
Environmental Effects among Differently Located and Fertile Sites on Forest Basal-Area Increment in Temperate Zone

Abstract
:1. Introduction
2. Material and Methods
2.1. Geographical Pattern
2.2. Field Sampling
2.3. Time-Series Analysis
2.4. Statistical Assessment
3. Results
3.1. Management Populations of Forest Growth Conditions
3.2. Properties of Assessed Ecosystems
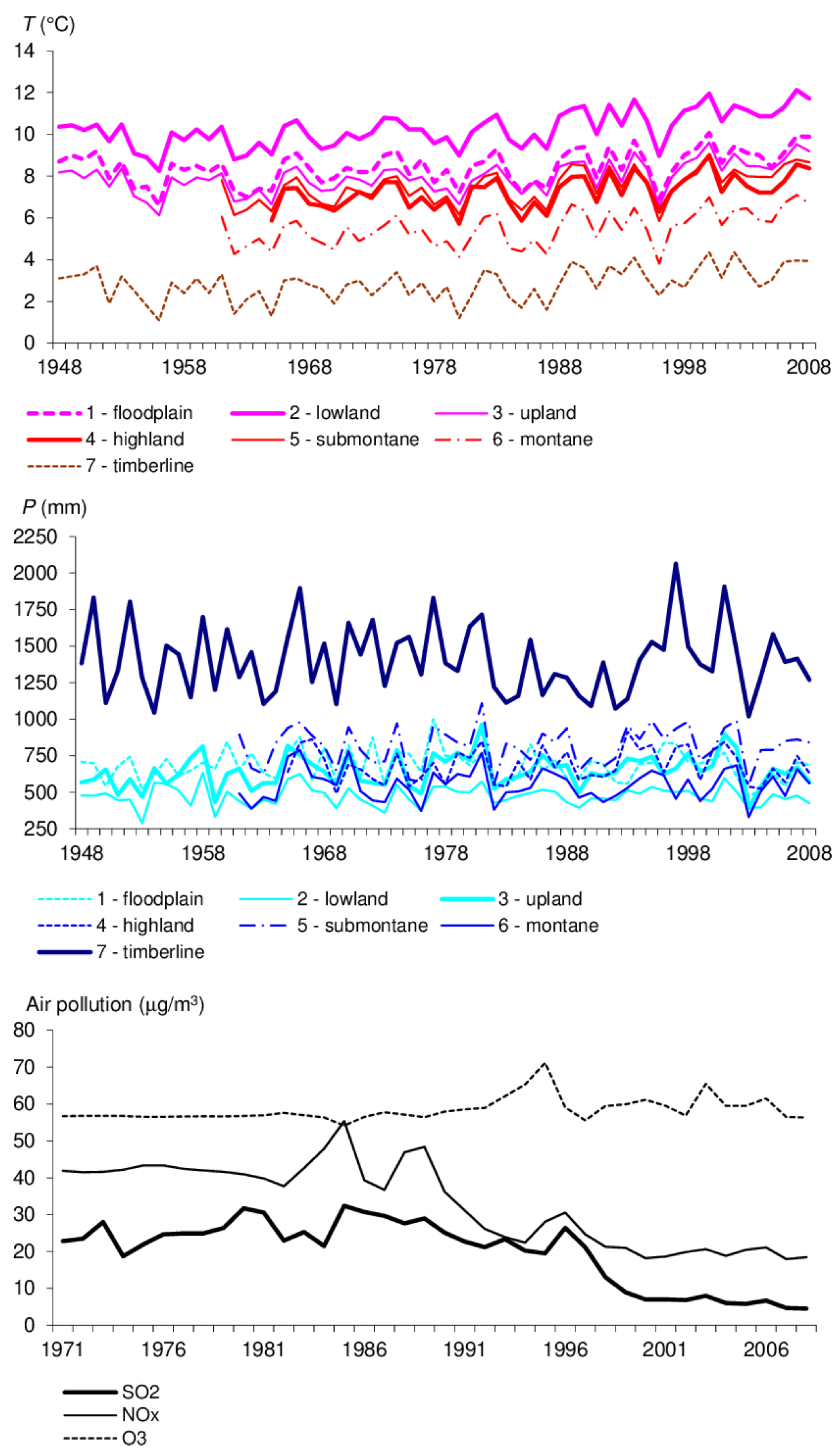
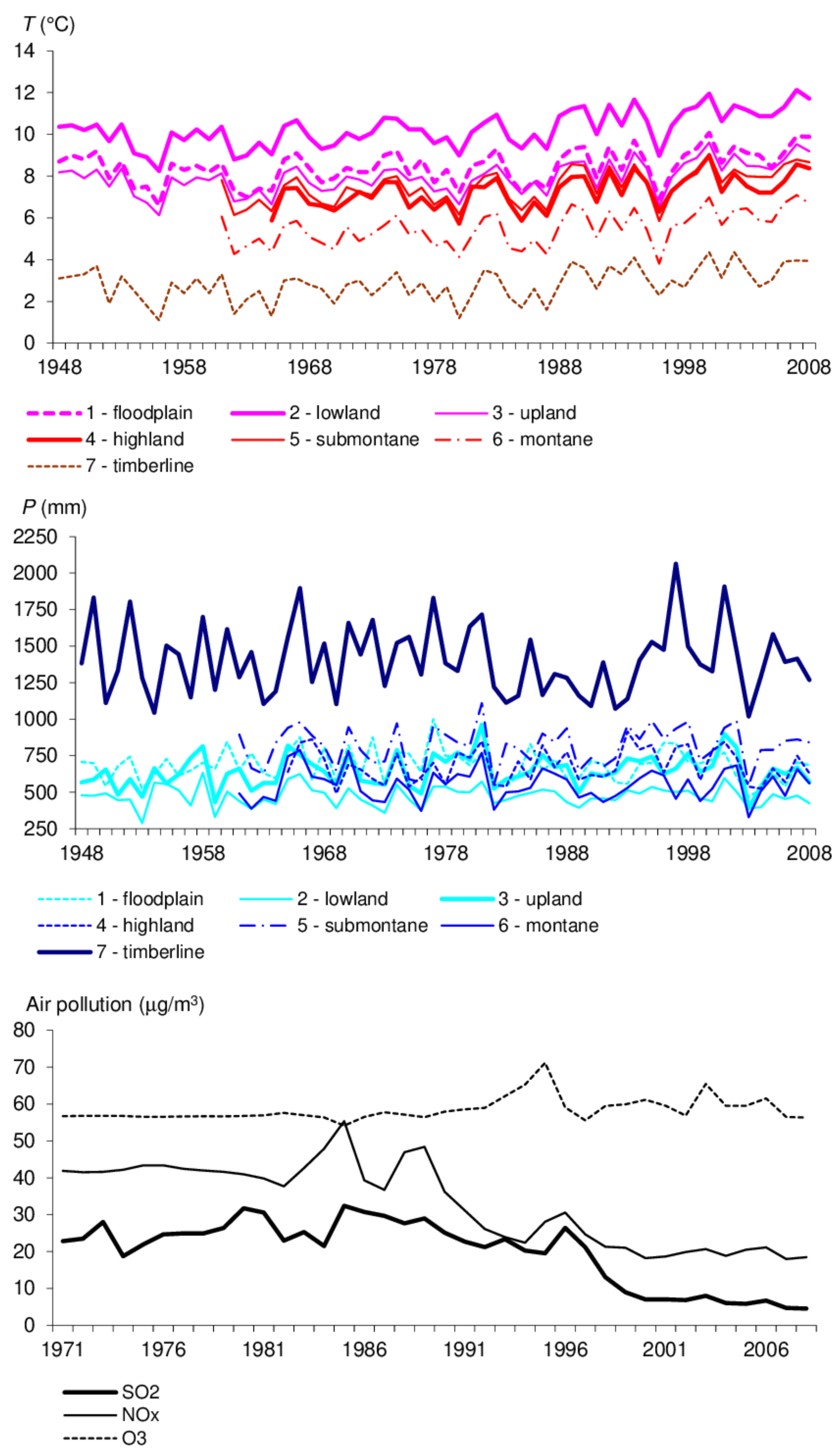
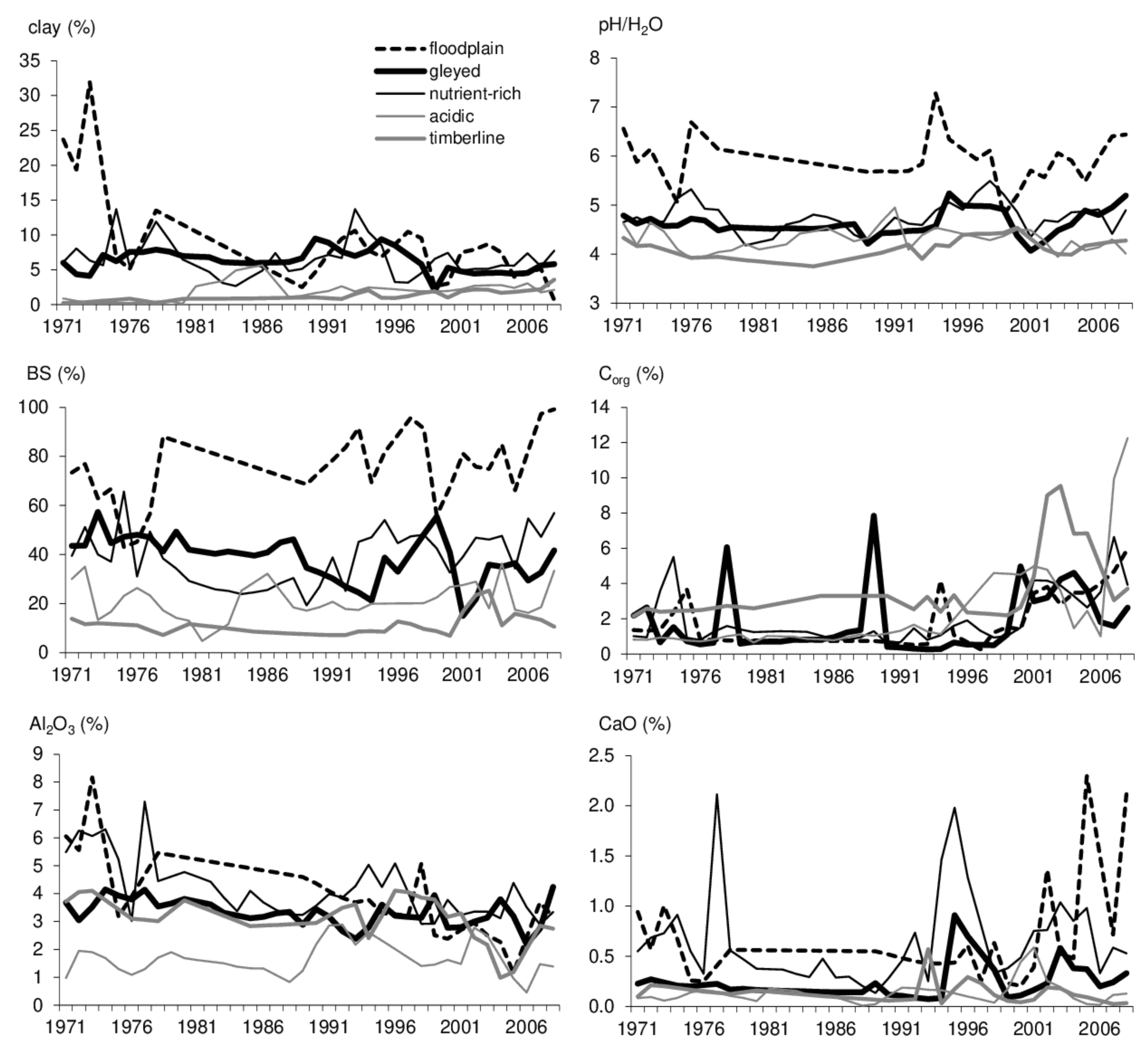
3.2.1. Growth Conditions
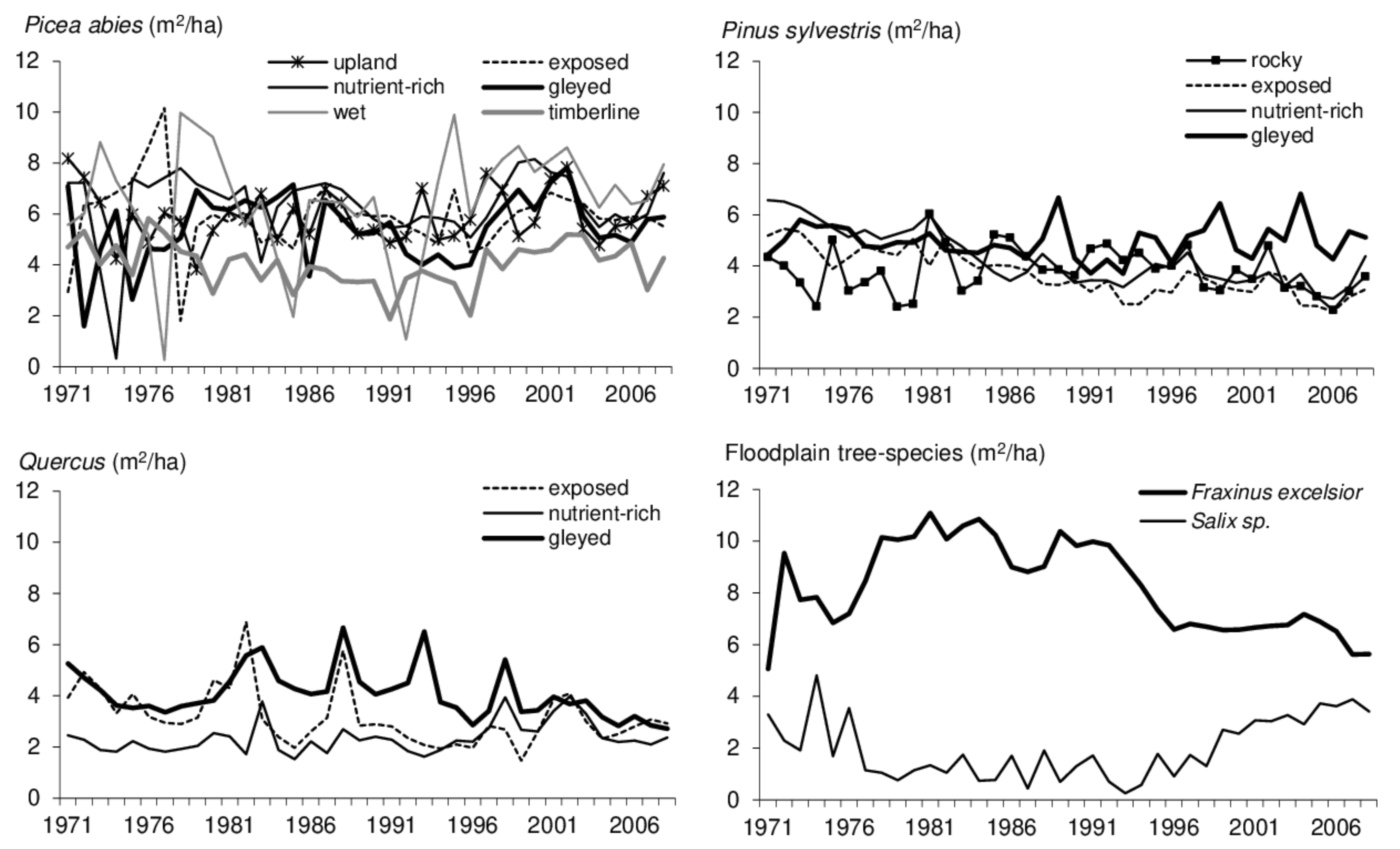
3.2.2. Stand Types
3.3. Response of Forest Stands
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Augustin, S.; Bolte, A.; Holzhausen, M.; Wolff, B. Exceedance of critical loads of nitrogen and sulphur and its relation to forest conditions. Eur. J. For. Res. 2005, 124, 289–300. [Google Scholar] [CrossRef]
- Yu, L.; Zanchi, G.; Akselsson, C.; Wallander, H.; Belyazid, S. Modeling the forest phosphorus nutrition in a southwestern Swedish forest site. Ecol. Modell. 2018, 369, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Motta, R.; Borghetti, M. Conifers in cold environment synchronize maximum growth rate of tree-ring formation with day lenght. New Phytol. 2006, 170, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Tomášková, I.; Pokorný, R.; Marek, M.V. Influence of stand density, thinning and elevated CO2 on stem wood density on spruce. J. For. Sci. 2007, 53, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Esper, J.; Frank, D.C.; Timonen, M.; Zorita, E.; Wilson, R.J.S.; Luterbacher, J.; Holzkämper, S.; Fischer, N.; Wagner, S.; Nievergelt, D.; et al. Orbital forcing of tree-ring data. Nat. Clim. Change 2012, 2, 862–866. [Google Scholar] [CrossRef]
- Samec, P. Analysis of paleoclimatic time series for evaluation of recent climate change on European North Atlantic Region and Bohemian Massif selected localities. Rep. For. Res. 2006, 51, 272–278. [Google Scholar]
- Ganopolski, A.; Winhelmann, R.; Schellnhuber, H.J. Critical insolation–CO2 relation for diagnosing past and future glacial inception. Nature 2016, 529, 200–203. [Google Scholar] [CrossRef]
- Miyake, F.; Panyushkina, I.P.; Jull, A.J.T.; Adolphi, F.; Brehm, N.; Helama, S.; Kanzawa, K.; Moriya, T.; Muscheler, R.; Nicolussi, K.; et al. A Single-Year Cosmic Ray Event at 5410 BCE Registered in 14C of Tree Rings. Geophys. Res. Lett. 2021, 48, e2021GL093419. [Google Scholar] [CrossRef]
- Liu, S.; Bond-Lamberty, B.; Hicke, J.A.; Vargas, R.; Zhao, S.; Chen, J.; Edburg, S.L.; Hu, Y.; Liu, J.; McGuire, A.D.; et al. Simulating the impacts of disturbances on forest carbon cycling in North America: Processes, data, models, and challenges. J. Geophys. Res. 2011, 116, G00K08. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-S.; Hung, D.V.; Kwak, J.-H.; Lim, S.-S.; Lee, K.-H.; Choi, W.-J. Tree Ring Ca/Al as an indicator of Historical Soil Acidification of Pinus densiflora Forest in Southern Korea. Korean J. Environ. Agric. 2011, 30, 229–233. [Google Scholar] [CrossRef]
- Chen, L.; Wu, F.-H.; Liu, T.-W.; Chen, J.; Li, Z.-J.; Pei, Z.-M.; Zheng, H.-L. Soil acidity reconstruction nased on tree ring information of a dominant species Abies fabri in the subalpine forest ecosystems in southwest China. Environ. Pollut. 2010, 158, 3219–3224. [Google Scholar] [CrossRef]
- Ngao, J.; Epron, D.; Delpierre, N.; Bréda, N.; Granier, A.; Longdoz, B. Spatial variability of soil CO2 efflux linked to soil parameters and ecosystem characteristics in a temperate beech forest. Agric. For. Meteorol. 2012, 154–155, 136–146. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution Global Maps of 21st-Centrury Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [Green Version]
- Vincent, C.; Le Meur, E.; Six, D. Solving the paradox of the end of the Little Ice Age in the Alps. Geophys. Res. Lett. 2005, 32, L09706. [Google Scholar] [CrossRef]
- Jones, P.D.; Mann, M.E. Climate Over Past Millennia. Rev. Geophys. 2004, 42, 143. [Google Scholar] [CrossRef] [Green Version]
- Mann, M.E.; Jones, P.D. Global surface temperatures over the past two millennia. Geophys. Res. Lett. 2003, 30, 17814. [Google Scholar] [CrossRef] [Green Version]
- Briffa, K.R.; Osborn, T.J.; Schweingruber, F.H.; Harris, I.C.; Jones, P.D.; Shiyatov, S.G.; Vaganov, E.A. Low-frequency Temperature Variations from a Nothern Tree Ring Density Network. J. Geophys. Res. 2001, 106, 2629–2941. [Google Scholar] [CrossRef]
- Osborn, T.J.; Briffa, K.R. The spatial extent of 20th century warmth in the context of the past 1200 years. Science 2006, 311, 841–844. [Google Scholar] [CrossRef]
- Gül, E.; Erşahin, S. Evaluating the desertification vulnerability of a semiarid landscape under different land uses with the environmental sensitivity index. Land Degrad. Dev. 2019, 30, 811–823. [Google Scholar] [CrossRef]
- Augustin, S. Linking critical thresholds for acidity to forest condition by using element contents in tree rings: Consequences for the development of an integrated ecosystem assessment. iForest 2009, 2, 51–53. [Google Scholar] [CrossRef]
- Srivastava, P.; Singh, R.; Tripathi, S.; Singh, P.; Singh, S.; Singh, H.; Raghubanshi, A.S.; Mishra, P.K. Soil Carbon Dynamics Under Changing Climate—A Research Transition from Absolute to Relative Roles of Inorganic Nitrogen Pools and Associated Microbial Processes: A Review. Pedoshepere 2017, 27, 792–806. [Google Scholar] [CrossRef]
- Ďurský, J.; Škvarenina, J.; Mindáš, J.; Miková, A. Regional analysis of climate change impact on Norway spruce (Picea abies L. Karst) growth in Slovak mountain forests. J. For. Sci. 2006, 51, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Sigurdsson, B.D.; Medhurst, J.L.; Wallin, G.; Eggertsson, O.; Linder, S. Growth of mature boreal Norway spruce was not affected by elevated [CO2] and/or air temperature unless nutrient availability was improved. Tree Physiol. 2013, 33, 1192–1205. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carrer, M. Age-dependent xylogenesis in timberline conifers. New Phytol. 2008, 177, 199–208. [Google Scholar] [CrossRef]
- Ziche, D.; Seidling, W. Homogenisation of climate time series from ICP Forests Level II monitoring sites in Germany based on interpolated climate data. Ann. For. Sci. 2010, 67, 804. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.; Yang, L.; Zhu, A.-X.; Rossiter, D.G.; Liu, J.; Liu, J.; Qin, C.; Wang, D. Mapping soil organic matter concentration at different scales using a mixed geographically weighted regression method. Geoderma 2016, 281, 69–82. [Google Scholar] [CrossRef]
- Minasny, B.; McBratney, A.B.; Salvador-Blanes, S. Quantitative models for pedogenesis—A review. Geoderma 2008, 144, 140–157. [Google Scholar] [CrossRef]
- Debel, A.; Meier, W.J.-H.; Bräuning, A. Climate Signals for Growth Variations of F. sylvatica, P. abies and P. sylvestris in Southeast Germany over the Past 50 Years. Forests 2021, 12, 1433. [Google Scholar] [CrossRef]
- Neuhäuslová, Z.; Blažková, D.; Grulich, V.; Husová, M.; Chytrý, M.; Jeník, J.; Jirásek, J.; Kolbek, J.; Kropáč, Z.; Ložek, V.; et al. Map of Potential Natural Vegetation of the Czech Republic /Text part/; Academia: Prague, Czech Republic, 1998; pp. 1–341. [Google Scholar]
- Šindelář, J. A preliminary report on autochthonous species composition of forests in the territory of the Czech Republic. Lesnictví For. 1995, 41, 293–299. [Google Scholar]
- Zouhar, V. Dřevinná skladba. In Oblastní Plány Rozvoje Lesů 2; ÚHÚL Brandýs nad Labem, Česká Lesnická Společnost, Ministerstvo Zemědělství České Republiky: Prague, Czech Republic, 2018; pp. 20–22. [Google Scholar]
- Samec, P.; Caha, J.; Tuček, P.; Zapletal, M.; Cudlín, P.; Kučera, M. Discrimination between acute and chronic decline of Central European forests using map algebra of the growth condition and forest biomass fuzzy sets: A case study. Sci. Total Environ. 2017, 599–600, 899–909. [Google Scholar] [CrossRef]
- Mikšovský, J.; Brázdil, R.; Štěpánek, P.; Zahradníček, P.; Pišoft, P. Long-term variability of temperature and precipitation in the Czech Lands: An attribution analysis. Clim. Chang. 2014, 125, 253–264. [Google Scholar] [CrossRef]
- Horn, P.S.; Pesce, A.J.; Copeland, B.E. A robust approach to reference interval estimation and evaluation. Clin. Chem. 1998, 44, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Peter, M.; Ayer, F.; Cudlín, P.; Egli, S. Belowground ectomycorrhizal communities in three Norway spruce stands with different degrees of decline in the Czech Republic. Mycorrhiza 2008, 18, 157–169. [Google Scholar] [CrossRef] [Green Version]
- Neteler, M.; Mitášova, H. Open Source GIS: A GRASS GIS Approach; Kluwer Academic Publishers: Oxford, UK; Springer: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Culek, M.; Grulich, V. Biogeographical division. 1:500,000. In Landscape Atlas of the Czech Republic; Hrnčianová, T., Mackovčin, P., Zvara, I., Eds.; Ministry of Environment, The Silva Tarouca Research Institute for Landscape and Ornamental Gardering: Prague, Czech Republic, 2009; pp. 134–135. [Google Scholar]
- Schad, P.; van Huysteen, C.; Michéli, E.; Vargas, R. (Eds.) World Reference Base for Soil Resources 2014. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2014. [Google Scholar]
- Cools, N.; De Vos, B. Availability and evaluation of European forest soil monitoring data in the study on the effects of air pollution on forests. iForest 2011, 4, 205–211. [Google Scholar] [CrossRef]
- Gut, U.; Árvai, M.; Bijak, S.; Camarero, J.; Cedro, A.; Cruz-García, R.; Garamszegi, B.; Hacket-Pain, A.; Hevia, A.; Huang, W.; et al. No systematic effects of sampling direction on climate-growth relationships in a large-scale, multi-species tree-ring data set. Dendrochronologia 2019, 57, 125624. [Google Scholar] [CrossRef]
- Mann, M.E.; Bradley, R.S.; Hughes, M.K. Global-Scale Temperature Patterns and Climate Forcing Over the Past Six Centuries. Nature 1998, 392, 779–787. [Google Scholar] [CrossRef]
- Feliksik, E.; Wilczyński, S.; Durło, G. Variability of air temperatures of the temperate climatic beltof the Beskid Sądecki Mountains in the period 1971–2000 as exemplified by the climatological station in Kopciowa. Electron. J. Pol. Agric. Univ. 2001, 4, 2. [Google Scholar]
- Hadas, P. Analysis of the Climate as a Stress Factor to Forest Ecosystems. Ekologia Bratislava 2000, 19, 162–176. [Google Scholar]
- Zapletal, M. Atmospheric deposition of nitrogen and sulphur in relation to critical loads of nitrogen and acidity in the Czech Republic. J. For. Sci. 2006, 52, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Oulehle, F.; Hleb, R.; Houška, J.; Šamonil, P.; Hofmeister, J.; Hruška, J. Anthropogenic acidification effects in primeval forests in the Transcarpathian Mts., western Ukraine. Sci. Total Environ. 2010, 408, 856–864. [Google Scholar] [CrossRef]
- Keller, K.; Černý, M.; Bytnerowitz, A.; Smith, L.; Šesták, M.; Michalec, M.; Pernegr, V.; Kučera, J. Air Pollution Status of a Representative Site in the Czech Republic Brdy Mountains. Environ. Pollut. 1997, 98, 291–297. [Google Scholar]
- Percy, K.E.; Ferretti, M. Air pollution and forest health: Toward new monitoring concepts. Environ. Pollut. 2004, 130, 113–126. [Google Scholar] [CrossRef]
- Schröder, W.; Nickel, S.; Jenssen, M.; Riediger, J. Methodology to assess and map the potential development of forest ecosystems exposed to climate change and atmospheric nitrogen deposition: A pilot study in Germany. Sci. Total Environ. 2015, 521–522, 108–122. [Google Scholar] [CrossRef]
- Monserud, R.A.; Sterba, H. A basal area increment model for individual tree growing in even- and uneven-aged forest stands in Austria. For. Ecol. Manag. 1996, 80, 57–80. [Google Scholar] [CrossRef]
- Korf, V. Analysis and pattern of forest stand increment. Lesnictví 1972, 18, 585–604. [Google Scholar]
- Knibbe, B. PAST4—Personal Analysis System for Tree Ring Research Version 4. Instruction Manual; SCIEM/Bernhard Knibbe: Vienna, Austria, 2004; pp. 1–140. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Meloun, M.; Militký, J.; Hill, M.; Brereton, R.G. Crucial problems in regression modelling and their solutions. Analyst 2002, 127, 433–450. [Google Scholar] [CrossRef]
- Chicco, D.; Warrens, M.J.; Jurman, G. The coefficient of determination R-squared is more informative than SMAPE, MAE, MAPE, MSE and RMSE in regression analysis evaluation. Comput. Sci. 2021, 7, e623. [Google Scholar] [CrossRef]
- Urban, J.; Holušová, K.; Menšík, L.; Čermák, J.; Kantor, P. Tree allometry of Douglas fir and Norway spruce on a nutrient-poor and a nutrient-rich site. Trees 2013, 27, 97–110. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Long-term change in the sensitivity of tree-ring growth to climate forcing in Larix decidua. New Phytol. 2006, 170, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Wilczyński, S.; Feliksik, E. The dendrochronological monitoring of the Western Beskid Mountains (Southern Poland) on the basis of radial increments of Norway spruce (Picea abies /L./ Karst.). Electron. J. Pol. Agric. Univ. 2004, 7, 7. [Google Scholar]
- Wang, Z.; Lyu, L.; Liu, W.; Liang, H.; Huang, J.; Zhang, Q.-B. Topographic patterns of forest decline as detected from tree rings and NDVI. Catena 2021, 198, 105011. [Google Scholar] [CrossRef]
- Pechanec, V.; Machar, I.; Kilianová, H.; Vyvlečka, P.; Seják, J.; Pokorný, J.; Štěrbová, L.; Prokopová, M.; Cudlín, P. Ranking the Key Forest Habitats in Ecosystem Function Provision: Case Study from Morava River Basin. Forests 2021, 12, 138. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
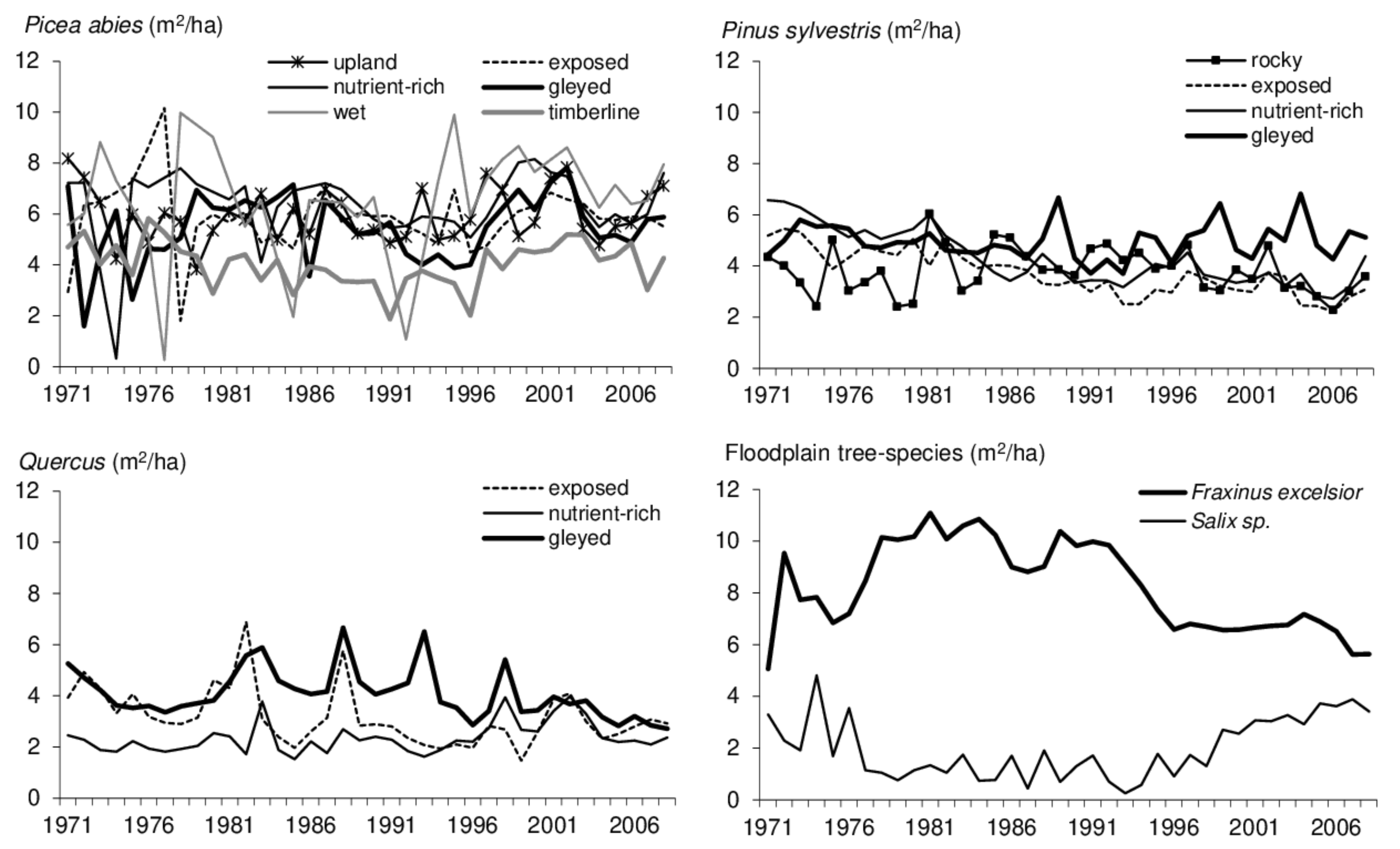{kind=link}
| Relief | Series | A | B | C | D | E | F | G | H | I | J | K | L | M | N | CR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Floodplain | Riverine | 0.2 | 0.1 | 0.1 | 0.7 | 0.1 | 1.1 | 0.0 | 4.2 | 0.4 | 0.1 | 1.0 | 0.5 | 44.1 | 2.0 | 1.5 |
| Swamp | 1.1 | 1.6 | 1.1 | 0.9 | 0.7 | 1.0 | 1.0 | 0.9 | 0.5 | 0.6 | 1.3 | 0.5 | 0.9 | 0.0 | 0.8 | |
| Rocky | Pine | 0.7 | 0.4 | 7.8 | 0.4 | 0.1 | 17.7 | 0.2 | 15.3 | 40.3 | 1.2 | - | 0.4 | 0.4 | - | 4.0 |
| Upland | Exposed | 0.1 | 7.8 | 5.1 | 4.2 | 0.1 | 0.3 | 0.0 | 2.4 | 2.4 | 0.1 | 0.0 | 4.4 | 0.7 | 0.3 | 1.8 |
| Acidic | 1.1 | 3.4 | 14.3 | 10.9 | 0.4 | 3.2 | 0.1 | 21.7 | 5.2 | 0.7 | 0.4 | 11.5 | 30.3 | 0.5 | 5.7 | |
| Nutrient-rich | 0.0 | 16.4 | 4.5 | 4.1 | 0.0 | 1.2 | 0.0 | 34.7 | 2.4 | 1.8 | 0.3 | 6.1 | 21.0 | 9.5 | 4.8 | |
| Gleyed | 0.6 | 0.0 | 2.9 | 0.7 | 0.1 | 10.4 | 0.2 | 17.7 | 0.3 | 0.1 | 0.1 | 0.2 | 0.8 | 0.7 | 1.6 | |
| Karstic | Exposed | 0.2 | 3.3 | 0.1 | 0.7 | 0.1 | 0.0 | 0.2 | - | - | - | 0.5 | 1.2 | - | 0.1 | 0.4 |
| Nutrient-rich | - | - | - | 0.0 | 0.1 | 0.0 | 0.0 | - | - | - | 0.0 | 0.6 | - | - | 0.1 | |
| Wet | 0.0 | - | 0.0 | 0.0 | 0.0 | 6.6 | 0.1 | - | 0.5 | - | - | 0.0 | - | - | 0.2 | |
| Foothill/highland | Ravine | 3.0 | 4.6 | 2.5 | 1.9 | 3.0 | 3.4 | 0.5 | 0.6 | 6.8 | 2.3 | 1.2 | 2.1 | 1.1 | 0.6 | 2.1 |
| Exposed | 1.0 | 10.4 | 4.4 | 5.3 | 1.3 | 0.8 | 1.6 | 0.0 | 2.9 | 3.1 | 6.1 | 8.0 | 0.0 | 4.4 | 3.7 | |
| Acidic | 7.1 | 5.7 | 22.5 | 31.1 | 5.8 | 9.1 | 7.2 | 1.4 | 14.7 | 14.7 | 2.7 | 12.7 | - | 1.3 | 10.7 | |
| Nutrient-rich | 0.2 | 28.0 | 13.4 | 24.2 | 3.9 | 5.4 | 6.5 | 0.5 | 4.1 | 8.6 | 34.7 | 40.5 | 0.7 | 44.2 | 17.9 | |
| Gleyed | 0.3 | 3.5 | 11.2 | 14.2 | 1.4 | 22.7 | 3.4 | 0.1 | 2.4 | 1.3 | 2.3 | 2.2 | 0.1 | 2.0 | 4.4 | |
| Montane | Exposed | 9.9 | 3.1 | 0.9 | 0.0 | 7.0 | - | 3.5 | - | 1.0 | 8.6 | 6.5 | 0.7 | - | 9.7 | 4.4 |
| Acidic | 28.1 | 0.2 | 2.2 | 0.0 | 29.0 | 0.0 | 29.7 | - | 9.5 | 21.9 | 5.6 | 2.8 | - | 0.4 | 11.5 | |
| Nutrient-rich | 10.6 | 9.1 | 0.5 | 0.2 | 12.8 | 0.1 | 22.7 | - | 3.7 | 15.2 | 25.0 | 3.2 | - | 23.5 | 11.1 | |
| Gleyed | 7.6 | 2.0 | 5.4 | 0.1 | 14.5 | 0.1 | 20.2 | - | 1.6 | 3.9 | 4.1 | 2.0 | - | 0.4 | 5.9 | |
| Wet | 0.5 | 0.3 | 0.9 | 0.4 | 2.1 | 17.0 | 1.9 | 0.3 | 1.4 | 1.0 | 0.6 | 0.3 | - | 0.1 | 1.3 | |
| High-mountain | Exposed | 0.7 | - | - | - | 0.9 | - | - | - | - | 2.1 | 1.3 | - | - | 0.1 | 0.5 |
| Acidic | 15.2 | - | - | - | 5.6 | - | - | - | - | 5.3 | 1.7 | - | - | 0.0 | 2.2 | |
| Nutrient-rich | 0.4 | - | - | - | 1.1 | - | - | - | - | 0.1 | 1.6 | - | - | 0.1 | 0.3 | |
| Gleyed | 1.9 | - | - | - | 3.9 | - | 0.1 | - | - | 0.7 | 0.3 | - | - | 0.0 | 0.7 | |
| Wet | 9.1 | - | 0.2 | - | 3.7 | - | 0.9 | - | - | 1.7 | 0.1 | - | - | 0.0 | 1.3 | |
| Timberline | 0.4 | - | - | - | 2.1 | - | - | - | - | 3.5 | 2.2 | - | - | 0.0 | 0.8 | |
| Supalpine | 0.0 | - | - | - | - | - | - | - | - | 1.5 | 0.3 | - | - | - | 0.1 |
| Geotectonics | Relief | Series | Tree-Species | Origin | Altitude | T | P | SO2 | NOx | O3 |
|---|---|---|---|---|---|---|---|---|---|---|
| Bohemian Massif | Rocky | Pine | Pinus sylvestris | Natural | 280–330 | 7.52 ± 0.50 | 687 ± 50 | 6.67 ± 1.43 | 21.03 ± 1.20 | 54.42 ± 2.07 |
| Pinus sylvestris | Planted | 265–296 | 7.59 ± 0.21 | 648 ± 3 | 6.72 ± 1.46 | 21.43 ± 1.22 | 53.65 ± 2.02 | |||
| Upland | Flat | Picea abies | Planted | 255–293 | 8.29 ± 0.13 | 690 ± 10 | 6.75 ± 1.48 | 21.74 ± 1.25 | 53.03 ± 1.99 | |
| Exposed | Quercus petraea | Natural | 516–588 | 7.04 ± 0.16 | 582 ± 14 | 6.19 ± 1.18 | 17.04 ± 1.00 | 62.26 ± 2.74 | ||
| Pinus sylvestris | Planted | 245–267 | 8.33 ± 0.05 | 559 ± 44 | 6.77 ± 1.48 | 21.83 ± 1.25 | 52.86 ± 1.98 | |||
| Picea abies | Planted | 457–503 | 7.17 ± 0.28 | 643 ± 35 | 6.33 ± 1.24 | 18.20 ± 1.04 | 59.97 ± 2.52 | |||
| Nutrient-rich | Quercus petraea | Natural | 386–398 | 7.69 ± 0.04 | 550 ± 4 | 6.50 ± 1.33 | 19.63 ± 1.11 | 57.18 ± 2.27 | ||
| Pinus sylvestris | Planted | 197–262 | 8.56 ± 0.02 | 578 ± 5 | 6.82 ± 1.52 | 22.26 ± 1.29 | 52.02 ± 1.94 | |||
| Picea abies | Planted | 236–253 | 8.47 ± 0.01 | 572 ± 9 | 6.79 ± 1.50 | 22.02 ± 1.27 | 52.49 ± 1.96 | |||
| Gleyded | Quercus robur | Natural | 195–200 | 8.58 ± 0.01 | 559 ± 2 | 6.88 ± 1.56 | 22.78 ± 1.33 | 51.00 ± 1.89 | ||
| Pinus sylvestris | Planted | 192–203 | 8.59 ± 0.06 | 556 ± 7 | 6.88 ± 1.56 | 22.78 ± 1.33 | 51.00 ± 1.89 | |||
| Picea abies | Planted | 277–319 | 8.33 ± 0.29 | 570 ± 6 | 6.68 ± 1.44 | 21.15 ± 1.20 | 54.20 ± 2.06 | |||
| Montane | Foothill | Picea abies | Planted | 500–704 | 6.49 ± 0.61 | 940 ± 65 | 6.15 ± 1.17 | 16.70 ± 1.00 | 62.91 ± 2.82 | |
| Ravine | Pinus sylvestris | Natural | 321–371 | 6.88 ± 0.63 | 624 ± 44 | 6.36 ± 1.27 | 18.46 ± 1.07 | 59.47 ± 2.50 | ||
| Exposed | Picea abies | Planted | 805–840 | 5.05 ± 0.31 | 976 ± 63 | 5.68 ± 1.02 | 12.84 ± 1.01 | 70.49 ± 3.65 | ||
| Nutrient-rich | Picea abies | Planted | 741–820 | 5.17 ± 0.43 | 963 ± 56 | 5.72 ± 1.03 | 13.12 ± 1.00 | 69.93 ± 3.58 | ||
| Gleyed | Picea abies | Planted | 800–840 | 4.99 ± 0.16 | 873 ± 5 | 5.66 ± 1.01 | 12.70 ± 1.01 | 70.76 ± 3.68 | ||
| Wet | Picea abies | Planted | 849–860 | 4.82 ± 0.04 | 871 ± 2 | 5.60 ± 1.00 | 12.14 ± 1.03 | 71.86 ± 3.81 | ||
| Timberline | Picea abies | Natural | 1046–1123 | 3.51 ± 0.34 | 1005 ± 21 | 5.15 ± 1.02 | 8.42 ± 1.26 | 79.16 ± 4.70 | ||
| Carpathians | Floodplain | Swamp | Salix fragilis | Natural | 235–335 | 7.90 ± 0.40 | 677 ± 13 | 6.71 ± 1.45 | 21.36 ± 1.22 | 53.79 ± 2.04 |
| Riverine | Fraxinus excelsior | Natural | 223–313 | 7.96 ± 0.37 | 796 ± 145 | 6.68 ± 1.44 | 21.14 ± 1.21 | 54.22 ± 2.07 | ||
| Montane | Foothill | Picea abies | Planted | 581–626 | 6.61 ± 0.33 | 1107 ± 19 | 6.17 ± 1.17 | 16.88 ± 0.99 | 62.57 ± 2.77 | |
| Ravine | Picea abies | Planted | 1050–1150 | 3.71 ± 0.44 | 1281 ± 88 | 5.12 ± 1.03 | 8.17 ± 1.28 | 79.65 ± 4.76 | ||
| Exposed | Picea abies | Planted | 559–811 | 5.83 ± 0.62 | 1160 ± 77 | 5.89 ± 1.07 | 14.54 ± 0.99 | 67.16 ± 3.27 | ||
| Nutrient-rich | Picea abies | Planted | 697–1062 | 5.31 ± 1.02 | 1191 ± 82 | 5.65 ± 1.04 | 12.61 ± 1.05 | 70.93 ± 3.71 | ||
| Timberline | Picea abies | Natural | 1136–1173 | 3.59 ± 0.19 | 1209 ± 11 | 5.01 ± 1.05 | 7.28 ± 1.34 | 81.39 ± 4.98 |
| Geotectonics | Relief | Series | Tree-Species | Origin | Bedrock | Soil Group | clay | pH | BS | Corg | Al2O3 | CaO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bohemian Massif | Rocky | Pine | Pinus sylvestris | Natural | sandstone | Arenic Podzol | 1.17 ± 0.66 | 4.19 ± 0.25 | 25.46 ± 14.40 | 0.75 ± 0.58 | 0.07 ± 0.03 | 0.02 ± 0.01 |
| Pinus sylvestris | Planted | sandstone | Arenic Podzol | 2.05 ± 1.02 | 4.07 ± 0.26 | 19.14 ± 7.70 | 0.83 ± 0.69 | 0.13 ± 0.07 | 0.02 ± 0.03 | |||
| Upland | Flat | Picea abies | Planted | loess | Haplic Luvisol | 2.73 ± 2.00 | 3.94 ± 0.24 | 12.65 ± 6.86 | 4.08 ± 1.38 | 1.75 ± 0.15 | 0.11 ± 0.02 | |
| Exposed | Quercus petraea | Natural | phonolite | Haplic Cambisol | 17.97 ± 11.28 | 5.27 ± 0.65 | 70.47 ± 32.88 | 5.61 ± 1.81 | 4.10 ± 0.33 | 2.09 ± 0.25 | ||
| Pinus sylvestris | Planted | sandstone | Luvic Cambisol | 15.44 ± 10.54 | 5.85 ± 1.75 | 60.98 ± 42.31 | 1.22 ± 0.32 | 1.27 ± 0.39 | 1.21 ± 1.92 | |||
| Picea abies | Planted | granite | Haplic Cambisol | 12.79 ± 26.22 | 4.22 ± 0.22 | 18.29 ± 5.56 | 2.46 ± 0.63 | 2.99 ± 0.36 | 0.18 ± 0.11 | |||
| Nutrient-rich | Quercus petraea | Natural | phonolite | Haplic Luvisol | 31.47 ± 20.88 | 5.48 ± 0.45 | 92.56 ± 7.09 | 6.46 ± 2.95 | 5.60 ± 1.61 | 0.80 ± 0.22 | ||
| Pinus sylvestris | Planted | phonolite | Epidystric Stagnosol | 2.13 ± 0.35 | 4.34 ± 0.60 | 32.78 ± 19.62 | 2.65 ± 0.83 | 1.03 ± 0.13 | 0.06 ± 0.04 | |||
| Picea abies | Planted | marlstone | Luvic Cambisol | 19.1 ± 15.55 | 3.87 ± 0.11 | 20.63 ± 3.96 | 2.95 ± 2.43 | 1.75 ± 1.33 | 0.04 ± 0.02 | |||
| Gleyded | Quercus robur | Natural | gravel-sand | Endogleyic Stagnosol | 9.85 ± 10.47 | 4.35 ± 0.33 | 22.2 ± 21.23 | 1.75 ± 0.44 | 0.72 ± 0.24 | 0.03 ± 0.03 | ||
| Pinus sylvestris | Planted | gravel-sand | Entic Podzol | 4.86 ± 4.75 | 4.46 ± 0.94 | 34.07 ± 32.61 | 1.21 ± 1.06 | 0.61 ± 0.20 | 0.05 ± 0.05 | |||
| Picea abies | Planted | gravel-sand | Stagni-luvic Cambisol | 19.62 ± 23.46 | 3.85 ± 0.10 | 11.37 ± 3.41 | 4.43 ± 2.35 | 2.02 ± 1.92 | 0.05 ± 0.05 | |||
| Montane | Foothill | Picea abies | Planted | amphibolite | Haplic Cambisol | 2.41 ± 3.22 | 4.40 ± 0.67 | 41.09 ± 32.07 | 9.81 ± 7.17 | 4.16 ± 0.46 | 0.37 ± 0.14 | |
| Ravine | Pinus sylvestris | Natural | sandstone | Arenic Podzol | 6.38 ± 8.09 | 3.72 ± 0.09 | 16.62 ± 6.77 | 1.67 ± 0.70 | 0.33 ± 0.11 | 0.02 ± 0.01 | ||
| Exposed | Picea abies | Planted | orthogneiss | Skeletic Cambisol | 1.10 ± 0.54 | 3.91 ± 0.14 | 16.44 ± 2.96 | 8.96 ± 4.06 | 3.35 ± 0.54 | 0.13 ± 0.06 | ||
| Nutrient-rich | Picea abies | Planted | phyllite | Entic Podzol | 0.98 ± 0.40 | 4.04 ± 0.31 | 17.65 ± 10.1 | 7.15 ± 2.18 | 3.28 ± 1.24 | 0.15 ± 0.04 | ||
| Gleyed | Picea abies | Planted | paragneiss | Stagnic Cambisol | 1.68 ± 0.73 | 3.97 ± 0.32 | 12.05 ± 1.33 | 5.85 ± 1.81 | 3.62 ± 1.19 | 0.16 ± 0.06 | ||
| Wet | Picea abies | Planted | paragneiss | Haplic Gleysol | 2.53 ± 0.63 | 4.02 ± 0.20 | 15.5 ± 6.41 | 6.08 ± 1.94 | 4.15 ± 1.31 | 0.14 ± 0.07 | ||
| Timberline | Picea abies | Natural | orthogneiss | Haplic Podzol | 2.60 ± 0.90 | 3.86 ± 0.18 | 16.56 ± 5.8 | 9.19 ± 8.11 | 1.25 ± 0.49 | 0.07 ± 0.02 | ||
| Carpathians | Floodplain | Swamp | Salix fragilis | Natural | fluvial loam | Stagnic Fluvisol | 16.50 ± 2.15 | 6.10 ± 0.03 | 97.72 ± 2.47 | 4.60 ± 1.46 | 2.29 ± 0.33 | 0.25 ± 0.05 |
| Riverine | Fraxinus excelsior | Natural | fluvial loam | Haplic Fluvisol | 3.18 ± 2.34 | 5.34 ± 1.06 | 66.77 ± 36.38 | 4.07 ± 1.35 | 3.18 ± 1.07 | 0.39 ± 0.32 | ||
| Montane | Foothill | Picea abies | Planted | sandstone | Haplic Cambisol | 1.29 ± 0.32 | 3.78 ± 0.08 | 11.34 ± 4.52 | 10.64 ± 5.61 | 2.68 ± 0.29 | 0.08 ± 0.03 | |
| Ravine | Picea abies | Planted | sandstone | Umbric Cambisol | 15.66 ± 8.11 | 3.95 ± 0.13 | 5.40 ± 1.14 | 4.40 ± 1.01 | 2.55 ± 0.42 | 0.01 ± 0.00 | ||
| Exposed | Picea abies | Planted | sandstone | Skeletic Cambisol | 1.24 ± 0.72 | 3.81 ± 0.17 | 15.63 ± 7.08 | 13.75 ± 3.46 | 2.65 ± 0.38 | 0.10 ± 0.05 | ||
| Nutrient-rich | Picea abies | Planted | sandstone | Skeletic Cambisol | 1.59 ± 0.56 | 3.67 ± 0.07 | 11.63 ± 3.60 | 16.23 ± 5.82 | 2.45 ± 0.40 | 0.06 ± 0.01 | ||
| Timberline | Picea abies | Natural | sandstone | Haplic Podzol | 1.22 ± 0.74 | 3.72 ± 0.10 | 17.85 ± 6.46 | 17.02 ± 14.25 | 1.38 ± 0.46 | 0.07 ± 0.05 |
| Geotectonics | Relief | Series | Tree-Species | Origin | Age | Vha | h | d1.3 | BAI |
|---|---|---|---|---|---|---|---|---|---|
| Bohemian Massif | Rocky | Pine | Pinus sylvestris | Natural | 115 ± 2 | 296 ± 4 | 23 ± 1 | 24.00 ± 2.07 | 4.48 ± 1.62 |
| Pinus sylvestris | Planted | 89 ± 3 | 225 ± 4 | 19 ± 4 | 22.00 ± 2.02 | 3.79 ± 1.53 | |||
| Upland | Flat | Picea abies | Planted | 66 ± 13 | 278 ± 54 | 21 ± 10 | 23.00 ± 1.99 | 7.45 ± 4.4 | |
| Exposed | Quercus petraea | Natural | 77 ± 1 | 151 ± 1 | 19 ± 1 | 22.50 ± 2.74 | 3.17 ± 1.30 | ||
| Pinus sylvestris | Planted | 103 ± 1 | 256 ± 2 | 21 ± 4 | 25.00 ± 1.98 | 3.72 ± 1.37 | |||
| Picea abies | Planted | 109 ± 1 | 342 ± 26 | 23 ± 1 | 24.50 ± 2.52 | 5.28 ± 1.62 | |||
| Nutrient-rich | Quercus petraea | Natural | 100 ± 2 | 203 ± 25 | 19 ± 3 | 27.00 ± 2.27 | 2.36 ± 0.92 | ||
| Pinus sylvestris | Planted | 86 ± 3 | 328 ± 4 | 24 ± 3 | 27.00 ± 1.94 | 4.31 ± 1.28 | |||
| Picea abies | Planted | 85 ± 4 | 446 ± 6 | 25 ± 5 | 27.50 ± 1.96 | 5.94 ± 1.81 | |||
| Gleyded | Quercus robur | Natural | 97 ± 0 | 241 ± 5 | 23 ± 2 | 28.50 ± 1.89 | 4.08 ± 1.20 | ||
| Pinus sylvestris | Planted | 86 ± 1 | 334 ± 11 | 23 ± 0 | 27.00 ± 1.89 | 4.97 ± 1.28 | |||
| Picea abies | Planted | 84 ± 1 | 376 ± 10 | 23 ± 3 | 25.00 ± 2.06 | 5.48 ± 2.21 | |||
| Montane | Foothill | Picea abies | Planted | 75 ± 1 | 365 ± 30 | 24 ± 5 | 29.50 ± 2.82 | 8.18 ± 6.2 | |
| Ravine | Pinus sylvestris | Natural | 123 ± 0 | 256 ± 6 | 22 ± 4 | 28.50 ± 2.50 | 3.39 ± 2.38 | ||
| Exposed | Picea abies | Planted | 96 ± 3 | 505 ± 16 | 28 ± 6 | 36.50 ± 3.65 | 5.89 ± 1.74 | ||
| Nutrient-rich | Picea abies | Planted | 76 ± 8 | 458 ± 37 | 26 ± 14 | 30.00 ± 3.58 | 6.55 ± 2.04 | ||
| Gleyed | Picea abies | Planted | 78 ± 6 | 391 ± 35 | 27 ± 9 | 33.00 ± 3.68 | 8.67 ± 7.56 | ||
| Wet | Picea abies | Planted | 63 ± 8 | 252 ± 22 | 20 ± 11 | 23.50 ± 3.81 | 6.52 ± 2.16 | ||
| Timberline | Picea abies | Natural | 90 ± 6 | 346 ± 15 | 21 ± 8 | 24.50 ± 4.70 | 4.03 ± 1.24 | ||
| Carpathians | Floodplain | Swamp | Salix fragilis | Natural | 45 ± 2 | 220 ± 10 | 14 ± 8 | 36.40 ± 2.04 | 2.02 ± 1.76 |
| Riverine | Fraxinus excelsior | Natural | 74 ± 3 | 402 ± 19 | 26 ± 6 | 31.50 ± 2.07 | 8.47 ± 2.87 | ||
| Montane | Foothill | Picea abies | Planted | 79 ± 5 | 227 ± 12 | 28 ± 2 | 39.06 ± 2.77 | 7.13 ± 5.81 | |
| Ravine | Picea abies | Planted | 72 ± 4 | 216 ± 13 | 18 ± 1 | 26.50 ± 4.76 | 3.45 ± 1.66 | ||
| Exposed | Picea abies | Planted | 89 ± 4 | 555 ± 22 | 30 ± 8 | 33.50 ± 3.27 | 7.14 ± 1.86 | ||
| Nutrient-rich | Picea abies | Planted | 79 ± 4 | 462 ± 15 | 26 ± 1 | 32.50 ± 3.71 | 7.32 ± 2.63 | ||
| Timberline | Picea abies | Natural | 86 ± 6 | 240 ± 20 | 20 ± 8 | 27.50 ± 4.98 | 3.25 ± 1.33 |
| Geotectonics | Relief | Series | Tree-Species | Origin | T | P | SO2 | NOx | O3 | clay | pH | BS | Corg | Al2O3 | CaO | RSD | ER | AR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bohemian Massif | Rocky | Pine | Pinus sylvestris | Natural | −0.09 | 0.02 | 0.39 | 0.08 | −0.03 | 0.29 | 0.16 | −0.08 | −0.29 | 0.18 | −0.08 | 0.24 | −0.14 | −0.72 |
| Pinus sylvestris | Planted | 0.12 | −0.12 | 0.29 | 0.20 | −0.28 | 0.10 | −0.08 | −0.10 | −0.13 | 0.13 | −0.02 | 0.24 | −0.02 | 0.39 | |||
| Upland | Flat | Picea abies | Planted | −0.01 | 0.23 | 0.07 | −0.21 | 0.15 | 0.24 | 0.24 | 0.22 | −0.14 | −0.01 | 0.12 | 1.37 | −1.44 | 0.05 | |
| Exposed | Quercus petraea | Natural | 0.10 | −0.17 | −0.03 | 0.01 | −0.11 | 0.00 | −0.32 | −0.10 | 0.06 | 0.04 | −0.28 | 0.30 | 1.81 | 1.29 | ||
| Pinus sylvestris | Planted | −0.28 | 0.07 | 0.09 | 0.23 | −0.07 | 0.03 | 0.07 | 0.05 | −0.04 | 0.16 | −0.03 | 0.21 | −0.47 | −0.26 | |||
| Picea abies | Planted | −0.26 | 0.17 | 0.13 | −0.41 | 0.19 | −0.05 | 0.02 | −0.11 | −0.06 | −0.01 | −0.08 | 8.51 | −0.50 | −0.28 | |||
| Nutrient−rich | Quercus petraea | Natural | 0.11 | 0.09 | −0.18 | −0.31 | 0.32 | −0.16 | −0.36 | −0.10 | 0.06 | −0.09 | −0.18 | 0.22 | 1.22 | 0.88 | ||
| Pinus sylvestris | Planted | −0.36 | −0.11 | 0.01 | 0.51 | −0.34 | 0.35 | 0.17 | −0.04 | −0.15 | 0.18 | 0.12 | 0.24 | −0.13 | −0.18 | |||
| Picea abies | Planted | −0.05 | −0.05 | −0.12 | 0.27 | −0.15 | 0.27 | 0.17 | −0.01 | −0.08 | 0.00 | 0.11 | 0.43 | −0.31 | 0.16 | |||
| Gleyded | Quercus robur | Natural | −0.09 | −0.29 | 0.38 | 0.34 | −0.29 | 0.04 | −0.12 | −0.27 | 0.06 | −0.13 | −0.24 | 0.21 | −0.76 | 0.22 | ||
| Pinus sylvestris | Planted | 0.00 | −0.01 | −0.23 | 0.19 | −0.07 | 0.39 | 0.23 | −0.17 | −0.09 | 0.30 | 0.02 | 0.19 | −0.26 | 0.32 | |||
| Picea abies | Planted | −0.12 | −0.12 | 0.10 | 0.41 | −0.35 | 0.13 | 0.12 | −0.12 | 0.09 | 0.00 | −0.13 | 0.45 | 0.70 | 0.99 | |||
| Montane | Foothill | Picea abies | Planted | 0.51 | −0.30 | 0.10 | 0.10 | −0.67 | −0.09 | 0.03 | 0.00 | 0.07 | 0.00 | −0.01 | 0.98 | −0.64 | 0.45 | |
| Ravine | Pinus sylvestris | Natural | 0.07 | 0.10 | 0.32 | −0.03 | −0.06 | 0.10 | 0.00 | 0.02 | −0.03 | 0.13 | 0.10 | 0.17 | 0.43 | 0.03 | ||
| Exposed | Picea abies | Planted | −0.03 | −0.01 | −0.30 | 0.04 | 0.00 | −0.06 | 0.05 | 0.10 | 0.13 | −0.14 | 0.03 | 0.28 | 1.36 | 0.18 | ||
| Nutrient−rich | Picea abies | Planted | 0.29 | −0.26 | 0.16 | −0.07 | 0.55 | 0.08 | −0.18 | −0.10 | −0.01 | 0.16 | 0.14 | 0.89 | 4.38 | 1.28 | ||
| Gleyed | Picea abies | Planted | −0.22 | −0.12 | 0.27 | 0.40 | −0.45 | −0.25 | 0.21 | 0.33 | −0.07 | 0.12 | 0.00 | 0.92 | 1.60 | −0.81 | ||
| Wet | Picea abies | Planted | −0.06 | −0.14 | 0.18 | 0.31 | −0.29 | −0.17 | 0.38 | 0.48 | −0.13 | 0.24 | −0.11 | 0.54 | −0.81 | −0.22 | ||
| Timberline | Picea abies | Natural | 0.02 | −0.43 | 0.06 | 0.28 | −0.38 | 0.40 | 0.33 | 0.16 | 0.33 | −0.19 | −0.01 | 0.42 | −0.79 | −0.44 | ||
| Carpathians | Floodplain | Swamp | Salix fragilis | Natural | 0.14 | 0.20 | −0.06 | −0.23 | 0.03 | −0.24 | −0.03 | 0.06 | 0.03 | −0.24 | −0.04 | 1.03 | 3.31 | −0.02 |
| Riverine | Fraxinus excelsior | Natural | −0.32 | −0.15 | 0.24 | 0.02 | 0.19 | 0.31 | 0.16 | −0.19 | −0.23 | 0.25 | −0.02 | 0.71 | 2.18 | 1.64 | ||
| Montane | Foothill | Picea abies | Planted | −0.29 | −0.28 | 0.20 | 0.03 | −0.01 | −0.21 | 0.05 | −0.10 | −0.18 | 0.31 | 0.01 | 0.52 | 4.06 | 2.00 | |
| Ravine | Picea abies | Planted | 0.27 | −0.13 | −0.36 | −0.09 | −0.14 | 0.19 | 0.37 | 0.38 | −0.02 | 0.42 | 0.35 | 0.25 | 1.37 | 1.33 | ||
| Exposed | Picea abies | Planted | 0.42 | −0.40 | −0.01 | −0.46 | −0.39 | 0.07 | 0.30 | 0.14 | −0.06 | 0.22 | 0.25 | 0.68 | 2.81 | −0.03 | ||
| Nutrient−rich | Picea abies | Planted | 0.14 | −0.49 | 0.13 | 0.00 | −0.43 | 0.03 | 0.43 | 0.14 | −0.11 | 0.32 | 0.33 | 0.33 | 0.16 | 0.45 | ||
| Timberline | Picea abies | Natural | 0.03 | −0.08 | −0.17 | 0.08 | 0.05 | 0.10 | 0.26 | 0.32 | −0.01 | 0.35 | 0.36 | 0.38 | 2.35 | 1.46 |
| Geotectonics | Relief | Series | Tree-Species | Origin | Fmin | r2 | T | P | SO2 | NOx | O3 | clay | pH | BS | Corg | Al2O3 | CaO | b |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bohemian Massif | Rocky | Pine | Pinus sylvestris | Natural | 5.07 | 0.61–0.85 | 0.341 | −0.179 | 0.552 | −0.587 | −0.546 | 0.248 | −0.049 | 0.469 | −0.041 | 0.300 | −0.521 | 4.406 |
| Pinus sylvestris | Planted | 1.58 | 0.40–0.45 | 0.140 | 0.174 | 0.544 | −0.081 | −0.059 | −0.021 | −0.025 | −0.080 | −0.050 | 0.155 | 0.272 | 3.017 | |||
| Upland | Flat | Picea abies | Planted | 0.57 | 0.20–0.64 | 0.360 | 1.974 | 5.069 | −2.422 | 1.813 | −0.537 | −0.552 | −0.153 | −1.830 | −1.498 | 0.385 | 10.233 | |
| Exposed | Quercus petraea | Natural | 0.83 | 0.26–0.55 | −0.148 | 0.028 | 0.118 | −0.109 | 0.227 | −0.018 | −0.242 | 0.131 | −0.030 | 0.554 | −0.549 | 3.097 | ||
| Pinus sylvestris | Planted | 1.86 | 0.44–0.69 | 0.060 | −0.050 | −0.395 | 1.468 | 0.104 | −0.164 | −0.097 | 0.248 | −0.150 | 0.479 | −0.642 | 4.400 | |||
| Picea abies | Planted | 0.44 | 0.16–0.43 | −0.058 | −0.209 | −0.105 | −0.249 | −0.055 | −0.141 | −0.672 | 0.862 | −0.064 | −0.045 | −0.160 | 4.625 | |||
| Nutrient−rich | Quercus petraea | Natural | 1.59 | 0.40–0.70 | −0.079 | 0.198 | −0.704 | 0.199 | 0.134 | −0.347 | −0.520 | 0.514 | −0.293 | −0.158 | −0.293 | 2.814 | ||
| Pinus sylvestris | Planted | 4.13 | 0.64–0.71 | −0.133 | −0.022 | −0.526 | 1.116 | 0.090 | 0.006 | −0.077 | −0.040 | 0.047 | 0.383 | −0.457 | 4.865 | |||
| Picea abies | Planted | 2.06 | 0.47–0.58 | 0.590 | −0.001 | 0.507 | −0.710 | −1.229 | −0.717 | −0.278 | 0.718 | −0.149 | 0.595 | −0.723 | 6.887 | |||
| Gleyded | Quercus robur | Natural | 2.91 | 0.55–0.64 | 0.136 | −0.161 | 0.238 | 0.229 | −0.148 | −0.272 | −0.235 | 0.072 | −0.030 | 0.177 | −0.102 | 4.125 | ||
| Pinus sylvestris | Planted | 3.52 | 0.47–0.55 | 0.365 | −0.026 | −0.323 | 0.178 | −0.111 | −0.473 | −0.405 | 0.117 | 0.400 | 0.236 | 0.364 | 5.810 | |||
| Picea abies | Planted | 2.96 | 0.40–0.48 | −0.304 | −0.441 | −0.061 | −0.944 | −0.304 | −0.770 | −1.458 | 0.431 | 0.225 | 1.412 | 0.706 | 4.887 | |||
| Montane | Foothill | Picea abies | Planted | 1.17 | 0.33–0.50 | −0.206 | 0.243 | 0.154 | −0.834 | −0.544 | −1.233 | −1.350 | 0.037 | 0.045 | 0.921 | −0.088 | 9.057 | |
| Ravine | Pinus sylvestris | Natural | 3.04 | 0.43–0.52 | −0.025 | 0.152 | 0.409 | 0.351 | −0.063 | 0.911 | −0.181 | −0.545 | 0.299 | 0.179 | 0.442 | 2.858 | ||
| Exposed | Picea abies | Planted | 4.23 | 0.56–0.72 | 0.382 | −0.223 | −0.090 | −0.131 | 0.457 | −0.447 | −0.556 | 0.025 | −0.682 | −0.717 | 0.610 | 6.117 | ||
| Nutrient-rich | Picea abies | Planted | 0.98 | 0.23–0.42 | −0.301 | −0.444 | −0.292 | −0.081 | −0.653 | −0.603 | −0.628 | 0.508 | −0.275 | −0.441 | 0.791 | 6.777 | ||
| Gleyed | Picea abies | Planted | 3.99 | 0.55–0.85 | 2.567 | −0.051 | −8.886 | 0.066 | −1.305 | 0.472 | −25.071 | 23.376 | −14.938 | 4.943 | 4.655 | 10.945 | ||
| Wet | Picea abies | Planted | 6.45 | 0.70–0.76 | 1.278 | −3.053 | −14.224 | 19.046 | 21.629 | 4.129 | −18.899 | 12.463 | −18.377 | 19.398 | −3.738 | 19.110 | ||
| Timberline | Picea abies | Natural | 11.60 | 0.78–0.84 | 0.019 | −0.141 | −0.583 | −0.631 | 0.795 | −0.381 | −0.392 | 0.405 | −0.597 | −0.275 | 0.046 | 3.875 | ||
| Carpathians | Floodplain | Swamp | Salix fragilis | Natural | 2.69 | 0.53–0.60 | 0.245 | −0.185 | 1.048 | 0.336 | 0.074 | 2.598 | 0.683 | 0.000 | 1.107 | −1.858 | −0.363 | 1.758 |
| Riverine | Fraxinus excelsior | Natural | 1.63 | 0.41–0.77 | −0.504 | 1.055 | −0.860 | 0.713 | −0.024 | −2.397 | 1.598 | −0.888 | −1.602 | 0.808 | −0.518 | 8.951 | ||
| Montane | Foothill | Picea abies | Planted | 4.11 | 0.59–0.66 | 2.635 | −0.003 | 0.226 | 1.251 | 0.025 | 0.562 | −18.671 | 0.033 | 0.585 | 0.073 | 0.640 | 41.357 | |
| Ravine | Picea abies | Planted | 14.89 | 0.82–0.92 | −0.303 | 1.254 | 0.243 | 0.844 | −0.148 | 0.081 | 0.305 | −0.071 | 0.798 | 0.652 | −0.704 | 3.698 | ||
| Exposed | Picea abies | Planted | 8.71 | 0.73–0.84 | −0.015 | 0.116 | −0.862 | −0.292 | 0.122 | −0.290 | 0.132 | 0.021 | −0.230 | 0.303 | −0.402 | 7.097 | ||
| Nutrient−rich | Picea abies | Planted | 5.86 | 0.64–0.70 | −0.644 | 0.793 | 0.976 | −1.067 | 0.326 | 0.591 | −0.361 | −0.793 | 0.245 | 0.850 | −0.027 | 8.381 | ||
| Timberline | Picea abies | Natural | 6.12 | 0.72–0.91 | −0.032 | −0.197 | −0.462 | 0.113 | −0.200 | 0.209 | 0.604 | 0.689 | −0.872 | −0.748 | 0.864 | 4.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samec, P.; Rychtecká, P.; Zeman, M.; Zapletal, M. Environmental Effects among Differently Located and Fertile Sites on Forest Basal-Area Increment in Temperate Zone. Forests 2022, 13, 588. https://doi.org/10.3390/f13040588
Samec P, Rychtecká P, Zeman M, Zapletal M. Environmental Effects among Differently Located and Fertile Sites on Forest Basal-Area Increment in Temperate Zone. Forests. 2022; 13(4):588. https://doi.org/10.3390/f13040588
Chicago/Turabian StyleSamec, Pavel, Petra Rychtecká, Miroslav Zeman, and Miloš Zapletal. 2022. "Environmental Effects among Differently Located and Fertile Sites on Forest Basal-Area Increment in Temperate Zone" Forests 13, no. 4: 588. https://doi.org/10.3390/f13040588
APA StyleSamec, P., Rychtecká, P., Zeman, M., & Zapletal, M. (2022). Environmental Effects among Differently Located and Fertile Sites on Forest Basal-Area Increment in Temperate Zone. Forests, 13(4), 588. https://doi.org/10.3390/f13040588