine Rift | Semi-deciduous | Simple opposite, Ovate | 0.70 |
| Mki | Macaranga kilimandscharica, Pax | Euphorbiaceae | TMF | ES | East Africa mountains | Semi-deciduous | Simple alternate, Ovate truncate | 0.66 |
| Mla | Maesa lanceolata, Forssk. | Myrsinaceae/Primulaceae | TMF | ES | Sub-Sahara Africa and Madagascar | Semi-deciduous | Simple alternate, Ovate | 0.67 |
| Mlu | Markhamia lutea, (Benth.) K.Schum | Bignoniaceae | LVTF | ES * | Paleotropical | Evergreen | Pinnately Compound | 0.71 |
| Paf | Prunus africana, (Hook.f.) Kalkman | Rosaceae | TMF | LS | Sub-Sahara Africa and Madagascar | Evergreen | Simple alternate, Oblong | 0.73 |
| Pfu | Polyscias fulva, (Hiern) Harms | Araliaceae | TMF | ES | Pantropical Africa | Semi-deciduous | Pinnately Compound, Oblong | 0.79 |
| Sgu | Syzygium guineense, (Willd.) DC. | Myrtaceae | TMF | LS | Sub-Sahara Africa and Madagascar | Evergreen | Simple opposite, Elliptic | 0.67 |
| Leaf L/W Ratio | Leaf Size | LMA | LD | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | 2019 | ||||||||||||||||||||||||||||||||||||||
| Spec | SG | FT | C | P | H | M | L | C | P | H | M | L | C | P | H | M | L | C | P | H | M | L | C | P | H | M | L | C | P | H | M | L | C | P | H | M | L | |||||||
| Bbr | ES | TMF | h | 0.013 | ab | a | b | → | 0.101 | h | <0.001 | a | b | a | → | 0.028 | a | a | b | ↘ | 0.003 | a | b | a | → | 0.468 | 0.845 | |||||||||||||||||
| Hmo | ES | TMF | h | <0.001 | a | b | ab | → | 0.369 | 0.001 | a | b | a | → | h | 0.016 | ab | a | b | ↘ | <0.001 | a | b | b | ↗ | 0.006 | a | a | b | ↘ | 0.528 | |||||||||||||
| Mki | ES | TMF | 0.013 | a | ab | b | ↗ | 0.124 | h | <0.001 | a | b | b | ↘ | <0.001 | a | b | b | ↘ | <0.001 | a | a | b | ↘ | <0.001 | a | a | b | ↘ | 0.162 | ||||||||||||||
| Mla | ES | TMF | 0.077 | 0.047 | a | b | ab | → | <0.001 | a | b | b | ↘ | 0.127 | h | <0.001 | a | b | ab | → | 0.067 | <0.001 | a | b | b | ↘ | ||||||||||||||||||
| Pfu | ES | TMF | 0.785 | 0.003 | a | b | b | ↗ | h | <0.001 | a | b | b | ↘ | 0.001 | a | b | b | ↘ | <0.001 | a | a | b | ↘ | <0.001 | a | a | b | ↘ | h | 0.065 | |||||||||||||
| Bmi | ES | LVTF | 0.143 | 0.023 | a | ab | b | ↗ | h | 0.107 | 0.009 | a | a | b | ↘ | h | 0.137 | 0.606 | 0.060 | |||||||||||||||||||||||||
| Cme | ES | LVTF | 0.522 | 0.366 | h | 0.036 | a | ab | b | ↘ | h | 0.563 | 0.075 | 0.001 | a | a | b | ↘ | h | 0.003 | a | a | b | ↘ | ||||||||||||||||||||
| Dto | ES | LVTF | h | <0.001 | a | b | ab | → | 0.164 | h | <0.001 | a | b | b | ↘ | <0.001 | a | b | b | ↘ | 0.011 | a | a | b | ↘ | h | 0.230 | 0.008 | a | a | b | ↘ | ||||||||||||
| Hma | ES | LVTF | h | 0.001 | a | b | ab | → | h | 0.920 | <0.001 | a | b | a | → | h | 0.023 | ab | a | b | → | h | 0.106 | h | 0.096 | 0.797 | ||||||||||||||||||
| Mlu | ES | LVTF | h | 0.053 | 0.109 | h | 0.896 | h | 0.001 | a | a | b | ↘ | h | 0.002 | a | b | ab | → | 0.009 | a | b | a | → | 0.421 | |||||||||||||||||||
| Afa | LS | TMF | h | 0.361 | 0.321 | <0.001 | a | b | b | ↘ | 0.056 | h | 0.119 | 0.001 | a | b | a | → | 0.228 | |||||||||||||||||||||||||
| Cgr | LS | TMF | h | 0.820 | 0.218 | h | 0.007 | ab | a | b | → | h | 0.199 | 0.006 | a | b | b | ↘ | h | <0.001 | a | a | b | ↘ | 0.782 | |||||||||||||||||||
| Fsa | LS | TMF | <0.001 | a | a | b | ↗ | 0.002 | a | b | ab | → | h | 0.024 | a | b | ab | → | h | 0.747 | 0.254 | h | 0.012 | a | ab | b | ↘ | 0.485 | ||||||||||||||||
| Paf | LS | TMF | 0.023 | a | ab | b | ↘ | 0.572 | h | 0.203 | h | 0.882 | h | 0.056 | 0.480 | 0.195 | ||||||||||||||||||||||||||||
| Sgu | LS | TMF | h | <0.001 | a | b | c | ↗ | h | <0.001 | a | b | c | ↗ | h | <0.001 | a | b | b | ↘ | 0.039 | ab | a | b | → | h | 0.008 | a | b | ab | → | h | 0.012 | a | b | ab | → | h | 0.813 | |||||
| Cgo | LS | LVTF | 0.024 | a | b | b | ↗ | 0.017 | a | ab | b | ↗ | h | 0.434 | h | 0.451 | h | 0.038 | * | ↘ | h | <0.001 | a | a | b | ↘ | <0.001 | a | a | b | ↘ | |||||||||||||
| Eex | LS | LVTF | 0.788 | 0.313 | <0.001 | a | b | a | → | h | 0.948 | h | <0.001 | a | b | b | ↘ | <0.001 | a | b | c | ↘ | <0.001 | a | a | b | ↘ | |||||||||||||||||
| Fth | LS | LVTF | 0.547 | 0.063 | h | 0.093 | h | <0.001 | a | b | ab | → | 0.735 | h | 0.023 | ab | a | b | ↘ | h | 0.027 | ab | a | b | ↘ | |||||||||||||||||||
| p-Values for Two Years Data | p-Values for One Year Data | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Source | df | Leaf W/L | Leaf Size | LMA | LVA | LD | NM | PM | NA | PA |
| Tree height (covariate) | 1 | ns | <0.001 | <0.001 | 0.022 | ns | ns | ns | ns | ns |
| Year (repeated) | 1 | <0.001 | <0.001 | 0.039 | ||||||
| Site | 2 | <0.001 | 0.596 | 0.249 | 0.116 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Species | 17 | <0.001 | 0.007 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Year * Site | 2 | 0.108 | 0.114 | 0.007 | ||||||
| Year * Species | 17 | <0.001 | <0.001 | <0.001 | ||||||
| Year * Tree height | 1 | <0.001 | 0.011 | |||||||
| Site * Species | 34 | <0.001 | 0.033 | <0.001 | 0.217 | 0.035 | <0.001 | <0.001 | <0.001 | <0.001 |
| Site * Tree height | 2 | 0.815 | 0.717 | 0.203 | ||||||
| Species * Tree height | 17 | <0.001 | 0.028 | 0.016 | ||||||
| Year * Site * Species | 34 | <0.001 | 0.242 | <0.001 | ||||||
| Year * Site * Tree height | 2 | 0.517 | 0.313 | |||||||
| Year * Site * Tree height | 17 | <0.001 | <0.001 | |||||||
| Site * Species * Tree height | 34 | <0.001 | 0.196 | 0.702 | ||||||
| Year * Site * Species * Tree height | 34 | 0.043 | 0.000 | |||||||
| p-Values for Species | ||||
|---|---|---|---|---|
| Variable/Source | Cgr | Mki | Pfu | Sgu |
| Leaf size | ||||
| Age | 0.20 | <0.001 | 0.41 | 0.12 |
| Elevation | 0.72 | <0.001 | 0.64 | 0.54 |
| Age * Elevation | 0.53 | <0.001 | 0.14 | 0.019 |
| LMA | ||||
| Age | 0.070 | 0.005 | 0.21 | 0.83 |
| Elevation | 0.075 | 0.013 | 0.47 | 0.82 |
| Age * Elevation | 0.31 | 0.20 | 0.86 | 0.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manishimwe, A.; Ntirugulirwa, B.; Zibera, E.; Nyirambangutse, B.; Mujawamariya, M.; Dusenge, M.E.; Bizuru, E.; Nsabimana, D.; Uddling, J.; Wallin, G. Warming Responses of Leaf Morphology Are Highly Variable among Tropical Tree Species. Forests 2022, 13, 219. https://doi.org/10.3390/f13020219
Manishimwe A, Ntirugulirwa B, Zibera E, Nyirambangutse B, Mujawamariya M, Dusenge ME, Bizuru E, Nsabimana D, Uddling J, Wallin G. Warming Responses of Leaf Morphology Are Highly Variable among Tropical Tree Species. Forests. 2022; 13(2):219. https://doi.org/10.3390/f13020219
Chicago/Turabian StyleManishimwe, Aloysie, Bonaventure Ntirugulirwa, Etienne Zibera, Brigitte Nyirambangutse, Myriam Mujawamariya, Mirindi E. Dusenge, Elias Bizuru, Donat Nsabimana, Johan Uddling, and Göran Wallin. 2022. "Warming Responses of Leaf Morphology Are Highly Variable among Tropical Tree Species" Forests 13, no. 2: 219. https://doi.org/10.3390/f13020219