Abstract
Today, the use of nursery-produced seedlings is the most widely adopted method in forest restoration processes. To ensure and enhance the performance of transplanting seedlings into a specific area, soil amendments are often used due to their ability to improve soil physicochemical properties and, in turn, plant growth and development. The aim of the present study was to evaluate Populus euramericana growth and development on a growing substrate added with biochar and compost, both alone and in combination. To accomplish this aim, a pot experiment was performed to test biochar and/or compost effects on growing substrate physicochemical characteristics, plant morpho-physiological traits, and plant phenology. The results showed that biochar and/or compost improved growing substrate properties by increasing electrical conductivity, cation exchange capacity, and nutrient concentrations. On the one hand, these ameliorations accelerated poplar growth and development. On the other hand, amendments did not have positive effects on some plant morphological traits, although compost alone increased plant height, and very fine and fine root length. The combined use of biochar and compost did not show any synergistic or cumulative beneficial effects and led to a reduction in plant growth and development. In conclusion, compost alone seems to be the best solution in both ameliorating substrate characteristics and increasing plant growth, highlighting the great potential for its proper and effective application in large-scale forest restoration strategies.
1. Introduction
Forest ecosystems cover thirty-one percent of the global land area [1] and are important for human livelihoods, climate stability, and biodiversity conservation [2]. However, forests are still under constant threat because of deforestation and forest degradation, which continue at an alarming rate [3]. Since 1990, it has been estimated that 420 million hectares of forest have been lost through conversion to other land uses [4]. Between 2015 and 2020, the rate of deforestation was estimated at 10 million hectares per year, and 15 billion trees are cut down every year [5]. Currently, forests are no longer endangered solely by deforestation practices, but in an increasingly warming world, phenomena such as windstorms, heat waves, and droughts pose new and severe threats [6]. Recently, it has been documented that forest death in southwestern Australia has been directly linked to a heat wave that occurs during a “warmer drought” [7].
At the international level, the sustainable management of reforested and afforested sites has been promoted to contribute to maintaining forest status, preserving biodiversity [8], guaranteeing ecosystem services, reducing climate change impact [9], and avoiding soil degradation and desertification [10]. Today, many countries are increasing reforestation and afforestation efforts to remediate the many practices of forest clearance that occurred in the past and forest losses caused by harmful events related to climate change [11].
However, both reforestation and afforestation processes are complex, time-demanding [12], and expensive considering the costs of soil preparation and fertilization, plant tree species selection and acquirement, maintenance, and management practices [13]. Moreover, forest restoration projects have high failure rates due to the high mortality of plants before reaching maturity [14]. For these reasons, it has become increasingly important to develop methods that may increase seedling survival, growth, and vigor while reducing labor costs [15]. In particular, it is important to ensure that planted seedlings have high survival rates and good growth [16], and the use of nursery-produced seedlings is the most widely adopted method in forest restoration processes [17]. Successful plant establishment depends greatly on decisions made prior to planting, which must take into account species and site characteristics [18] and that nursery-produced seedlings will face a variety of stress factors after nursery production [19].
Several methods have been tested in order to ensure and increase the performance of seedling outplanting in deforested and degraded lands, in which salinity, low water holding capacity, and lack of nutrient availability affect the realization of reforestation and afforestation methods [12,20,21]. To overcome these limitations, soil amendments can be used to improve the physicochemical properties of soils [22] and guarantee land restoration success [23].
Among the different amendments, biochar—a charcoal produced by the pyrolysis of organic waste feedstocks, such as manure and crop residues—has received attention [24]. It is a carbon sequestration agent able to reduce atmospheric CO2 concentrations and is an excellent soil conditioner [25]. Indeed, it is effective in improving degraded sites because biochar application increases soil aggregate stability and water holding capacity by enhancing soil pore characteristics and water retention [26]. In addition to soil responses, biochar may yield a wide range of benefits for plant germination, growth, productivity, and survival, and stress management [20]. Several mechanisms may enhance plant development in response to soil biochar additions, including: (i) reduction of nutrient leaching and improvement of plant-available nutrients; (ii) release of carbon, nitrogen, phosphorus, and potassium; and (iii) increase of soil biota density and diversity [27]. Thus, it is evident that biochar has several properties of particular interest from the perspective of forest restoration. First, its recalcitrance implies that biochar added in the context of a restoration project will not rapidly decompose [28]. Secondly, biochar positively acts on plant growth and survival in highly degraded soils, which are frequent in the context of forest restoration [29]. It is also particularly effective in adsorbing a wide range of materials that are either generally toxic or adverse to plant growth at high concentrations, including metals, salts, polyaromatic hydrocarbons, and residual herbicides [30]. Lastly, biochar may be relatively easily and economically generated from locally available feedstocks, and thus it offers important potential advantages in both economic and logistic terms [15].
Another soil conditioner that may be profitably used in forest restoration is compost, a fertilizer able to improve soil quality by: (i) incorporating organic matter [31,32], nutrients, and electrolytes into soil [33]; and (ii) enhancing soil structure, density, and porosity [34], which increase water retention capacity and reduce soil erosion and nutrient leaching [35]. In turn, such compost properties result in the compost having positive effects on plant growth, even because compost amendment may activate a wide range of natural disease suppressiveness mechanisms against plant pathogens [36]. Moreover, by enhancing the carbon storage capacity in the soil, compost might be used in reducing global warming [37].
Numerous studies have suggested the application of biochar in combination with compost as a promising strategy to promote plant growth and performance, having positive synergistic effects on soil properties [23,38,39,40]. However, this positive effect is strictly related to specific soil characteristics, plant species, amendment application rates, and feedstock [20]. Thus, more experiments are needed to accurately test synergism using quantifiable metrics to determine what a “target” seedling might be [41].
The Target Plant Concept (TPC) is an effective framework for defining, producing, and managing plant material (e.g., seeds, cuttings, and seedlings) based on characteristics appropriate for a specific site [16]. These characteristics are often scientifically derived from test factors that are linked to outplanting performance, such as seedling morphology and physiology [42], genetic origin, and the ability to overcome limiting conditions on outplanting sites [43]. Commonly measured morphological attributes include bud development, dry weight fraction, stem height and diameter, and root development, which are used in seedling quality assessment programs to monitor plant growth and survival [10]. Seedling morphological attributes cannot be used alone to assess seedling quality because morphology does not describe physiological vigor [44], and, as mentioned before, seedling morphology is combined with the assessment of physiological attributes (e.g., nutrient status, root growth potential, stress tolerance) to relate seedling quality at lifting to field performance after planting [41].
In line with the above, our study examined the potential for biochar and compost amendments, alone or in combination, to have benefits in nursery-produced seedling systems, concomitantly enhancing a range of soil properties, and improving plant growth and development. The main goal of this study was to conduct targeted research on poplar plants to highlight the potential beneficial use of biochar and/or compost in reforested environments. Poplar and its hybrids can be used to create economic benefits and improve environmental quality in forestry and agroforestry worldwide [45,46]. They have shown the capacity for rapid biomass accretion [47] and are currently assuming growing importance for timber and bioenergy production [48]. The adoption of poplar species into the agroforestry system has the added benefit of sequestering carbon emitted from agricultural practices [49]. Thus, the outcomes of this research might enlarge the understanding of soil amendments as ameliorants of soil properties, as well as survival of key species in reforestation and afforestation programs [50].
To accomplish the objective of the present research, biochar and compost, both alone and in combination, were tested on hybrid poplar seedlings with the aim of determining if the addition of biochar and/or compost to a growing media may (i) enhance soil physicochemical properties and (ii) improve plat morphophysiological traits, with particular regard to root morphology and development.
2. Materials and Methods
2.1. Experimental Design and Growth Characteristics
Two-year-old woody cuttings of the hybrid poplar clone I-214, Populus deltoides × Populus nigra (Populus euramericana (Dode) Guinier) were rooted in 3 L pots containing vermiculite and kept in a growth chamber under controlled conditions (25 °C air temperature, 50–70% relative humidity, 15-h photoperiod) for 60 days.
Then, 40 homogeneously rooted cuttings, with similar morphological traits (e.g., size), were selected for the experiment and transplanted, separately, into 20 L plastic pots. These pots were filled with four different growing medium combinations (hereafter also called growing substrates or treatments), and ten replicates were set up for each of them. P. euramericana plants were grown for each treatment in a greenhouse for 12 months. The experimentation began in July and ended in July of the following year. In the 12 months of experimentation, hereinafter, are indicated by the acronyms ranging from T0 to T12 (Figure 1). Growth occurred under a controlled water regime and natural photoperiod and temperature (for minimum, maximum, and average temperature values see Table S1 in Supplementary Material), and the plants were arranged in a randomized complete block design and rotated to a different position within the block throughout the trial. The pots were fully irrigated to prevent water stress (twice a day, as required), and a suspended net was used to reduce exposure to sunlight.
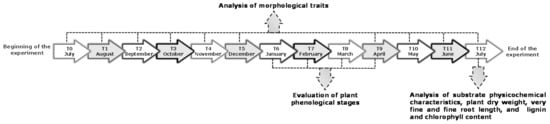
Figure 1.
Experimental schedule with the main analyses and the sampling times (months).
The four treatments were: (i) control mix (Ctrl), composed of soil and sand (1:1, v:v); (ii) biochar mix (B), composed of soil and sand (1:1) plus biochar; (iii) compost mix (Co), composed of soil and sand (1:1) plus compost; and (iv) biochar and compost mix (BCo), composed of soil and sand (1:1) plus biochar and compost. For treated pots, biochar and compost were added to the soil and sand mixture at a concentration of 25 g·kg−1 of dry substrate (application rate of 2.5% w/w).
2.2. Soil, Biochar, and Compost Characteristics
The soil was collected from an uncultivated pasture area, located in Pesche (Molise, Italy), with a floral composition predominantly of graminoid grasses, not under a rotation system, and that includes hedges. This area is mainly used for grazing, but the fodder is harvested mechanically. The soil was loamy mixed mesic soil, according to the United States Department of Agriculture (USDA) classification [51]. As reported previously, the soil was mixed with commercial sand, and this mixture was characterized to determine the pre-planting physicochemical properties. The soil–sand mix was moderately subalkaline with a silt loam texture according to the USDA classification. Moreover, it was characterized by low electrical conductivity (EC), cation exchange capacity (CEC), and nitrogen and carbon content. For the experiment, the mixture of the soil and sand was air-dried for 72 h, weighed, finely crushed, and then mixed thoroughly before packing lightly in the pots on top of the pebbles placed on the base to improve drainage.
The biochar used was a commercial charcoal (provided by Romagna Carbone s.n.c., Bagnacavallo, Italy), obtained from orchard pruning biomass through a slow pyrolysis process with an average residence time of 3 h, at a temperature of 500 °C, in a kiln of 2.2 m in diameter, and holding around 2 ton of feedstock.
The compost was a commercial product (composted olive mill residues) prepared in a standardized experimental composting process reported by Alfano et al. [52]. Briefly, compost was prepared by mixing humid olive husks from a two-phase extraction plant with olive leaves (8% w/w); one-year-old, humid, and composted husks (25% w/w) were then added to this mixture. A complete overview of both biochar and compost characteristics is reported by Trupiano et al. [53].
2.3. Growing Substrate Analysis
At the end of the experiment (T12), after 12 months of plant growth, soil samples were collected (Figure 1) and air dried at 20–25 °C for 72 h. The moisture content was calculated according to the Black method [54] as the difference in sample weight before and after oven drying to a constant weight at 105 °C. The pH was measured in H2O and 0.01 M CaCl2 using a pH meter (Eutech Instruments, Thermo Fisher Scientific, Waltham, MA, USA) and a 1:2.5 soil weight:extract-volume ratio. The alkalinity of samples with a pH value greater than 7.0 was determined by titrimetry according to Rayment and Higginson [55]. Electrical conductivity (EC) was determined by a conductivity meter (Cond 510, XS Instruments, Carpi, Italy) on a 1:5 soil:water suspension [56]. Cation exchange capacity (CEC) was assessed according to the method of Mehlich [57] using BaCl2. Total organic carbon (TOC) and total nitrogen (Ntot) contents were determined by dry combustion [58] using a CHN elemental analyzer (Mod 1500, series 2, Carlo Erba Instruments, Cornaredo, Italy). In the case of TOC, combustion was carried out after the complete removal of inorganic carbon with acid. Total phosphorus (Ptot) was detected by spectrophotometry (UV-1601 Shimadzu) according to the test method described by Bowman [59], while available phosphorus (Pav) was extracted by a NaHCO3 solution at pH 8.5 and evaluated by spectrophotometry according to the Olsen test method [60]. Particle size distribution (also named soil texture) was quantified by hydrometer analysis through a modification of the Bouyoucos method [61] (according to Beretta et al. [62]) on samples previously dry-sieved at 2 mm. The fraction with a diameter smaller than 2 mm was treated with H2O2 and wet sieved at 200 µm, 50 µm, and 20 µm. Measurements of density were carried out by a hydrometer on samples smaller than 20 µm previously dispersed with sodium hexametaphosphate solution. Moreover, in order to quantify the large fraction of macro aggregates, particles that did not pass through the 2 mm sieve were treated with sodium hexametaphosphate solution to disrupt aggregates, and subsequently, the difference in weight before and after wet sieving at 2 mm was measured [63].
2.4. Plant Analysis
2.4.1. Morphological Traits
From the beginning (T0) to the end of the experiment (T12), plant growth was monitored monthly by measuring morphological traits. More detailed, morphological traits were assessed at time points T1–T5, corresponding to the months of August–December, and at time points T9–T12, corresponding to the months of April–July. The T1–T5 time points were equivalent to the months preceding the dormancy phase (January—T6, February—T7, and March—T8) and T9–T12 to the months of the vegetative phase of P. euramericana (Figure 1). For morphological traits, the variations in plant height and leaf number (Δ) were determined. Additionally, the main leaf parameters were assessed: leaf area, perimeter, length, and width. Image J 1.8.0 software (Wayne Rasbanb National Institute of Health, Bethesda, MD, USA; http://rsb.info.nih.gov/ij/ (accessed on 20 November 2020)) was used for these analyses. The aboveground (leaves and stems) and belowground (roots) dry weight, at time T12, were also determined after two days of drying in an oven at 80 °C, and stem/root ratio (S/R ratio) was calculated in terms of dry weight. Furthermore, roots were analyzed using WinRhizo Pro V. 2007d (Regent Instruments Inc. Quebec, QC, Canada). In particular, roots were divided into two groups on the basis of their diameter to distinguish very fine roots (diameter measurement between 0 and 0.5 mm) and fine roots (diameter measurement between 0.5 and 2 mm) [21,64].
2.4.2. Phenology
Phenological stage assessment was performed on poplar plants evaluating apical bud development in the three months of dormancy (January—T6, February—T7, and March—T8) and in the first month of active vegetative growth (April—T9) (Figure 1). Bud development was quantified using six levels of morphology scores (0–6) according to Trupiano et al. [65]. Briefly, a minimal score (0) was given to the winter bud and a maximum score (6) to the flushing buds, with a growing stem.
2.4.3. Lignin and Chlorophyll Content
At the end of the experiment (T12), lignin and chlorophyll contents were also assessed (Figure 1).
Lignin content was measured using the protocol of Doster and Bostock [66] with a few modifications, as detailed in Trupiano et al. [67]. Briefly, lignin amount within each sample was calculated by measuring the absorbance at 280 nm, using a specific absorbance coefficient of 6.0 L·g−1 cm. Since this specific absorbance coefficient provides only an approximate conversion [66], the samples with the highest lignin content value were used as a standard in the relative measurements of lignin concentration of the other samples.
Chlorophyll content was measured in three randomly sampled leaf discs (10 mm in diameter). Extraction was performed with N, N dimethylformamide (DMF) for 48 h at 4 °C, in the dark at a ratio of 1:20 (plant material:solvent, w:v). The extinction coefficients proposed by Inskeep and Bloom [68] were used for quantification by spectrophotometric analysis. The following equations were used: Chl a = 12.70A664.5 − 2.79A647; Chl b = 20.70A647 − 4.62A664.5; total Chl = 17.90A647 + 8.08A664.5 where A is absorbance in 1.00 cm cuvettes.
2.5. Statistical Analysis
To evaluate the effect of each treatment on the conducted tests, all statistical analyses were performed with the R software version 3.4.3 (R Development Core Team 2017). The normality and homoscedasticity of the data were assessed with the Shapiro and Bartlett tests, respectively. The mean values were compared using the parametric analysis of variance test (ANOVA) for normal data or the non-parametric Kruskal test for non-normal data. Following this, a post hoc test (Tukey HSD or pairwise Wilcox tests, respectively) was performed.
3. Results
3.1. Growing Substrate Characteristics
At the end of the experiment (T12), the moisture content of the medium with the compost addition (Co substrate) (1.15 g·kg−1) was 32% lower compared to the control (1.52 g·kg−1). However, no significant difference was present among the four different substrates in pH and alkalinity values (Table 1).

Table 1.
Main physicochemical characteristics determined after 12 months of P. euramericana grown on four different substrates (Ctrl, B, Co, and BCo). Different letters indicate significant difference (p < 0.05) (n = 3 ± SE). Ctrl: control mix of soil and sand; B: mix of soil and sand + biochar; Co: mix of soil and sand + compost; BCo: mix of soil and sand + biochar + compost.
Electrical conductivity (EC) was increased (2-fold) in the Co treatment (0.447 dS·m−1) compared with the Ctrl growth medium (0.233 dS·m−1). Cation exchange capacity (CEC) was increased in all three treatments with biochar and/or compost addition (B, Co, and BCo) compared to Ctrl (6.49 cmol(+)·kg−1). In detail, CEC was increased by 33%, 49%, and 48% in the B, Co, and BCo treatments compared to the Ctrl medium (Table 1). The contents of total organic carbon (TOC), total nitrogen (Ntot), and total (Ptot) and available phosphorus (Pav) varied significantly and differently among the four treatments. However, the values of these nutrients were all higher in the Co and BCo treatments. Specifically, TOC was 2-fold higher in both the Co and BCo substrates compared to both other two growing media (Ctrl and B). With regard to Ntot, it was increased 2-fold in the B substrate (0.52 g·kg−1), 3-fold in the Co medium (1.60 g·kg−1), and 9-fold in the BCo mix (2.06 g·kg−1) with respect to the control (0.23 g·kg−1). Ptot was also raised by 66% in the B treatment (215 mg·kg−1) and was increased 2-fold in the Co (301 mg·kg−1) and BCo treatments (324 mg·kg−1) compared with the control (129.33 mg·kg−1). The highest values of Pav were measured in the two Co (16.90 mg·kg−1) and BCo treatments (18.63 mg·kg−1) compared to the control growth medium and the B treatment in which there was an amount of phosphorus less than 12 mg·kg−1. Particle size analysis showed that there were differences among the treatments only for the clay and fine sand fractions (Table 1). Soil with the compost (Co substrate) had the highest clay content (13.5%), and in detail, the clay fraction was 52% higher than the control (8.9%). In contrast, the biochar decreased the clay component of the B substrate (6.27%) by 30% compared to the Ctrl. Regarding the fine sand content, the analyses showed only a difference between the sand fraction found in the B treatment (9.5%) and that measured in the BCo mix (6.73%).
3.2. Plant Characteristics
3.2.1. Morphological Traits
After one month of poplar plant growth (T1), the combined addition of the biochar and compost to the growth substrate (BCo) appeared to negatively influence the increase in stem height of Populus euramericana seedlings (Figure 2). In the control growing medium (Ctrl), the stem height increased (50%) with respect to the BCo substrate in which the percentage of stem height increased only 27% (Figure 2). At time T2, despite the plant grown on the B, Co, and BCo treatments increasing in stem height of 31%, 7%, and 17%, respectively, the highest increase for this parameter was observed in the Ctrl plants (67%) (Figure 2). From October to March (T3 to T8), as the winter dormancy period advanced, no changes in stem height measurements were recorded in the four different growing media. At times T9 (April) and T10 (May), the development and growth phase restart of P. euramericana plants resulted in a significantly higher increases in stem height in both treatments with the compost (Co) and biochar (B) added alone compared to the Ctrl and BCo substrates. Indeed, at time T9, the percentage increase in stem height was 9% in the Co substrate and 5% in the B treatment, while it was 1% in Ctrl and 2% in the BCo mix (Figure 2). At time T10, the increases in stem height were 7% in Co and 6% in B, while stem height increased by 5% and 2% in the Ctrl and BCo substrates, respectively (Figure 2). At time T11, the addition of Co to the soil resulted in the highest percentage of stem growth (59%) compared to the Ctrl (12%), B (14%) and the BCo mix (21%) (Figure 2).
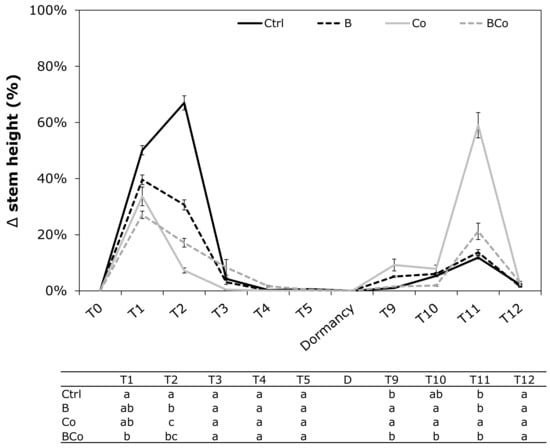
Figure 2.
Variation (Δ) in stem height (%) determined monthly during the 12 months of P. euramericana growth on four different substrates (Ctrl, B, Co, and BCo). Different letters indicate significant differences among the treatments for each sampling time (p < 0.05) (n = 10 ± SE). Ctrl: control mix of soil and sand; B: mix of soil and sand + biochar; Co: mix of soil and sand + compost; BCo: mix of soil and sand + biochar + compost. T0 to T12: Sampling times (months).
The leaf number changed in poplar seedlings throughout the experiment, and for the four different treatments, it showed highly variable and different trends in each month (Figure 3). The greatest changes between the different treatments of the experiment were observed at T9 and T10. At time T9, the highest increase in leaf number was measured in plants grown on B substrate (559%), followed by 261% and 309% percentage increases observed in the Ctrl and Co growth media, respectively; in the BCo mix, conversely, a strong decrease in leaf number was reported (−92%) (Figure 3). At time T10, the situation changed: the combined addition of biochar and compost (BCo) resulted in the highest percentage of leaf number increase (611%) compared to the Ctrl, B, and Co substrates, in which the percentages were 43%, 23%, and 8%, respectively.
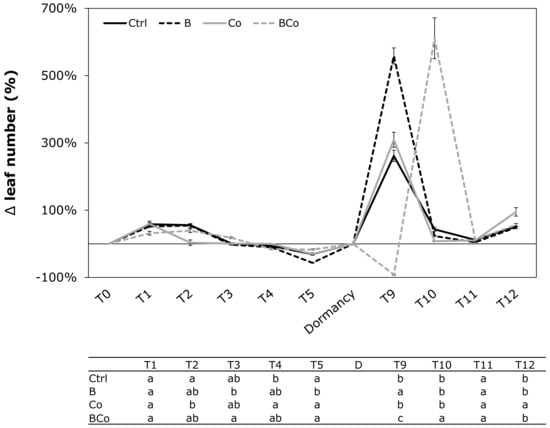
Figure 3.
Variation (Δ) in leaf number (%) determined monthly during the 12 months of P. euramericana growth on four different substrates (Ctrl, B, Co, and BCo). Different letters indicate significant differences among the treatments for each sampling time (p < 0.05) (n = 10 ± SE). Ctrl: control mix of soil and sand; B: mix of soil and sand + biochar; Co: mix of soil and sand + compost; BCo: mix of soil and sand + biochar + compost. T0 to T12: Sampling times (months).
The main leaf parameters (leaf area, perimeter, length, and width) reported significant differences (Figure 4) in the month immediately preceding the winter dormancy phase (December—T5) and in the two months immediately following dormancy and in which there was active vegetative growth of P. euramericana plants (April—T9 and May—T10). In detail, at time T5, all four leaf parameters were lower in B, C, and BCo than in the Ctrl. After the dormancy phase (times T9 and T10), leaf parameters were unchanged in B and Co, whereas they were lower in BCo compared to the Ctrl (Figure 4).
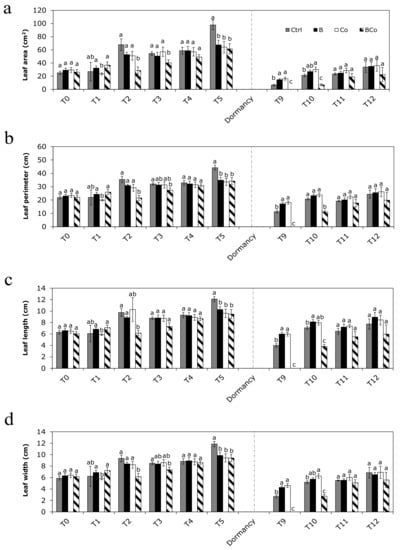
Figure 4.
Leaf area (a), perimeter (b), length (c), and width (d) determined monthly during the 12 months of P. euramericana growth on four different substrates (Ctrl, B, Co, and BCo). Different letters indicate significant differences among the treatments for each sampling time (p < 0.05) (n = 10 ± SE). Ctrl: control mix of soil and sand; B: mix of soil and sand + biochar; Co: mix of soil and sand + compost; BCo: mix of soil and sand + biochar + compost. T0 to T12: Sampling times (months).
Indeed, more specifically, at time T5, the leaf area in the Ctrl was 98 cm2, while it was almost twice as small in the B (68 cm2), Co (64 cm2), and BCo substrate (62 cm2) (Figure 4a). At time T9, as also shown in Figure 3, poplar plants grown in the BCo treatment were without leaves, and therefore, leaf area was assigned a value of zero (Figure 4a). Conversely, in the B and Co substrates, they were 15 cm2 and 16 cm2, respectively, and thus 3 times higher than that measured for the leaves of the Ctrl plants (6 cm2) (Figure 4a). At time T10, plants grown in the Ctrl, B, and Co growing media had the highest value of leaf area (30 cm2), which was 7 times bigger than the leaf area of plants grown in the BCo mix (6 cm2) (Figure 4a).
Regarding the leaf perimeter (Figure 4b), at time T5, the lowest values were found in the B, Co, and BCo treatments (approximately 34 cm compared to the 44 cm measured in the Ctrl plants). At time T9, it was one and a half times bigger in the B (17 cm) and Co (18 cm) treatments with respect to the Ctrl (11 cm), whereas, as mentioned above, plants grown in the BCo mix had no leaves. At time T10, plants grown on the Ctrl, B, and Co substrate had leaf perimeter values of 21 cm, 23 cm, and 24 cm, respectively, which were twice as high as that of plants grown on the BCo mix (11 cm).
The leaf length value was also lower at time T5 for B (10 cm), Co (10 cm), and BCo (9 cm) plants compared to the Ctrl poplar seedlings (12 cm) (Figure 4c). At time T9, it was two-fold higher in B and Co (6 cm in both treatments) compared to the Ctrl and BCo (Figure 4c), whereas at time T10, it was twice as low only in plants grown on the BCo mix (4 cm).
The leaf width (Figure 4d), at time T5, was slightly lower in the B (10 cm), Co (9 cm), and BCo (9 cm) plants than in the Ctrl (12 cm), whereas at time T9, it was approximately twice as large in the B (4 cm) and Co (5 cm) plants compared to the Ctrl plants (Figure 4d). This parameter resulted in the lowest BCo at time T9 and T10 (Figure 4d).
The addition of biochar and compost alone (B and Co treatments) had no effect on the leaf dry weight (Figure 5a). In contrast, the BCo mix decreased the leaf dry weight of P. euramericana plants by 40% (9.2 g) compared to the Ctrl growth medium (15.2 g) (Figure 5a). The three treatments—B, Co, and BCo substrates—decreased the stem dry weight by 13%, 12%, and 10%, respectively, with respect to the dry weight measured in the Ctrl (87.8 g) (Figure 5a). However, for the root dry weight, the combination of the two amendments (BCo) resulted in a decrease in dry weight of 47% (11.3 g) compared to the Ctrl (21.5 g) and 45% with respect to the Co treatment (20.4 g) (Figure 5a). The highest stem/root ratio (S/R ratio) was found for plants grown in the BCo mix; these plants had a S/R ratio of 7.42 that was almost two times higher than that found in the Ctrl (4.12) (Figure 5b). For plants grown in the B and Co substrates, the S/R ratio was not significantly different from the Ctrl, although it showed an increased trend in the case of B (5.86) (Figure 5b).
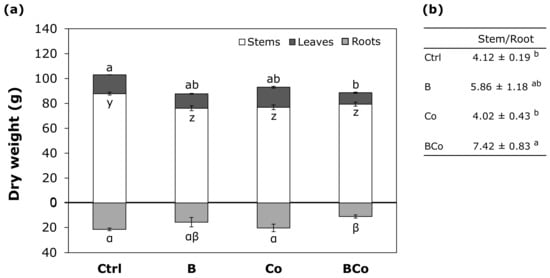
Figure 5.
(a) Dry weight (g) of the different organs (black box, leaves; white box, stems; gray box, roots) of P. euramericana grown on four different substrates (Ctrl, B, Co, and BCo). Different letters indicate a significant difference among substrates (p < 0.05) (n = 3 ± SE): letters a, b for leaves; letters y, z for stems; letters α, β for roots. (b) Stem/root ratio calculated as dry weight for stems/dry weight for roots. Different letters indicate a significant difference among substrates (p < 0.05) (n = 3 ± SE). Ctrl: control mix of soil and sand; B: mix of soil and sand + biochar; Co: mix of soil and sand + compost; BCo: mix of soil and sand + biochar + compost.
The addition of compost to the soil (Co treatment) resulted in the highest increase in the length of both very fine and fine roots (Figure 6). More specifically, the Co treatment led to a 33% and 16% increase in very fine root length (12,160 cm) and fine root length (2932 cm) compared to the control growing medium (9115 cm and 2517 cm, respectively; Figure 6). On the other hand, there was a 34% decrease in the BCo plants with respect to the Ctrl substrate (9115 cm) in the case of a very fine root length (Figure 6). Moreover, the two biochar treatments (B and BCo substrates) resulted in a decrease in fine root length of 27% (1837 cm) and 34% (1660 cm) with respect to the Ctrl (2517 cm), respectively (Figure 6).
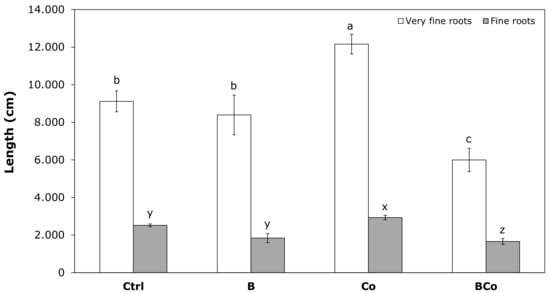
Figure 6.
Length (cm) of very fine roots (0 < d < 0.5 mm) and fine roots (0.5 < d < 2 mm) (white box, very fine roots; gray box, fine roots) determined after 12 months of P. euramericana growth on four different substrates (Ctrl, B, Co, and BCo). Different letters indicate a significant difference among substrates (p < 0.05) (n = 3 ± SE). Letters a, b, c regard very fine roots; letters x, y, z regard fine roots. Ctrl: control mix of soil and sand; B: mix of soil and sand + biochar; Co: mix of soil and sand + compost; BCo: mix of soil and sand + biochar + compost.
3.2.2. Phenology
The evaluation of P. euramericana plant phenological stages (Figure 7) showed that, during winter dormancy, at time T6 (January), all seedlings (100%) in all substrates (Ctrl, B, Co, and BCo) had buds with a morphological score of 0. Successively, at time T7 (February), only seedlings grown in the Ctrl growing medium had buds with a morphological score of 0 (100%). The biochar use (B growth medium) resulted in 20% of the poplar plants having buds with a morphological score of 1. In the two treatments with the compost addition (Co and BCo), the seedlings had buds not only in stage 1 but also in stage 2. In detail, 30% and 10% of plants grown in the Co treatment had buds in stages 1 and 2, respectively. In the BCo mix, 20% and 10% of P. euramericana seedlings had buds that were given morphological scores of 1 and 2, respectively.
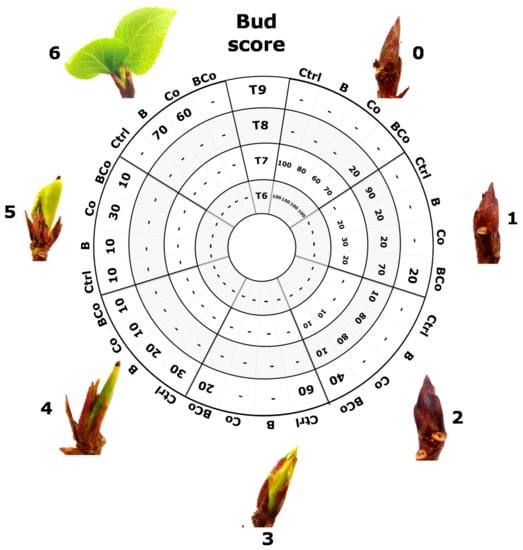
Figure 7.
Evaluation of phenological stages of P. euramericana grown on four different substrates (Ctrl, B, Co, and BCo) by bud development analysis at different time points: winter dormancy condition (T6, January and T7, February), release from dormancy (T8, March), and active vegetative phase (T9, April). For each time point, the percentage of plants characterized by a specific stage of bud development is reported. Bud development was quantified using six levels of morphology score (0–6); minimal score (0) was given to the winter bud, and maximum score (6) to buds with a growing stem. Ctrl: control mix of soil and sand; B: mix of soil and sand + biochar; Co: mix of soil and sand + compost; BCo: mix of soil and sand + biochar + compost.
At time T8 (release from dormancy condition), almost all plants in the control substrate showed buds at stage 1 with only 10% of P. euramericana plants at stage 2. Whereas both treatments with the addition of either biochar alone (B substrate) or compost alone (Co substrate) resulted in 20% of plants having stage 1 and those of the remaining 80% having a morphological score of 2. Instead, the combined biochar and compost addition to the soil (BCo mix) resulted in the poplar plants still having stage 0 buds, at time T8. In detail, 20% of the seedlings had buds with the minimum score (0), 70% of P. euramericana plants had buds at stage 1, and the remaining 10% showed buds with a morphological score of 2.
During the active vegetative phase (time T9, April), bud development differed among the four distinct treatments. Indeed, in the control growing medium (Ctrl), 60% of the plants had buds at stage 3, and 30% and 10% of the poplar seedlings had buds with morphological scores of 4 and 5, respectively. At time T9, the addition of the biochar alone (B substrate) or compost alone (Co substrate) to the growing substrate resulted in buds characterized by development stages to which scores of 4, 5, and 6 were assigned. In detail, 20% of P. euramericana grown in the B substrate had buds with a score of 4, 10% showed buds with a score of 5, and the majority of plants (70%) had buds that were given the maximum score (6). For the Co treatment, 10% of the seedlings were characterized by buds in stage 4, 30% of poplar plants had buds in stage 5, and 60% showed stage 6 buds. Plants grown in the BCo mix had buds with morphological scores ranging from 1 to 5. Specifically, 20% of the plants contained buds with a score of 1, 40% were characterized by buds in stage 2, 20% showed buds with a morphological score of 3, and the remaining plants were half (10%) characterized by buds in stage 4 and half (10%) by buds in stage 5.
3.2.3. Lignin and Chlorophyll Content
The two treatments with the biochar, both added alone (B substrate) and in combination with the compost (BCo substrate), showed the highest lignin content values in the P. euramericana roots (Figure 8). In the B treatment, the lignin content was 100%, while in the BCo mix, it was 87%. Thus, in the two biochar treatments, the lignin content was higher than the value found for plants grown in the Ctrl growing medium (70%) and 2 times higher with respect to the content measured for plants developed in the Co substrate (41%).
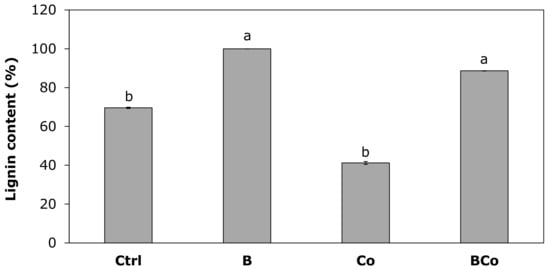
Figure 8.
Lignin content (%) determined after 12 months of P. euramericana growth on four different substrates (Ctrl, B, Co, and BCo). Lignin content is expressed as a percentage of the value measured in the roots of plants grown in the B substrate (considered 100%). Different letters indicate significant difference (p < 0.05) (n = 3 ± SE). Ctrl: control mix of soil and sand; B: mix of soil and sand + biochar; Co: mix of soil and sand + compost; BCo: mix of soil and sand + biochar + compost.
Regarding the chlorophyll content, Table 2 shows that the values of the total chlorophyll content (Chl), chlorophyll a content (Chl a), and chlorophyll b content (Chl b) were higher in the B and BCo substrates than in the Ctrl and Co substrates. In detail, the total chlorophyll content in both B (11.51 mg·cm−2) and BCo (11.58 mg·cm−2) was 46% higher than that measured for plants grown in the Ctrl growing medium (7.91 mg·cm−2) and two times higher than the total content measured in the Co treatment (5.47 mg·cm−2). The same trend, as mentioned above, was also observed for Chl a and Chl b content; however, there were no significant differences between the four different treatments in the ratio of Chl a to Chl b content.
Table 2.
Chlorophyll content (mg·cm−2) of P. euramericana grown on four different substrates (Ctrl, B, Co, and BCo). Different letters indicate significant difference (p < 0.05) (n = 10 ± SE).
4. Discussion
In forest restoration processes, soil amendments (e.g., biochar and compost) may help to ensure and enhance the performance of nursery-produced seedling transplants, increasing plant survival rates and growth [16]. This soil conditioner ability is not only associated with amendment characteristics (e.g., application rate and feedstock), but it is also strictly related and influenced by soil physicochemical properties and the choice of plant species [20].
Several studies have established that pyrolysis temperature and type (fast or slow), together with feedstock choice, influence final biochar physicochemical characteristics and consequently its properties as a soil conditioner [70,71,72,73]. Pyrolysis temperature affects biochar longevity, with pyrolysis temperatures higher than 500 °C generally leading to longer-term half-lives [70]. Greater pyrolysis temperatures also led to biochar containing greater carbon concentrations and a specific surface area [71]. However, it appears that feedstock selection has the largest influence on biochar properties [72]. Wood-based biochars have the greatest specific surface area, while crop- and other grass-based biochars appear to have cation exchange capacities greater than other biochars, which could potentially lead to longer-term changes in soil nutrient retention [73].
The selection and combination of feedstock material is also important for the quality of compost, which is produced primarily from animal and agricultural waste. In general, animal wastes, such as cow dung, poultry litter, pig manure, and chicken manure, contain low levels of carbon but high levels of N [74]. Most agricultural wastes, such as crop residues, sawdust, and rice straw, contain large amounts of carbon but low amounts of nitrogen and cause a slower decomposition process [75]. It is important to combine the right materials to achieve compost characterized by correct proportions of C and N. The ideal C/N ratio for compost should be kept at 25–50 because if it is lower, ammonia is emitted, and biological activity can be hindered. On the other hand, at higher ratios, N can be a limiting nutrient, and the composting process will be slow, and too much nitrogen will cause the material to become acidic and smelly [76].
Thus, more experiments are needed to accurately test the effect (negative, positive, synergistic/antagonistic) of soil amendments on selected forestry seedlings, such as poplar plants. In our study, the potential use of biochar and compost, both alone and in combination, on nursery-produced seedlings was evaluated, taking into account changes in growing substrate properties and Populus euramericana morphophysiological traits and phenology.
The biochar used was a commercial charcoal obtained from orchard pruning biomass through a slow pyrolysis process at a temperature of 500 °C. This biochar was characterized by an alkaline pH and a high C/N ratio (125.5). The compost used was prepared from olive waste and had a pH close to neutrality (7.5) and an appropriate C/N ratio (28.1).
The study showed that the compost amendment alone produced some benefits in P. euramericana growth performance compared to the biochar that, both alone and in combination with the compost, seemed to have neutral or antagonistic effects on plant morphological and physiological traits, although with some differences during the diverse plant growth phases.
In detail, during the first months of plant growth (T0–T5), the biochar or compost addition alone had a negligible or negative effect on P. euramericana growth and development. Conversely, after the winter dormancy phase (T9–T12), the situation was quite different, and their addition led to an increase in plant height and in leaf number reaching the maximum in the compost alone amended substrate. The study by Jarvis et al. [77] also showed that the addition of compost soil conditioner significantly increased the height growth of alder, birch, and aspen species. Similar to the results of the present study, Heiskanen et al. [78] found that compost increased the growth of pine, willow, red clover, red fescue, and maiden pink seedlings. This positive effect was also confirmed by phenological analysis, which showed that the two amendments alone accelerated poplar growth and development compared to the control, leading to 60% more seedlings having buds with fully formed leaves after dormancy breaking. Nevertheless, the compost and biochar mixture induced slowing in bud development and overall plant growth.
The time effect could have been due to the fact that P. euramericana plants were potted as cuttings, and it was possible that nutrients in initial cuttings were translocated to offset nutrient demand. Successively, the ability of these two amendments to ameliorate substrate characteristics could have increased the available nutrient amounts for plants and, consequently, accelerated plant growth and development [77,78]. Indeed, despite the addition of biochar or compost alone having no effect on soil pH [20,79], they increased soil electrical conductivity and cation exchange capacity; moreover, both amendments ameliorated substrate chemical characteristics by increasing nutrient concentrations (TOC and Ntot, and Ptot and Pav), probably due to their content of organic matter and inorganic ions (e.g., N and P) [80]. Furthermore, the biochar amendment could have facilitated the biochemical cycling of nitrogen and phosphorus [27], and the compost could have improved soil aeration and water availability [78].
If growth responses to the biochar and compost alone addition were mainly due to nutrient provision, their combined use negatively affected the growth of all plant aboveground parts, despite enhanced soil nutrient availability. Seehausen et al. [81] also found that the combined addition of biochar and compost in nutrient-limited media had neutral or antagonistic interactive effects on several plant growth traits and physiological performance. More specifically, the authors found mostly neutral effects on Salix purpurea plants and antagonistic effects on maximum leaf area, aboveground, and belowground biomass, reproductive allocation, maximum plant height, chlorophyll fluorescence, and stomatal conductance of Abutilon theophrasti plants. This could be related to a saturation of plant nutrient demands resulting in a non-additive positive effect [82] and might have brought to a decline in poplar growth [83]. The biochar and compost combination might also have resulted in an oversupply of toxic elements (e.g., Al, As, Pb) and micronutrients (e.g., B, Mn) [81]. Alternatively, the poplar growth decline may also be due to the fact that biochar can immobilize/retain nutrients [23] thus reacting with them and acting as a competitor instead of providing nutrients to plants. For example, biochar could facilitate phosphate precipitation/absorption reactions and lead to reduced P and N availability to plants [84,85], as observed in the results reported above. Indeed, biochar has a high cation exchange capacity, which may significantly increase nutrient retention because of a high surface charge [78].
In the present study, the soil–sand mix used as the growing plant substrate was already moderately subalkaline, and an improvement in pH after biochar application was not expected, as also reported in other studies [86,87]. It is well documented that biochar application impacts soil pH, which, together with cation exchange capacity, influences nutrient interactions in soil. Given that, in the present study, biochar application did not affect substrate pH, immobilization, and retainment of nutrients were likely to be attributed to the increases in cation exchange capacity. Similarly, Liu et al. [88] documented that adsorption of NH4+-N may be due to the high cation exchange capacity of biochar, and adsorption of ammonium by biochar has also been well documented by Spokas et al. [89]. Nguyen et al. [90] demonstrated, in a meta-analysis, that biochar addition reduced soil inorganic nitrogen by about 11% (NH4+) and 10% (NO3−) per 56 works published between 2010 and 2015. The soil availability of other important micronutrients can be affected by biochar amendments. Sadowska et al. [91] reported that significantly more soluble Ca, K, and SO42− were found in the soil amended with biochar (pH 7.03) as compared with the control. Similarly, Bista et al. [92] and Marks et al. [93] found that biochar increased soil concentrations of K+ and SO42−, which was attributed to a direct additive effect. Nevertheless, again, the availability of nutrients and micronutrients is influenced by biochar and soil properties.
As mentioned, many of the changes in nutrient cycling are related to specific biochar characteristics (e.g., feedstock, pyrolysis temperature), as well as to how it ages within soil [10]. For instance, biochar has an inhibitory effect on soil aging, and the intermittent addition of fresh biochar biomass may be necessary for optimal nutrient cycling in soil [23]. Furthermore, biochar produced from plant feedstocks generally decomposes faster than biochar produced from wood or grasses [94]. Therefore, the fact that our biochar was produced from orchid pruning biomass could explain its relatively fast decomposition rate, which may have led to the lack of positive and long-lasting effects on plant growth. Indeed, as reported in Trupiano et al. [53] about physicochemical characteristics of the soil amendments used in the present study, the C/N ratio for the biochar was high (125.5), and this may confirm a biochar fast decomposition rate. In addition, soil clay particles have been found to play an important role in biochar stabilization [95]. Thus, the relatively low clay content in the growing substrates may have provided low protection against biochar biophysical degradation.
Another result to point out was that the ability of the biochar and/or compost amendment to induce growth acceleration was not reflected in biomass accumulation. In detail, we found a reduction in the stem biomass of P. euramericana grown on all the amended substrates (B, Co, and BCo) and in the leaf and root biomass of plant grown on the biochar and compost mix. Moreover, we also found a higher S/R ratio in plants grown on the substrate amended with both biochar and compost. The mixture also negatively affected the very fine and fine root length, contrary to the compost addition alone, which showed a great positive impact on this parameter. The results were presumably due to the biochar, which remained ineffective in other experiments [96]. Mertens et al. [96] showed that root length density, fine root dry weight, shoot dry weight, and shoot-root ratio of three-year-old Spondias tuberosa seedlings grown on soil amended with biochar did not report significant differences compared to control soil.
Shoot/root biomass partitioning is an important mechanism by which plants cope with the limitations imposed by growth-constraining resources in the environment [97]. Song et al. [98] indicated that nutrient deficiency (nitrogen and potassium) promoted root growth and increased N and K allocation in storage organs, especially promoting the growth of fine roots. In addition, improving fertilizer under sufficient nutrient conditions did not promote nutrient accumulation in the storage organs, and most of the nutrients were lost with defoliation [98]. In our case, the high value of S/R ratio for poplar plants grown in the biochar-compost mixture confirmed the findings reported above. As previously explained, the combined use of the two amendments might have led to an accumulation of micronutrients, which, in accordance with the study of Song et al. [98], did not promote root growth that could be potentially related also to a high vulnerability to water deficiency [99].
Furthermore, taking into account the fact that plant investment in fine root production is related to efficiently spending resources on water and nutrient uptake [100], the raised concentrations of all nutrient contents in the biochar and compost mix (BCo) should be the cause of very fine and fine root length decreasing [101]. Fine roots are a highly dynamic part of tree biomass that not only have a large influence on forest water and nutrient cycles, but also represent a major source of soil organic carbon [21,64]. The morphological plasticity of roots has been reported in previous studies, especially for fine root fractions [102,103]. In light of this knowledge, we may assert that poplar plants, when growing in substrates amended by the biochar or compost alone, optimized the investment of carbon to dynamically and plastically change root morphology and ensured water and nutrient uptake, without affecting root biomass accumulation [104]. In particular, the compost addition alone enhanced the very fine and fine root length, presumably, for the low soil moisture content, high nutrient levels, and good soil aeration [77,79]; whereas, the compost and biochar combination seemed to confirm an antagonistic effect also on root system development, inducing a decrease in both root biomass and very fine and fine root length.
Plant growth and biomass accumulation are tightly coordinated with photosynthesis to meet the plant demand for the energy required during development [46,47]. Biomass change is the mass balance between production and loss. However, in this counting, it is important to take into account that biomass can be allocated to growth, defense, partitioning, or storage [105], which participate differently in sensu stricto or sensu lato plant growth. Indeed, plant growth in sensu stricto refers to the irreversible increase in total biomass stored as compounds that form the structure of plants (cellulose and lignin in the cell walls, lipids in the membranes, and proteins within the cell). Growth in sensu lato refers to irreversible increase in cell, organ, or plant volume, together with a reversible increase of storage proteins and lipids, secondary metabolites, or nonstructural carbohydrates (e.g., sugars, starch). Storage compounds can be reversible depleted (negative biomass change) for the production, accumulation, and release of secondary metabolites as a buffer against any disturbance-based fluctuation in carbohydrate assimilation, particularly important for plant defense. Thus, structural growth and storage are potentially competing for carbon resources to ensure the growth–defense tradeoff [106] and must therefore be highly regulated.
On the basis of these findings, the accelerated growth rate induced by the biochar could be related to changes in the priority of soluble carbohydrate distribution to growing organs [94] that should be guaranteed by active photosynthetic activity (high leaf chlorophyll content) in plants grown on biochar amended substrates (biochar alone and in combination with the compost).
Several studies showed that exogenous application of biochar increased the chlorophyll content due to the enhanced availability of nutrients and water [107,108] and, contrary to that revealed here, is generally strictly related to a higher plant biomass accumulation. Other studies demonstrated that the addition of biochar could misbalance the photosynthetic machinery and impair the mechanisms recognizing pathogen-derived molecules inducing plant defense machinery dysfunction [109,110]. Thus, we can hypothesize, in the case of poplar plants, a diversion of resources away from energy reserves and toward defense to optimize plant fitness [111].
In particular, considering that in poplar grown on the biochar amended substrates (biochar alone and in combination with compost), the root lignin content increased in accordance with the high leaf chlorophyll content, the carbon flux could be extended in root toward the phenylpropanoid pathway, and possibly includes the biosynthesis of lignin [112,113]. It has recently been reported that biochar induces the up-regulation of several enzymes involved in lignin synthesis that should be indispensable for plant growth-defense tradeoff, acting as an important physical barrier that protects against pathogen invasion and preventing toxic compounds ingress [114,115,116]. However, to date, crosstalk among lignin content and plant growth–defense tradeoff is complex and remains unpredictable due to the limited understanding of the underlying mechanisms [117].
In conclusion, our study showed that biochar and/or compost applications improved growing medium physicochemical characteristics by increasing electrical conductivity, cation exchange capacity, and nutrient concentrations. These ameliorations led to accelerated P. euramericana growth and development—as revealed by poplar phenology evaluation—when the biochar and compost were used alone, whereas the biochar-compost combination induced a slowing in plant bud development. However, the amendment additions to the growing substrate had negligible or negative effects on poplar morphological traits, except for the compost added alone, which increased plant height and very fine and fine root length. The biochar–compost combination was found to have negative effects on plant growth that could have been due to an accumulation of nutrients and/or micronutrients that, at high concentrations, could become toxic and reduce plant development.
Overall, our results showed that the compost addition alone promoted P. euramericana growth without affecting the structural features and, thus, plant biomass accumulation. However, the compost was able to enhance very fine and fine root lengths, guaranteeing water and nutrient uptake. Conversely, the biochar, both alone and in combination with the compost, produced a negative/summative effect that was reflected in a carbon metabolism shift (from primary to secondary) toward lignin biosynthesis for optimizing the growth-defense tradeoff.
Consequently, the compost amendment alone should be the best solution for the nursery-produced poplar seedlings, being able to improve both substrate properties and root system characteristics, which are key aspects in a forest restoration program. These characteristics may make seedlings better able to cope with the period after transplanting, overcoming periods of aridity exacerbated by current climate change, and enhancing plant stability in steep soils.
Future work is anticipated on investigations for the use of biochar and compost in the long term, focusing on all rhizosphere components and functional interaction. Given the importance of nutrient concentration and availability in soil for plant growth, it is critical that future studies provide more comprehensive details on the surface properties of biochar in the soil environment, comparing biochar-nutrient interaction against biochar-compost. Moreover, collectively, our findings suggest that the responses of soil and plants to biochar are strongly influenced by amendment physicochemical characteristics. Therefore, determining the practical effectiveness of biochar in the long term requires additional studies on the type and rate of biochar application, in addition to optimization of feedstock properties and pyrolysis conditions suitable for better biochar usage. Including short-term and long-term evaluation of biochar must complement each other to unravel the possible effect of age on biochar, clarifying the effects of aged versus fresh biochar. It may also be important to evaluate biochar and compost both developed from the same feedstock as a part of future line of research. Additionally, a better understanding of the factors determining very fine and fine root lifespan, turnover, and decomposition will be crucial for a mechanistic insight into tree responses to diverse amendments and changing environments and for the quantification of forest carbon turnover. This information will be useful to scientists/managers involved in smart selection and innovation in properly applying soil amendments in effective forest restoration strategies.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f13040550/s1, Table S1: Seasonal changes during experimental trial.
Author Contributions
Conceptualization, D.C., G.S.S. and D.T.; methodology and formal analysis, E.D.Z., A.M., M.T. and D.T.; software, validation and data curation, M.S.; writing—original draft preparation, M.S. and D.T.; writing—review and editing, M.S., E.D.Z., A.M., M.T., D.C., G.S.S. and D.T.; supervision, D.C., G.S.S. and D.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, Y.; Ye, Y.; Fang, X.; Liu, Y. Reconstruction of agriculture-driven deforestation in western Hunan province of China during the 18th century. Land 2022, 11, 181. [Google Scholar] [CrossRef]
- Skole, D.L.; Mbow, C.; Mugabowindekwe, M.; Brandt, M.S.; Samek, J.H. Trees outside of forests as natural climate solutions. Nat. Clim. Chang. 2021, 11, 1013–1016. [Google Scholar] [CrossRef]
- Hoekman, D.; Kooij, B.; Quiñones, M.; Vellekoop, S.; Carolita, I.; Budhiman, S.; Arief, R.; Roswintiarti, O. Wide-area near-real-time monitoring of tropical forest degradation and deforestation using Sentinel-1. Remote Sens. 2020, 12, 3263. [Google Scholar] [CrossRef]
- Curtis, P.G.; Slay, C.M.; Harris, N.L.; Tyukavina, A.; Hansen, M.C. Classifying drivers of global forest loss. Science 2018, 361, 1108–1111. [Google Scholar] [CrossRef]
- Ritchie, H.; Roser, M. Forests and Deforestation. 2021. Available online: https://ourworldindata.org/forests-and-deforestation (accessed on 20 January 2022).
- Breshears, D.D.; Fontaine, J.B.; Ruthrof, K.X.; Field, J.P.; Feng, X.; Burger, J.R.; Law, D.J.; Kala, J.; Hardy, G.E.S.J. Underappreciated plant vulnerabilities to heat waves. New Phytol. 2021, 231, 32–39. [Google Scholar] [CrossRef]
- Matusick, G.; Ruthrof, K.X.; Brouwers, N.C.; Dell, B.; Hardy, G.S. Sudden forest canopy collapse corresponding with extreme drought and heat in a Mediterranean-type eucalypt forest in south western Australia. Eur. J. For. Res. 2013, 132, 497–510. [Google Scholar] [CrossRef]
- Payne, R.J.; Anderson, A.R.; Sloan, T.; Gilbert, P.; Newton, A.; Ratcliffe, J.; Mauquoy, D.; Jessop, W.; Andersen, R. The future of peatland forestry in Scotland: Balancing economics, carbon and biodiversity. Scott. For. 2018, 100, 34–40. [Google Scholar]
- Brown, I. Challenges in delivering climate change policy through land use targets for afforestation and peatland restoration. Environ. Sci. Policy 2020, 107, 36–45. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Landis, T.D.; Pinto, J.R.; Haase, D.L.; Wilkinson, K.W.; Davis, A.S. Meeting forest restoration challenges: Using the target plant concept. Reforesta 2016, 1, 37–52. [Google Scholar] [CrossRef]
- Mansourian, S. In the eye of the beholder: Reconciling interpretations of forest landscape restoration. Land Degrad. Dev. 2018, 29, 2888–2898. [Google Scholar] [CrossRef]
- Le, H.D.; Smith, C.; Herbohn, J. Identifying interactions among reforestation success drivers: A case study from the Philippines. Ecol. Modell. 2015, 316, 62–77. [Google Scholar] [CrossRef]
- Fargione, J.; Haase, D.L.; Burney, O.T.; Kildisheva, O.A.; Edge, G.; Cook-Patton, S.C.; Chapman, T.; Rempel, A.; Hurteau, M.D.; Davis, K.T.; et al. Challenges to the reforestation pipeline in the United States. Front. For. Glob. Chang. 2021, 4, 8. [Google Scholar] [CrossRef]
- North, M.P.; Stevens, J.T.; Greene, D.F.; Coppoletta, M.; Knapp, E.E.; Latimer, A.M.; Restaino, C.M.; Tompkins, R.E.; Welch, K.R.; York, R.A.; et al. Tamm Review: Reforestation for resilience in dry western US forests. For. Ecol. Manag. 2019, 432, 209–224. [Google Scholar] [CrossRef]
- Puettmann, M.; Sahoo, K.; Wilson, K.; Oneil, E. Life cycle assessment of biochar produced from forest residues using portable systems. J. Clean. Prod. 2020, 250, 119564. [Google Scholar] [CrossRef]
- Davis, A.S.; Pinto, J.R. The scientific basis of the Target Plant Concept: An overview. Forests 2021, 12, 1293. [Google Scholar] [CrossRef]
- Andivia, E.; Villar-Salvador, P.; Oliet, J.A.; Puértolas, J.; Dumroese, R.K. How can my research paper be useful for future meta-analyses on forest restoration plantations? New For. 2019, 50, 255–266. [Google Scholar] [CrossRef]
- Meli, P.; Isernhagen, I.; Brancalion, P.H.; Isernhagen, E.C.; Behling, M.; Rodrigues, R.R. Optimizing seeding density of fast-growing native trees for restoring the Brazilian Atlantic Forest. Restor. Ecol. 2018, 26, 212–219. [Google Scholar] [CrossRef]
- Brancalion, P.H.; Holl, K.D. Guidance for successful tree planting initiatives. J. Appl. Ecol. 2020, 57, 2349–2361. [Google Scholar] [CrossRef]
- Drake, J.A.; Carrucan, A.; Jackson, W.R.; Cavagnaro, T.R.; Patti, A.F. Biochar application during reforestation alters species present and soil chemistry. Sci. Total Environ. 2015, 514, 359–365. [Google Scholar] [CrossRef]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Pinto, J.R.; Fulgaro, N.; Scippa, G.S.; Chiatante, D. Tree seedling response to LED spectra: Implications for forest restoration. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2018, 152, 515–523. [Google Scholar] [CrossRef]
- Barrow, C. Biochar: Potential for countering land degradation and for improving agriculture. Appl. Geogr. 2012, 34, 21–28. [Google Scholar] [CrossRef]
- Lefebvre, D.; Román-Dañobeytia, F.; Soete, J.; Cabanillas, F.; Corvera, R.; Ascorra, C.; Fernandez, L.E.; Silman, M. Biochar effects on two tropical tree species and its potential as a tool for reforestation. Forests 2019, 10, 678. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.; Thies, J.; Masiello, C.; Hockaday, W.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Woolf, D.; Amonette, J.E.; Street-Perrott, F.A.; Lehmann, J.; Joseph, S. Sustainable biochar to mitigate global climate change. Nat. Commun. 2010, 1, 56. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, K.; Miyamoto, T.; Iwata, Y.; Shiono, T. Influences of feedstock and pyrolysis temperature on the nitrate adsorption of biochar. Soil Sci. Plant Nutr. 2016, 62, 180–184. [Google Scholar] [CrossRef]
- Kavitha, B.; Reddy, P.V.L.; Kim, B.; Lee, S.S.; Pandey, S.K.; Kim, K.H. Benefits and limitations of biochar amendment in agricultural soils: A review. J. Environ. Manag. 2018, 227, 146–154. [Google Scholar] [CrossRef]
- Thomas, S.C.; Gale, N. Biochar and forest restoration: A review and meta-analysis of tree growth responses. New For. 2015, 46, 931–946. [Google Scholar] [CrossRef]
- Karim, M.; Halim, M.A.; Gale, N.V.; Thomas, S.C. Biochar effects on soil physiochemical properties in degraded managed ecosystems in northeastern Bangladesh. Soil Syst. 2020, 4, 69. [Google Scholar] [CrossRef]
- Wu, S.; He, H.; Inthapanya, X.; Yang, C.; Lu, L.; Zeng, G.; Han, Z. Role of biochar on composting of organic wastes and remediation of contaminated soils—A review. Environ. Sci. Pollut. Res. 2017, 24, 16560–16577. [Google Scholar] [CrossRef]
- Lal, R. Restoring soil quality to mitigate soil degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef]
- Saer, A.; Lansing, S.; Davitt, N.H.; Graves, R.E. Life cycle assessment of a food waste composting system: Environmental impact hotspots. J. Clean. Prod. 2013, 52, 234–244. [Google Scholar] [CrossRef]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and land-use change: Processes and potential. Glob. Chang. Biol. 2000, 6, 317–327. [Google Scholar] [CrossRef]
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility: A review. Agron. Sustain. Dev. 2010, 30, 401–422. [Google Scholar] [CrossRef]
- Persiani, A.; Diacono, M.; Monteforte, A.; Montemurro, F. Agronomic performance, energy analysis and carbon balance comparing different fertilization strategies in horticulture under Mediterranean conditions. Environ. Sci. Pollut. Res. 2019, 26, 19250–19260. [Google Scholar] [CrossRef] [PubMed]
- Pane, C.; Palese, A.M.; Spaccini, R.; Piccolo, A.; Celano, G.; Zaccardelli, M. Enhancing sustainability of a processing tomato cultivation system by using bioactive compost teas. Sci. Hort. 2016, 202, 117–124. [Google Scholar] [CrossRef]
- Martínez-Blanco, J.; Muñoz, P.; Anton, A.; Rieradevall, J. Life cycle assessment of the use of compost from municipal organic waste for fertilization of tomato crops. Resour. Conserv. Recycl. 2009, 53, 340–351. [Google Scholar] [CrossRef]
- Steiner, C.; Das, K.C.; Melear, N.; Lakly, D. Reducing nitrogen loss during poultry litter composting using biochar. J. Environ. Qual. 2010, 39, 1236–1242. [Google Scholar] [CrossRef]
- Fischer, D.; Glaser, B. Synergisms between compost and biochar for sustainable soil amelioration. In Management of Organic Waste; Kumar, S., Ed.; InTech: Rijeka, Croatia, 2012; pp. 167–198. [Google Scholar]
- Liu, J.; Schulz, H.; Brandl, S.; Miehtke, H.; Huwe, B.; Glaser, B. Short-Term effect of biochar and compost on soil fertility and water status of a Dystric Cambisol in NE Germany under field conditions. J. Plant Nutr. Soil Sci. 2012, 175, 698–707. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Seedling quality: History, application, and plant attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Why seedlings survive: Importance of plant attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; South, D.B. Seedling quality of southern pines: Influence of plant attributes. Tree Plant. Notes 2017, 60, 29–40. [Google Scholar]
- Pinto, J.R. Morphology targets: What do seedling morphological attributes tell us. In National Proceedings: Forest and Conservation Nursery Associations-2010; RMRS-P-65; U.S. Department of Agriculture, Forest Service: Fort Collins, CO, USA, 2011; pp. 74–79. [Google Scholar]
- Jha, K.K. Root carbon sequestration and its efficacy in forestry and agroforestry systems: A case of Populus euramericana I-214 cultivated in Mediterranean condition. Not. Sci. Biol. 2018, 10, 68–78. [Google Scholar] [CrossRef]
- Fuertes, A.; Oliveira, N.; Cañellas, I.; Sixto, H.; Rodríguez-Soalleiro, R. An economic overview of Populus spp. in Short Rotation Coppice systems under Mediterranean conditions: An assessment tool for decision-making. Renew. Sustain. Energy Rev. 2021, 151, 111577. [Google Scholar] [CrossRef]
- Pallardy, S.G.; Gibbins, D.E.; Rhoads, J.L. Biomass production by two-year-old poplar clones on floodplain sites in the Lower Midwest, USA. Agrofor. Syst. 2003, 59, 21–26. [Google Scholar] [CrossRef]
- Nielsen, U.B.; Madsen, P.; Hansen, J.K.; Nord-Larsen, T.; Nielsen, A.T. Production potential of 36 poplar clones grown at medium length rotation in Denmark. Biomass Bioenergy 2014, 64, 99–109. [Google Scholar] [CrossRef]
- Giri, A.; Kumar, G.; Arya, R.; Mishra, S.; Mishra, A.K. Carbon sequestration in Populus deltoides based agroforestry system in northern India. Int. J. Chem. Stud. 2019, 7, 2184–2188. [Google Scholar]
- VanWallendael, A.; Lowry, D.B.; Hamilton, J.A. One hundred years into the study of ecotypes, new advances are being made through large-scale field experiments in perennial plant systems. Curr. Plant Biol. 2022, 66, 102152. [Google Scholar] [CrossRef] [PubMed]
- U.S Department of Agriculture, Natural Resources Conservation Service. National Soil Survey Handbook. Available online: http://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/ref/?cid=nrcs142p2_054242 (accessed on 27 January 2022).
- Alfano, G.; Lustrato, G.; Lima, G.; Vitullo, D.; Ranalli, G. Characterization of composted olive mill wastes to predict potential plant disease suppressiveness. Biol. Control 2011, 58, 199–207. [Google Scholar] [CrossRef]
- Trupiano, D.; Cocozza, C.; Baronti, S.; Amendola, C.; Vaccari, F.P.; Lustrato, G.; Di Lonardo, S.; Fantasma, F.; Tognetti, R.; Scippa, G.S. The effects of biochar and its combination with compost on lettuce (Lactuca sativa L.) growth, soil properties, and soil microbial activity and abundance. Int. J. Agron. 2017, 12, 3158207. [Google Scholar] [CrossRef]
- Black, C.A. Methods of Soil Analysis: Part I Physical and Mineralogical Properties; American Society of Agronomy: Madison, WI, USA, 1965. [Google Scholar]
- Rayment, G.E.; Higginson, F.R. Australian Laboratory Handbook of Soil and Water Chemical Methods; Inkata Press Pty Ltd.: Melbourne, Australia, 1992. [Google Scholar]
- Jones, J.B., Jr.; Kalra, Y.P. Soil testing and plant analysis activities-The United States and Canada. Commun. Soil Sci. Plant Anal. 1992, 23, 2015–2027. [Google Scholar] [CrossRef]
- Mehlich, A. Use of triethanolamine acetate-barium hydroxide buffer for the determination of some base exchange properties and lime requirement of soil. Soil Sci. Soc. Am. Proc. 1938, 29, 374–378. [Google Scholar] [CrossRef]
- Dumas, J.B.A. Procédés de l’analyse organique. Ann. Chim. Phys. 1831, 247, 198–213. [Google Scholar]
- Bowman, R.A. A rapid method to determine total phosphorus in soils. Soil Sci. Soc. Am. J. 1988, 52, 1301–1304. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. USDA Circular 939; U.S. Government Publishing Office: Washington, DC, USA, 1954. [Google Scholar]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analysis of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Beretta, N.; Silbermann, A.V.; Paladino, L.; Torres, D.; Bassahun, D.; Musselli, R.; Lamohte, A.G. Soil texture analyses using a hydrometer modification of the Bouyoucos method. Cienc. Investig. Agrar. 2014, 41, 263–271. [Google Scholar] [CrossRef]
- Kemper, W.D.; Koch, E.J. Aggregate Stability of Soils from Western USA and Canada USDA Technical Bulletin No.1355; US Government Printing Office: Washington, DC, USA, 1966. [Google Scholar]
- Montagnoli, A.; Di Iorio, A.; Terzaghi, M.; Trupiano, D.; Scippa, G.S.; Chiatante, D. Influence of soil temperature and water content on fine-root seasonal growth of European beech natural forest in Southern Alps, Italy. Eur. J. For. Res. 2014, 133, 957–968. [Google Scholar] [CrossRef]
- Trupiano, D.; Rocco, M.; Renzone, G.; Scaloni, A.; Montagnoli, A.; Terzaghi, M.; Di Iorio, A.; Chiatante, D.; Scippa, G.S. Poplar woody root proteome during the transition dormancy-active growth. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2013, 147, 1095–1100. [Google Scholar] [CrossRef][Green Version]
- Doster, M.A.; Bostock, R.M. Quantification of lignin formation in almond bark in response to wounding and infection by Phytophthora species. Phytopathology 1988, 784, 73–477. [Google Scholar]
- Trupiano, D.; Di Iorio, A.; Montagnoli, A.; Lasserre, B.; Rocco, M.; Grosso, A.; Scaloni, A.; Marra, M.; Chiatante, D.; Scippa, G.S. Involvement of lignin and hormones in the response of woody poplar taproots to mechanical stress. Physiol. Plant. 2012, 146, 39–52. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll a and b in N,N Dimethylformamide and 80% acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef]
- Kroetsch, D.; Wang, C. Particle size distribution. Soil Sampl. Methods Anal. 2008, 2, 713–725. [Google Scholar]
- Wolf, M.; Lehndorff, E.; Wiesenberg, G.L.B.; Stockhausen, M.; Schward, L.; Amelung, W. Towards reconstruction of past fire regimes from geochemical analysis of charcoal. Org. Geochem. 2013, 55, 11–21. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Spokas, K.A.; Novak, J.M.; Lentz, R.D.; Cantrell, K.B. Biochar elemental composition and factors influencing nutrient retention. In Biochar for Environmental Management: Science, Technology and Implementation, 2nd ed.; Lehmann, J., Joseph, S., Eds.; Routledge: New York, NY, USA, 2015; pp. 137–161. [Google Scholar]
- Ippolito, J.A.; Cui, L.; Kammann, C.; Wrage-Mönnig, N.; Estavillo, J.M.; Fuertes-Mendizabal, T.; Cayuela, M.L.; Sigua, G.; Novak, J.; Spokas, K.; et al. Feedstock choice, pyrolysis temperature and type influence biochar characteristics: A comprehensive meta-data analysis review. Biochar 2020, 2, 421–438. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Berry, C.M.; Strawn, D.G.; Novak, J.M.; Levine, J.; Harley, A. Biochars reduce mine land soil bioavailable metals. J. Environ. Qual. 2017, 46, 411–419. [Google Scholar] [CrossRef]
- Khan, N.; Clark, I.; Sánchez-Monedero, M.A.; Shea, S.; Meier, S.; Bolan, N. Maturity indices in co-composting of chicken manure and sawdust with biochar. Bioresour. Technol. 2014, 168, 245–251. [Google Scholar] [CrossRef]
- Zhang, J.; Zeng, G.; Chen, Y.; Yu, M.; Yu, Z.; Li, H.; Yu, Y.; Huang, H. Effects of physico-chemical parameters on the bacterial and fungal communities during agricultural waste composting. Bioresour. Technol. 2011, 102, 2950–2956. [Google Scholar] [CrossRef]
- Reyes-Torres, M.; Oviedo-Ocaña, E.R.; Dominguez, I.; Komilis, D.; Sánchez, A. A systematic review on the composting of green waste: Feedstock quality and optimization strategies. Waste Manag. 2018, 77, 486–499. [Google Scholar] [CrossRef]
- Järvis, J.; Ivask, M.; Nei, L.; Kuu, A.; Luud, A. Effect of green waste compost application on afforestation success. Balt. For. 2016, 22, 90–97. [Google Scholar]
- Heiskanen, J.; Hagner, M.; Ruhanen, H.; Mäkitalo, K. Addition of recyclable biochar, compost and fibre clay to the growth medium layer for the cover system of mine tailings: A bioassay in a greenhouse. Environ. Earth Sci. 2020, 79, 422. [Google Scholar] [CrossRef]
- Lazdina, D.; Bardule, A.; Lazdins, A.; Stola, J. Use of waste water sludge and wood ash as fertiliser for Salix cultivation in acid peat soils. Agron. Res. 2011, 9, 305–314. [Google Scholar]
- Gul, S.; Whalen, J.K. Biochemical cycling of nitrogen and phosphorus in biochar amended soils. Soil Biol. Biochem. 2016, 103, 1–15. [Google Scholar] [CrossRef]
- Seehausen, M.L.; Gale, N.V.; Dranga, S.; Hudson, V.; Liu, N.; Michener, J.; Thurston, E.; Williams, C.; Smith, S.M.; Thomas, S.C. Is there a positive synergistic effect of biochar and compost soil amendments on plant growth and physiological performance? Agronomy 2017, 7, 13. [Google Scholar] [CrossRef]
- Tessier, J.T.; Raynal, D.J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. Soil Fertility and Fertilizers; Collier Macmillan Publishers: London, UK, 1985. [Google Scholar]
- Xu, G.; Zhang, Y.; Sun, J.; Shao, H. Negative interactive effects between biochar and phosphorus fertilization on phosphorus availability and plant yield in saline sodic soil. Sci. Total Environ. 2016, 568, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Kammann, C.I.; Shepherd, J.G.; Conte, P.; Schmidt, H.P.; Hagemann, N.; Rich, A.M.; Marjo, C.E.; Allen, J.; Munroe, P.; et al. Microstructural and associated chemical changes during the composting of a high temperature biochar: Mechanisms for nitrate, phosphate and other nutrient retention and release. Sci. Total Environ. 2018, 618, 1210–1223. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Bahar, M.M.; Sarkar, B.; Donne, S.W.; Ok, Y.S.; Palansooriya, K.N.; Kirkham, M.B.; Chowdhury, S.; Bolan, N. Biochar and its importance on nutrient dynamics in soil and plant. Biochar 2020, 2, 379–420. [Google Scholar] [CrossRef]
- Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
- Liu, Z.; He, T.; Cao, T.; Yang, T.; Meng, J.; Chen, W. Effects of biochar application on nitrogen leaching, ammonia volatilization and nitrogen use efficiency in two distinct soils. J. Soil Sci. Plant Nutr. 2017, 17, 515–528. [Google Scholar] [CrossRef]
- Spokas, K.A.; Novak, J.M.; Venterea, R.T. Biochar’s role as an alternative N-fertilizer: Ammonia capture. Plant Soil 2012, 350, 35–42. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Xu, C.Y.; Tahmasbian, I.; Che, R.; Xu, Z.; Zhou, X.; Wallace, H.M.; Bai, S.H. Effects of biochar on soil available inorganic nitrogen: A review and metaanalysis. Geoderma 2017, 288, 79–96. [Google Scholar] [CrossRef]
- Sadowska, U.; Domagała-Świątkiewicz, I.; Żabiński, A. Biochar and its effects on plant–soil macronutrient cycling during a three-year field trial on sandy soil with peppermint (Mentha piperita L.). Part I: Yield and macro element content in soil and plant biomass. Agronomy 2020, 10, 1950. [Google Scholar] [CrossRef]
- Bista, P.; Ghimire, R.; Machado, S.; Pritchett, L. Biochar effects on soil properties and wheat biomass vary with fertility management. Agronomy 2019, 9, 623. [Google Scholar] [CrossRef]
- Marks, A.N.; Mattan, S.; Alcañiz, J.M.; Pérez-Herrero, E.; Domene, X. Gasifier biochar effects on nutrient availability, organic matter mineralization, and soil fauna activity in a multi-year Mediterranean trial. Agric. Ecosyst. Environ. 2016, 215, 30–39. [Google Scholar] [CrossRef]
- Wang, J.; Xiong, Z.; Kuzyakov, Y. Biochar stability in soil: Metaanalysis of decomposition and priming effects. Glob. Chang. Biol. Bioenergy 2016, 8, 512–523. [Google Scholar] [CrossRef]
- Ventura, M.; Alberti, G.; Panzacchi, P.; Vedove, G.D.; Miglietta, F.; Tonon, G. Biochar mineralization and priming effect in a poplar short rotation coppice from a 3-year field experiment. Biol. Fertil. Soils 2019, 55, 67–78. [Google Scholar] [CrossRef]
- Mertens, J.; Germer, S.; Germer, J.; Sauerborn, J. Comparison of soil amendments for reforestation with a native multipurpose tree under semiarid climate: Root and root tuber response of Spondias tuberosa. For. Ecol. Manag. 2017, 396, 1–10. [Google Scholar] [CrossRef]
- Mašková, T.; Herben, T. Root: Shoot ratio in developing seedlings: How seedlings change their allocation in response to seed mass and ambient nutrient supply. Ecol. Evol. 2018, 8, 7143–7150. [Google Scholar] [CrossRef]
- Song, X.; Wan, F.; Chang, X.; Zhang, J.; Sun, M.; Liu, Y. Effects of nutrient deficiency on root morphology and nutrient allocation in Pistacia chinensis Bunge seedlings. Forests 2019, 10, 1035. [Google Scholar] [CrossRef]
- Jordan-Meille, L.; Martineau, E.; Bornot, Y.; Lavres, J.; Abreu-Junior, C.H.; Domec, J.C. How does water-stressed corn respond to potassium nutrition? A shoot-root scale approach study under controlled conditions. Agriculture 2018, 8, 180. [Google Scholar] [CrossRef]
- West, J.B.; Espeleta, J.F.; Donovan, L.A. Fine root production and turnover across a complex edaphic gradient of a Pinus palustris–Aristida stricta savanna ecosystem. For. Ecol. Manag. 2004, 189, 397–406. [Google Scholar] [CrossRef]
- Cheng, Y.; Han, Y.; Wang, Q.; Wang, Z. Seasonal dynamics of fine root biomass, root length density, specific root length, and soil resource availability in a Larix gmelinii plantation. Front. Biol. China 2006, 1, 310–317. [Google Scholar] [CrossRef]
- Bjork, R.G.; Majdi, M.; Klemedtsson, L.; Jonsson, L.L.; Molau, U. Long-term warming effects on root morphology, root mass distribution, and microbial activity in two dry tundra plant communities in northern Sweden. New Phytol. 2007, 176, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Makita, N.; Hirano, Y.; Mizoguchi, T.; Kominami, Y.; Dannoura, M.; Ishii, H.; Finer, L.; Kanazawa, Y. Very fine roots respond to soil depth: Biomass allocation, morphology, and physiology in a broad-leaved temperate forest. Ecol. Res. 2011, 26, 95–104. [Google Scholar] [CrossRef]
- Amendola, C.; Montagnoli, A.; Terzaghi, M.; Trupiano, D.; Oliva, F.; Baronti, S.; Miglietta, F.; Chiatante, D.; Scippa, G.S. Short-term effects of biochar on grapevine fine root dynamics and arbuscular mycorrhizae production. Agric. Ecosyst. Environ. 2017, 239, 236–245. [Google Scholar] [CrossRef]
- Hilty, J.; Muller, B.; Pantin, F.; Leuzinger, S. Tansley review: Plant growth: The what, the how, and the why. New Phytol. 2021, 232, 25–41. [Google Scholar] [CrossRef]
- Lind, E.M.; Borer, E.; Seabloom, E.; Adler, P.; Bakker, J.D.; Blumenthal, D.M.; Crawley, M.; Davies, K.; Firn, J.; Gruner, D.S.; et al. Life-history constraints in grassland plant species: A growth-defence trade-off is the norm. Ecol. Lett. 2013, 16, 513–521. [Google Scholar] [CrossRef]
- Agegnehu, G.; Bass, A.M.; Nelson, P.N.; Bird, M.I. Benefits of biochar, compost and biochar–compost for soil quality, maize yield and greenhouse gas emissions in a tropical agricultural soil. Sci. Total Environ. 2016, 543, 295–306. [Google Scholar] [CrossRef]
- Zhang, D.; Pan, G.; Wu, G.; Kibue, G.W.; Li, L.; Zhang, X.; Zheng, J.; Zheng, J.; Cheng, K.; Joseph, S. Biochar helps enhance maize productivity and reduce greenhouse gas emissions under balanced fertilization in a rainfed low fertility inceptisol. Chemosphere 2016, 142, 106–113. [Google Scholar] [CrossRef]
- Viger, M.; Hancock, R.D.; Miglietta, F.; Taylor, G. More plant growth but less plant defence? First global gene expression data for plants grown in soil amended with biochar. Glob. Chang. Biol. Bioenergy 2015, 7, 658–672. [Google Scholar] [CrossRef]
- Polzella, A.; De Zio, E.; Arena, S.; Scippa, G.S.; Scaloni, A.; Montagnoli, A.; Chiatante, D.; Trupiano, D. Toward an understanding of mechanisms regulating plant response to biochar application. Plant Biosyst. 2019, 153, 163–172. [Google Scholar] [CrossRef]
- Simiele, M.; Sferra, G.; Lebrun, M.; Renzone, G.; Bourgerie, S.; Scippa, G.S.; Morabito, D.; Scaloni, A.; Trupiano, D. In-depth study to decipher mechanisms underlying Arabidopsis thaliana tolerance to metal (loid) soil contamination in association with biochar and/or bacteria. Environ. Exp. Bot. 2021, 182, 104335. [Google Scholar] [CrossRef]
- Amthor, J.S. Efficiency of lignin biosynthesis: A quantitative analysis. Ann. Bot. 2003, 91, 673–695. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.A.; Campbell, M.M. The genetic control of lignin deposition during plant growth and development. New Phytol. 2004, 164, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Ithal, N.; Recknor, J.; Nettleton, D.; Maier, T.; Baum, T.J.; Mitchum, M.G. Developmental transcript profiling of cyst nematode feeding cells in soybean roots. Mol. Plant Microbe Interact. 2007, 20, 510–525. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Storme, V.; Vanholme, B.; Sundin, L.; Christensen, J.H.; Goeminne, G.; Halpin, C.; Rohde, A.; Morreel, K.; Boerjan, W. A systems biology view of responses to lignin biosynthesis perturbations in Arabidopsis. Plant Cell 2012, 24, 3506–3529. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Alkan, N.; Elad, Y.; Sela, N.; Philosoph, A.M.; Graber, E.R.; Frenkel, O. Molecular insights into biochar-mediated plant growth promotion and systemic resistance in tomato against Fusarium crown and root rot disease. Sci. Rep. 2020, 10, 13934. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).